
Suppressive Effect of Bioactive Extracts of Bacillus sp. H8-1 and Bacillus sp. K203 on Tomato Wilt Caused by Clavibacter michiganensis subsp. michiganensis

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Cell Viability Test and Cellulase Test
2.3. Plant Material and Inoculation
2.4. Bacterial Identification and Characterization
2.5. Preparation of Bacterial Extract and Biocontrol Activity in Tomato Plants
2.6. Cmm Colonization in Tomato Plants
2.7. Antioxidant Enzyme Assay
2.8. Pathogenicity-Related Gene Expression of Cmm
2.9. qRT-PCR for Plant Gene Expression
2.10. Soil Microbial Activities and Diversity by EcoPlate
2.11. Statistical Analyses
3. Results
3.1. Cell Viability and Cellulase Test
3.2. Disease Suppressive Activity
3.3. Bacterial Identification by 16S rRNA Gene Sequencing and Characterization
3.4. Bioactive Extracts Derived from Bacterial Supernatants
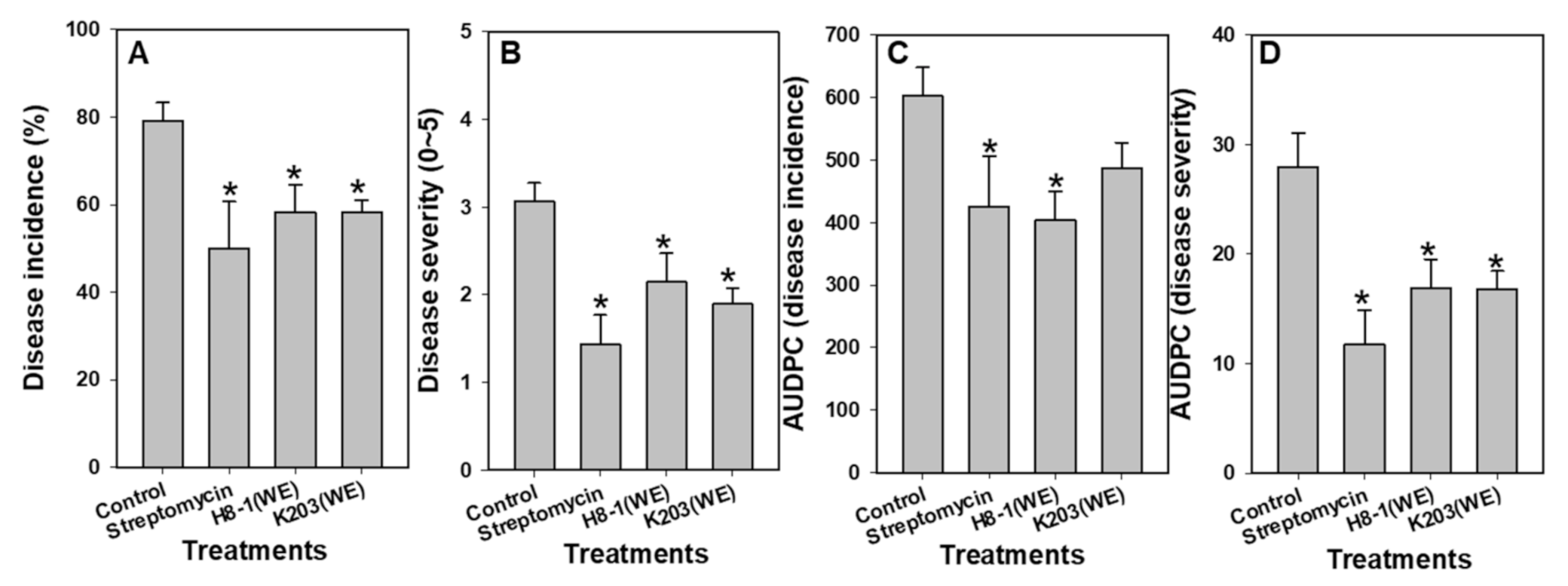
3.5. Biocontrol Activity in Plant Assays
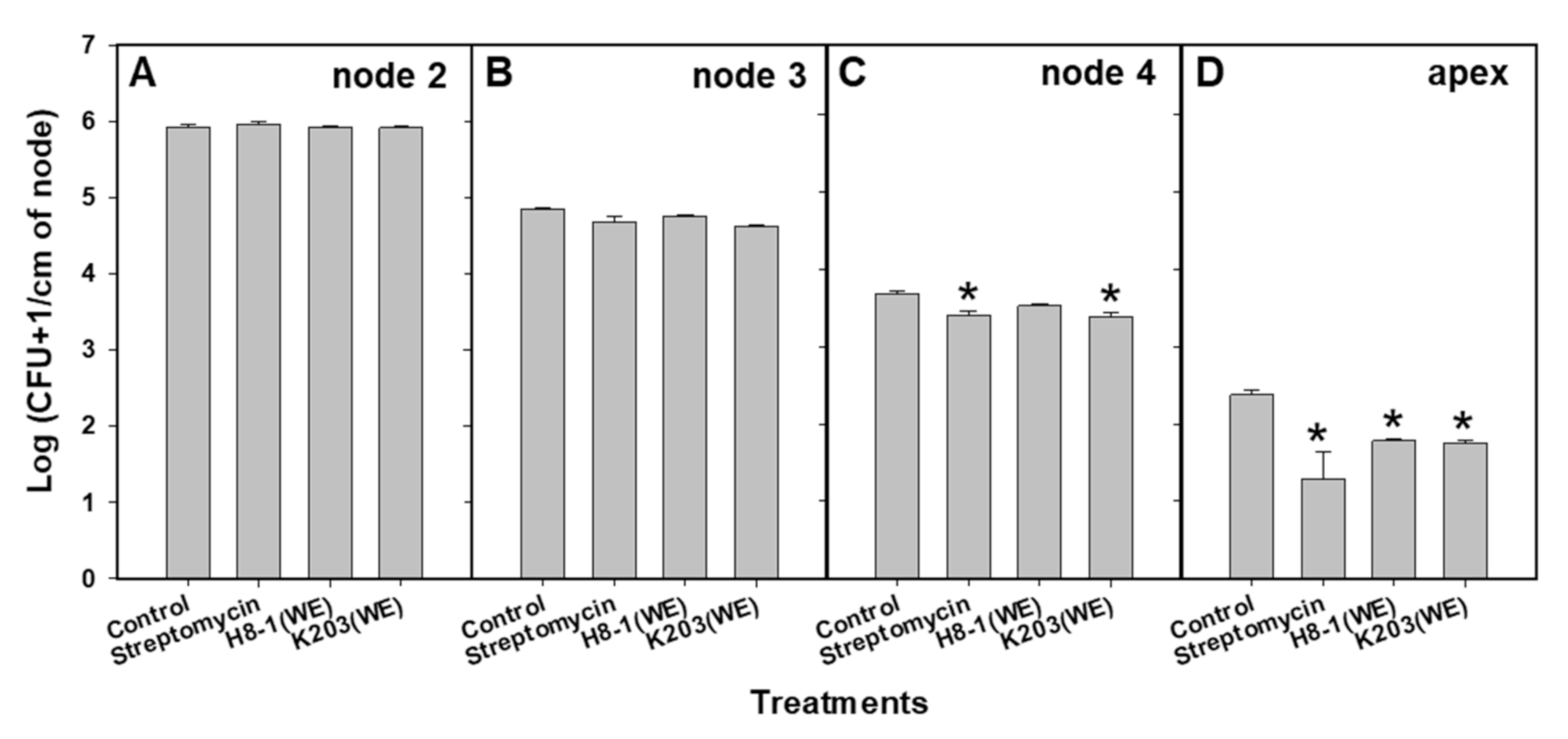
3.6. Colonization of Cmm in Tomato Plants
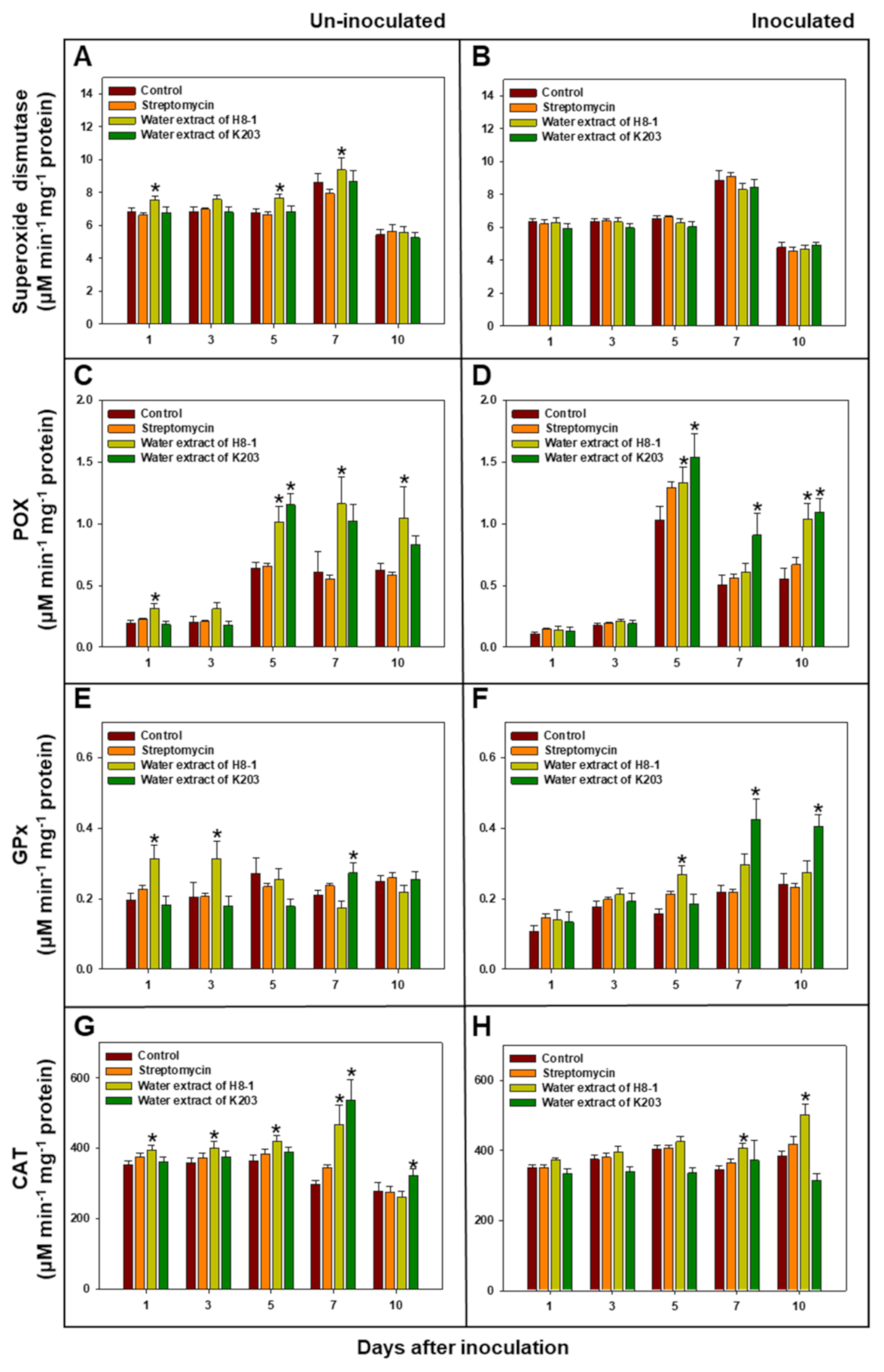
3.7. Antioxidant Enzyme Activity in Tomato Plants
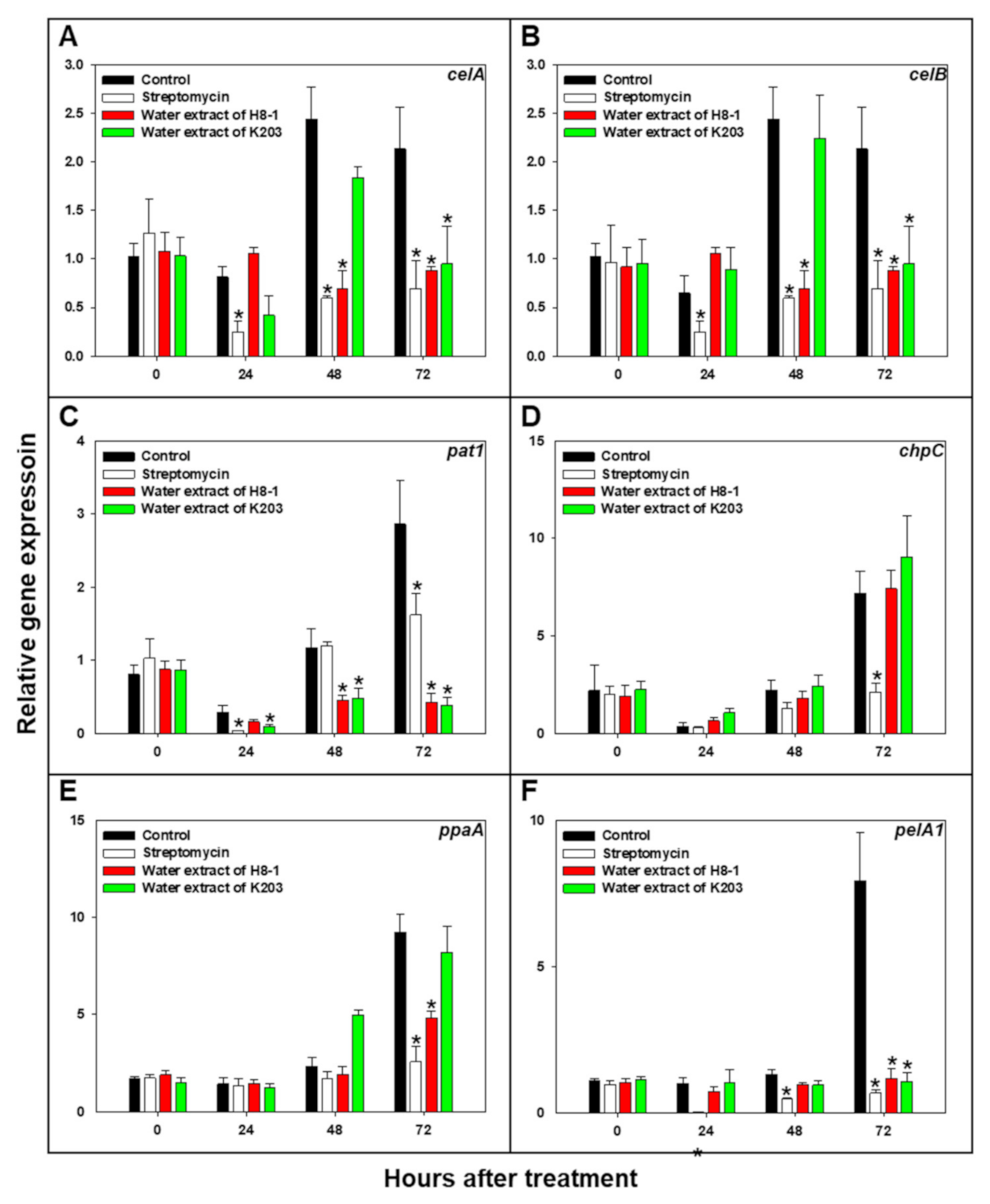
3.8. Cmm Pathogenicity-Related Gene Expression
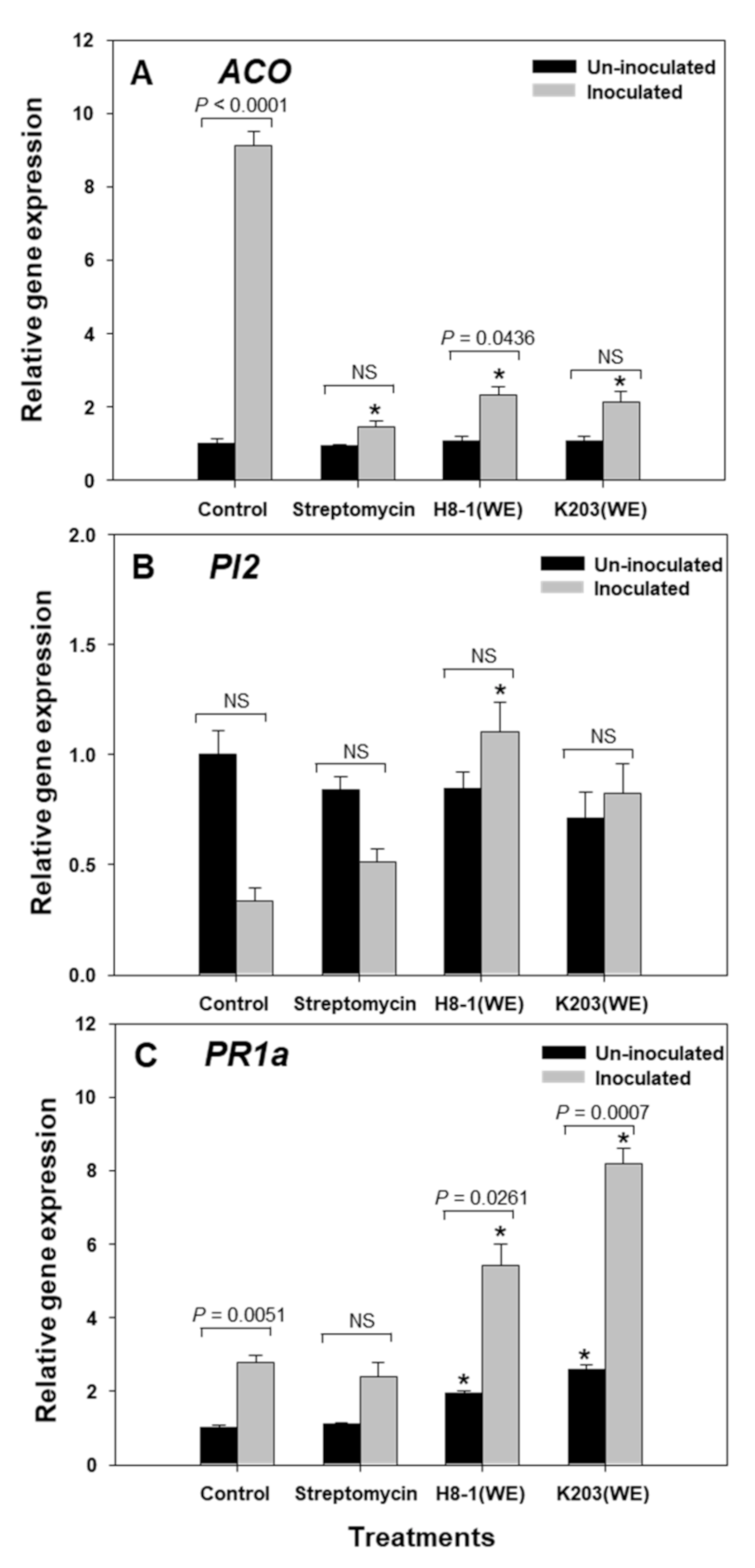
3.9. Relative Expression of Plant Genes, including ACO, PI2, and PR1a
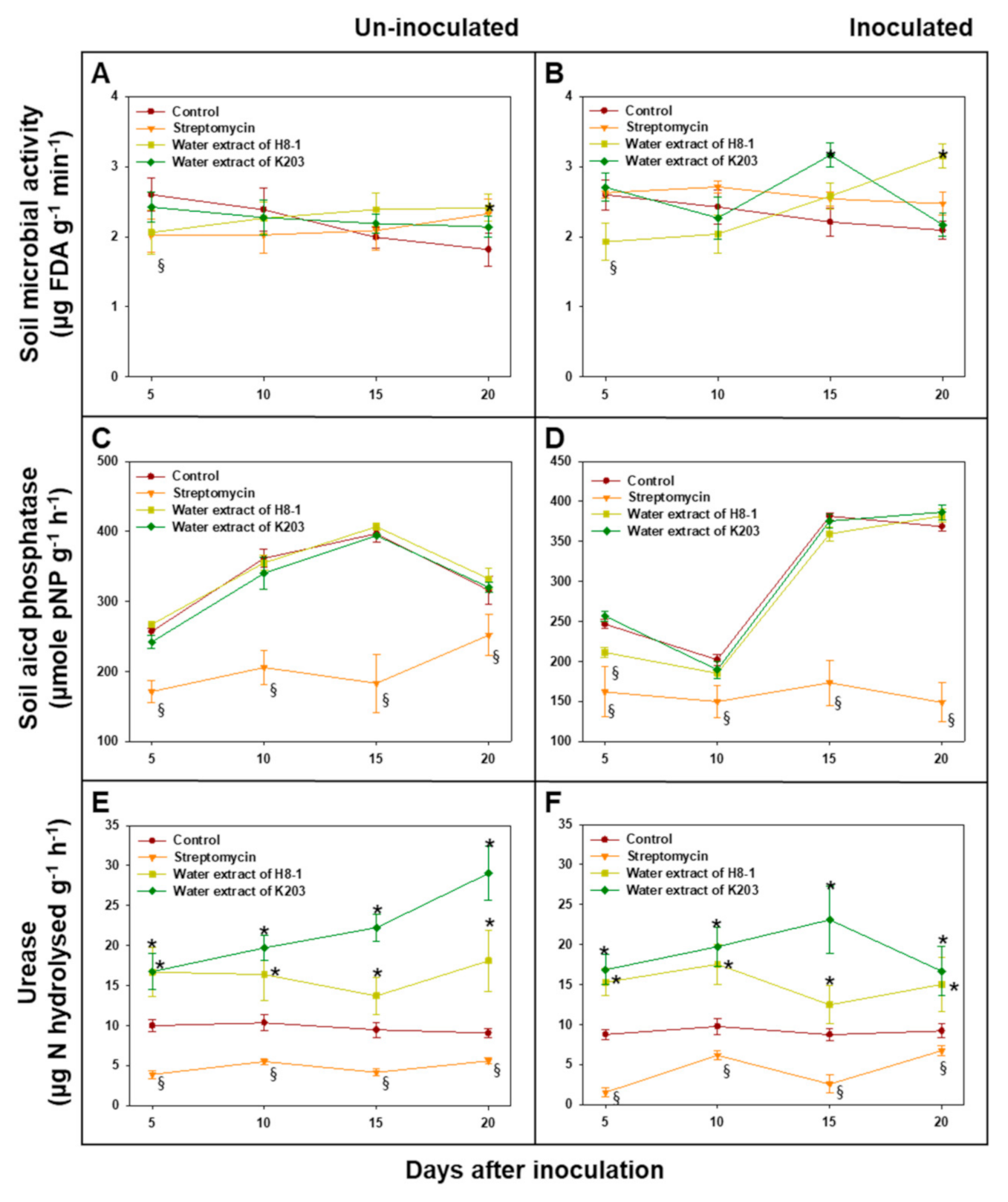
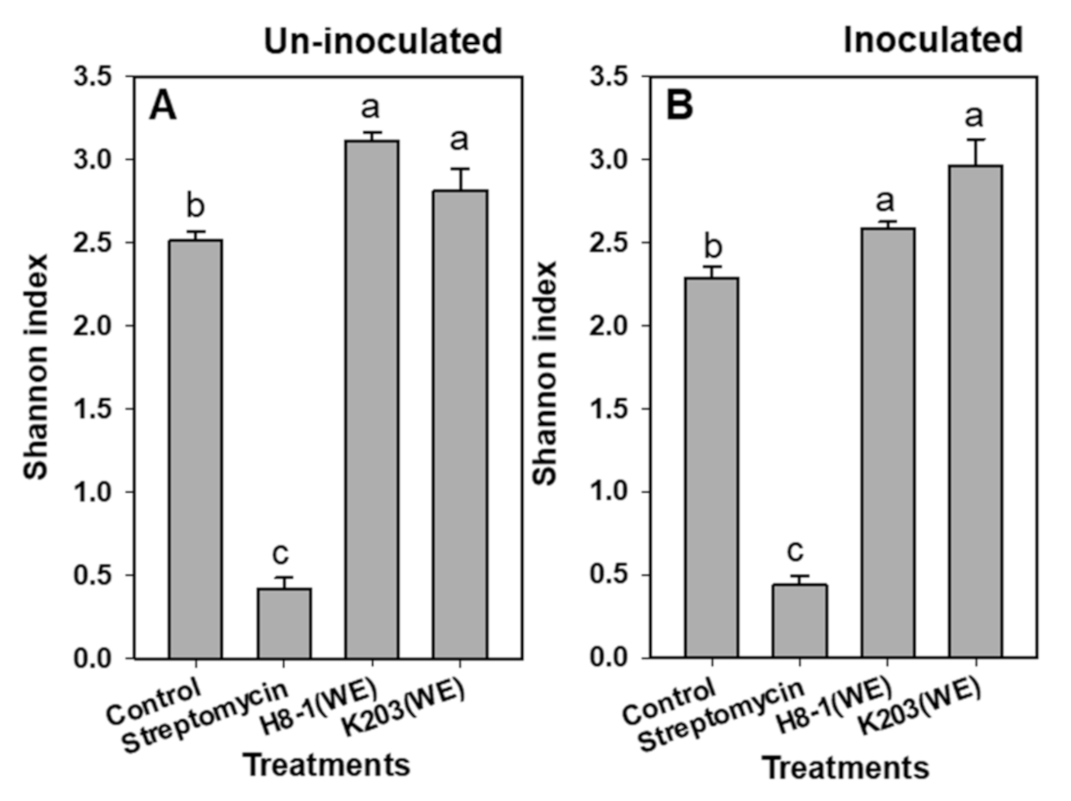
3.10. Soil Microbial Activity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. FAOSTAT. 2020. Available online: http://www.fao.org/faostat/en/#data/QC/visualize (accessed on 27 December 2021).
- Al-Maawali, S.S.; Al-Sadi, A.M.; Ali Khalifa Alsheriqi, S.; Nasser Al-Sabahi, J.; Velazhahan, R. The potential of antagonistic yeasts and bacteria from tomato phyllosphere and fructoplane in the control of Alternaria fruit rot of tomato. All Life 2021, 14, 34–48. [Google Scholar] [CrossRef]
- Eichenlaub, R.; Gartemann, K. The Clavibacter michiganensis subspecies: Molecular investigation of gram-positive bacterial plant pathogens. Annu. Rev. Phytopathol. 2011, 49, 445–464. [Google Scholar] [CrossRef] [PubMed]
- Gartemann, K.; Kirchner, O.; Engemann, J.; Gräfen, I.; Eichenlaub, R.; Burger, A. Clavibacter michiganensis subsp. michiganensis: First steps in the understanding of virulence of a Gram-positive phytopathogenic bacterium. J. Biotechnol. 2003, 106, 179–191. [Google Scholar] [PubMed]
- Sen, Y.; van der Wolf, J.; Visser, R.G.; van Heusden, S. Bacterial canker of tomato: Current knowledge of detection, management, resistance, and interactions. Plant Dis. 2015, 99, 4–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chalupowicz, L.; Barash, I.; Reuven, M.; Dror, O.; Sharabani, G.; Gartemann, K.; Eichenlaub, R.; Sessa, G.; Manulis-Sasson, S. Differential contribution of Clavibacter michiganensis ssp. michiganensis virulence factors to systemic and local infection in tomato. Mol. Plant Pathol. 2017, 18, 336–346. [Google Scholar] [PubMed]
- Strider, D.L. Bacterial canker of tomato caused by Corynebacterium michiganense; a literature review and bibliography. NC Agric. Exp. Stat. Tech. Bull. 1969, 193. [Google Scholar]
- Davis, M.J.; Gillaspie Jr, A.G.; Vidaver, A.K.; Harris, R.W. Clavibacter: A new genus containing some phytopathogenic coryneform bacteria, including Clavibacter xyli subsp. xyli sp. nov., subsp. nov. and Clavibacter xyli subsp. cynodontis subsp. nov., pathogens that cause ratoon stunting disease of sugarcane and bermudagrass stunting disease. Int. J. Syst. Evol. Microbiol. 1984, 34, 107–117. [Google Scholar]
- Stackebrandt, E.; Rainey, F.A.; Ward-Rainey, N.L. Proposal for a new hierarchic classification system, Actinobacteria classis nov. Int. J. Syst. Evol. Microbiol. 1997, 47, 479–491. [Google Scholar] [CrossRef] [Green Version]
- Gartemann, K.; Abt, B.; Bekel, T.; Burger, A.; Engemann, J.; Flügel, M.; Gaigalat, L.; Goesmann, A.; Gräfen, I.; Kalinowski, J. The genome sequence of the tomato-pathogenic actinomycete Clavibacter michiganensis subsp. michiganensis NCPPB382 reveals a large island involved in pathogenicity. J. Bacteriol. 2008, 190, 2138–2149. [Google Scholar] [CrossRef] [Green Version]
- Nandi, M.; Macdonald, J.; Liu, P.; Weselowski, B.; Yuan, Z. Clavibacter michiganensis ssp. michiganensis: Bacterial canker of tomato, molecular interactions and disease management. Mol. Plant Pathol. 2018, 19, 2036–2050. [Google Scholar] [CrossRef] [Green Version]
- Cantarel, B.L.; Coutinho, P.M.; Rancurel, C.; Bernard, T.; Lombard, V.; Henrissat, B. The Carbohydrate-Active EnZymes database (CAZy): An expert resource for glycogenomics. Nucleic Acids Res. 2009, 37, D233–D238. [Google Scholar] [CrossRef] [PubMed]
- Balaji, V.; Mayrose, M.; Sherf, O.; Jacob-Hirsch, J.; Eichenlaub, R.; Iraki, N.; Manulis-Sasson, S.; Rechavi, G.; Barash, I.; Sessa, G. Tomato transcriptional changes in response to Clavibacter michiganensis subsp. michiganensis reveal a role for ethylene in disease development. Plant Physiol. 2008, 146, 1797–1809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nürnberger, T.; Brunner, F.; Kemmerling, B.; Piater, L. Innate immunity in plants and animals: Striking similarities and obvious differences. Immunol. Rev. 2004, 198, 249–266. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, C.; Felix, G. Plants and animals: A different taste for microbes? Curr. Opin. Plant Biol. 2005, 8, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Asai, T.; Tena, G.; Plotnikova, J.; Willmann, M.R.; Chiu, W.; Gomez-Gomez, L.; Boller, T.; Ausubel, F.M.; Sheen, J. MAP kinase signalling cascade in Arabidopsis innate immunity. Nature 2002, 415, 977–983. [Google Scholar] [CrossRef] [PubMed]
- van Loon, L.C.; Rep, M.; Pieterse, C.M. Significance of inducible defense-related proteins in infected plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pieterse, C.M.; Leon-Reyes, A.; Van der Ent, S.; Van Wees, S.C. Networking by small-molecule hormones in plant immunity. Nat. Chem. Biol. 2009, 5, 308–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durrant, W.E.; Dong, X. Systemic acquired resistance. Annu. Rev. Phytopathol. 2004, 42, 185–209. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.M.; Van Wees, S.C.; Hoffland, E.; Van Pelt, J.A.; Van Loon, L.C. Systemic resistance in Arabidopsis induced by biocontrol bacteria is independent of salicylic acid accumulation and pathogenesis-related gene expression. Plant Cell. 1996, 8, 1225–1237. [Google Scholar] [PubMed] [Green Version]
- Pieterse, C.M.; Van Wees, S.C.; Van Pelt, J.A.; Knoester, M.; Laan, R.; Gerrits, H.; Weisbeek, P.J.; Van Loon, L.C. A novel signaling pathway controlling induced systemic resistance in Arabidopsis. Plant Cell. 1998, 10, 1571–1580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pozo, M.J.; Van Der Ent, S.; Van Loon, L.C.; Pieterse, C.M. Transcription factor MYC2 is involved in priming for enhanced defense during rhizobacteria-induced systemic resistance in Arabidopsis thaliana. New Phytol. 2008, 180, 511–523. [Google Scholar] [CrossRef] [PubMed]
- Werner, N.A.; Fulbright, D.W.; Podolsky, R.; Bell, J.; Hausbeck, M.K. Limiting populations and spread of Clavibacter michiganensis subsp. michiganensis on seedling tomatoes in the greenhouse. Plant Dis. 2002, 86, 535–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de León, L.; Siverio, F.; López, M.M.; Rodríquez, A. Comparative efficiency of chemical compounds for in vitro and in vivo activity against Clavibacter michiganensis subsp. Michiganensis, the causal agent of tomato bacterial canker. Crop Prot. 2008, 27, 1277–1283. [Google Scholar] [CrossRef]
- Carvalho, F.P. Agriculture, pesticides, food security and food safety. Environ. Sci. Policy. 2006, 9, 685–692. [Google Scholar] [CrossRef]
- Damalas, C.A.; Eleftherohorinos, I.G. Pesticide exposure, safety issues, and risk assessment indicators. Int. J. Environ. Res. Public Health. 2011, 8, 1402–1419. [Google Scholar] [CrossRef] [PubMed]
- Barratt, B.; Howarth, F.G.; Withers, T.M.; Kean, J.M.; Ridley, G.S. Progress in risk assessment for classical biological control. Biol. Control 2010, 52, 245–254. [Google Scholar] [CrossRef]
- Gay, H. Before and after Silent Spring: From chemical pesticides to biological control and integrated pest management—Britain, 1945–1980. Ambix 2012, 59, 88–108. [Google Scholar] [CrossRef] [PubMed]
- Amkraz, N.; Boudyach, E.H.; Boubaker, H.; Bouizgarne, B.; Aoumar, A.A.B. Screening for fluorescent pseudomonades, isolated from the rhizosphere of tomato, for antagonistic activity toward Clavibacter michiganensis subsp. michiganensis. World J. Microbiol. Biotechnol. 2010, 26, 1059–1065. [Google Scholar] [CrossRef]
- Abo-Elyousr, K.A.; Bagy, H.M.K.; Hashem, M.; Alamri, S.A.; Mostafa, Y.S. Biological control of the tomato wilt caused by Clavibacter michiganensis subsp. michiganensis using formulated plant growth-promoting bacteria. Egypt J. Bio.l Pest Control 2019, 29, 54. [Google Scholar] [CrossRef]
- Shin, D.J.; Yoo, S.-J.; Hong, J.K.; Weon, H.; Song, J.; Sang, M.K. Effect of Bacillus aryabhattai H26-2 and B. siamensis H30-3 on growth promotion and alleviation of heat and drought stresses in Chinese cabbage. Plant Pathol. J. 2019, 35, 178. [Google Scholar] [CrossRef] [PubMed]
- Marefat, A.; Ophel-Keller, K.; McKay, A. A real-time PCR assay for detection of Clavibacter michiganensis subsp. insidiosus in lucerne. Australas. Plant Pathol. 2007, 36, 262–269. [Google Scholar] [CrossRef]
- Baysal, Ö.; Gürsoy, Y.Z.; Örnek, H.; Duru, A. Induction of oxidants in tomato leaves treated with DL-β-Amino butyric acid (BABA) and infected with Clavibacter michiganensis ssp. michiganensis. Eur. J. Plant Pathol. 2005, 112, 361–369. [Google Scholar] [CrossRef]
- Hwang, I.S.; Oh, E.; Lee, H.B.; Oh, C. Functional characterization of two cellulase genes in the Gram-positive pathogenic bacterium Clavibacter michiganensis for wilting in tomato. Mol. Plant-Microbe Interact. 2019, 32, 491–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwak, A.M.; Min, K.J.; Lee, S.Y.; Kang, H.W. Water extract from spent mushroom substrate of Hericium erinaceus suppresses bacterial wilt disease of tomato. Mycobiology 2015, 43, 311–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohd Nadzir, M.M.; Vieira Lelis, F.M.; Thapa, B.; Ali, A.; Visser, R.; van Heusden, A.W.; van der Wolf, J.M. Development of an in vitro protocol to screen Clavibacter michiganensis subsp. michiganensis pathogenicity in different Solanum species. Plant Pathol. 2019, 68, 42–48. [Google Scholar] [CrossRef] [Green Version]
- Ftayeh, R.M.; von Tiedemann, A.; Rudolph, K.W. A new selective medium for isolation of Clavibacter michiganensis subsp. michiganensis from tomato plants and seed. Phytopathology 2011, 101, 1355–1364. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Chalupowicz, L.; Cohen-Kandli, M.; Dror, O.; Eichenlaub, R.; Gartemann, K.; Sessa, G.; Barash, I.; Manulis-Sasson, S. Sequential expression of bacterial virulence and plant defense genes during infection of tomato with Clavibacter michiganensis subsp. michiganensis. Phytopathology 2010, 100, 252–261. [Google Scholar] [CrossRef] [Green Version]
- Takishita, Y.; Charron, J.; Smith, D.L. Biocontrol rhizobacterium Pseudomonas sp. 23S induces systemic resistance in tomato (Solanum lycopersicum L.) against bacterial canker Clavibacter michiganensis subsp. michiganensis. Front Microbiol. 2018, 9, 2119. [Google Scholar] [CrossRef]
- Schnürer, J.; Rosswall, T. Fluorescein diacetate hydrolysis as a measure of total microbial activity in soil and litter. Appl. Environ. Microbiol. 1982, 43, 1256–1261. [Google Scholar] [CrossRef] [Green Version]
- Kandeler, E.; Gerber, H. Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol. Fertility Soils 1988, 6, 68–72. [Google Scholar] [CrossRef]
- Tabatabai, M.A.; Bremner, J.M. Use of p-nitrophenyl phosphate for assay of soil phosphatase activity. Soil Biol. Biochem. 1969, 1, 301–307. [Google Scholar] [CrossRef]
- Xu, W.; Ge, Z.; Poudel, D.R. Application and optimization of biolog ecoplates in functional diversity studies of soil microbial communities. MATEC Web Conf. 2015, 22, 04015. [Google Scholar] [CrossRef] [Green Version]
- Shaner, G.; Finney, R.E. The effect of nitrogen fertilization on the expression of slow-mildewing resistance in Knox wheat. Phytopathology 1977, 67, 1051–1056. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Qian, W.; He, C. The Xanthomonas oryzae pv. oryzae eglXoB endoglucanase gene is required for virulence to rice. FEMS Microbiol. Lett. 2007, 269, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Stork, I.; GARTEMANN, K.; Burger, A.; Eichenlaub, R. A family of serine proteases of Clavibacter michiganensis subsp. michiganensis: chpC plays a role in colonization of the host plant tomato. Mol. Plant Pathol. 2008, 9, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Baysal, Ö.; Soylu, E.M.; Soylu, S. Induction of defence-related enzymes and resistance by the plant activator acibenzolar-S-methyl in tomato seedlings against bacterial canker caused by Clavibacter michiganensis ssp. michiganensis. Plant Pathol. 2003, 52, 747–753. [Google Scholar] [CrossRef]
- Soylu, S.; Baysal, Ö.; Soylu, E.M. Induction of disease resistance by the plant activator, acibenzolar-S-methyl (ASM), against bacterial canker (Clavibacter michiganensis subsp. michiganensis) in tomato seedlings. Plant Sci. 2003, 165, 1069–1075. [Google Scholar] [CrossRef]
- Nicholson, R.L.; Hammerschmidt, R. Phenolic compounds and their role in disease resistance. Annu. Rev. Phytopathol. 1992, 30, 369–389. [Google Scholar] [CrossRef]
- López-Ráez, J.A.; Verhage, A.; Fernández, I.; García, J.M.; Azcón-Aguilar, C.; Flors, V.; Pozo, M.J. Hormonal and transcriptional profiles highlight common and differential host responses to arbuscular mycorrhizal fungi and the regulation of the oxylipin pathway. J. Exp. Bot. 2010, 61, 2589–2601. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Medina, A.; Fernández, I.; Sánchez-Guzmán, M.J.; Jung, S.C.; Pascual, J.A.; Pozo, M.J. Deciphering the hormonal signalling network behind the systemic resistance induced by Trichoderma harzianum in tomato. Front. Plant Sci. 2013, 4, 206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savidor, A.; Teper, D.; Gartemann, K.; Eichenlaub, R.; Chalupowicz, L.; Manulis-Sasson, S.; Barash, I.; Tews, H.; Mayer, K.; Giannone, R.J. The Clavibacter michiganensis subsp. michiganensis–tomato interactome reveals the perception of pathogen by the host and suggests mechanisms of infection. J. Proteome Res. 2012, 11, 736–750. [Google Scholar] [CrossRef] [PubMed]
- Goswami, D.; Patel, K.; Parmar, S.; Vaghela, H.; Muley, N.; Dhandhukia, P.; Thakker, J.N. Elucidating multifaceted urease producing marine Pseudomonas aeruginosa BG as a cogent PGPR and bio-control agent. Plant Growth Regul. 2015, 75, 253–263. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cmm Genes | Sequence (5′ → 3′) | Expressed Gene | References |
|---|---|---|---|
| Gyrase | Forward: GTGGTCGGCGAGGTC Reverse: GCGCGAGCGGGTAG | gyrA | [39] |
| Cellulase A | Forward: GGTTCTCCGCATCAAACTATCC Reverse: TGCTTGTCGCTCGTCGTC | celA | |
| Cellulase B | Forward: GGAGACCACCAGCGACAAG Reverse: TGAACGACCAGAACGACGAG | celB | |
| Serine protease | Forward: GCTGATTCGCGAGAGGATC Reverse: GTTCTCGGTTGCTGTGTCGC | pat-1 | |
| Chp family protease | Forward: GACTGCTAATCACTGTGTTG Reverse: CAATAAACCGTTCCGATGG | chpC | |
| Chymotrypsin-related serine protease | Forward: AATCGGGCTGGTTCTGGTTT Reverse: AGATTCTGCGGCATCTGCAT | ppaA | |
| Pectinase | Forward: GTGCGTTCCTGCGGTAAC Reverse: GCGGATGGTGATGTGGTC | pelA1 |
| Tomato Plant Genes | Sequence (5′ → 3′) | Expressed Gene | Reference |
|---|---|---|---|
| Pathogenesis-related protein | Forward: GTGGGATCGGATTGATATCCT Reverse: CCTAAGCCACGATACCATGAA | PR1a | [40] |
| Proteinase inhibitor | Forward: AATTATCCATCATGGCTGTTCAC Reverse: CCTTTTTGGATCAGATTCTCCTT | PI2 | |
| 1-Aminocyclopropane-1-carboxylix acid oxidase | Forward: AAGATGGCACTAGGATGTCAATAG Reverse: TCCTCTTCTGTCTTCTCAATCAAC | ACO | |
| Glyceraldehyde 3-phosphate dehydrogenase | Forward: CTGGTGCTGACTTCGTTGTTG Reverse: GCTCTGGCTTGTATTCATTCTCG | GAPDH |
| Treatments | Inhibition of Cmm Viability a | Cellulase Activity | Treatments | Inhibition of Cmm Viability | Cellulase Activity | Treatments | Inhibition of Cmm Viability | Cellulase Activity | Treatments | Inhibition of Cmm Viability | Cellulase Activity |
|---|---|---|---|---|---|---|---|---|---|---|---|
| GLSH03 | 98.32 ± 0.22 * | - | TS6-3 | 57.15 ± 1.42 * | + | GLS09 | 22.82 ± 2.33 * | + | JC72 | 5.21 ± 0.21 | + |
| HN12 | 98.27 ± 0.12 * | - | JC35 | 53.70 ± 1.62 * | + | GTH05 | 22.60 ± 2.55 * | - | GLCH05 | 4.85 ± 0.07 | + |
| H8-1 | 98.06 ± 0.03 * | - | TS5-2 | 51.78 ± 2.15 * | + | JTL04 | 22.10 ± 3.22 * | - | JC39 | 1.35 ± 0.30 | - |
| JC34 | 97.91 ± 0.11 * | + | MB7-3 | 48.88 ± 2.40 * | - | JTL08 | 21.54 ± 3.27 * | - | JC18 | −1.66 ± 6.73 | + |
| K203 | 97.71 ± 0.26 * | - | JC46 | 45.07 ± 0.62 * | + | JC54 | 20.82 ± 1.91 * | + | H19-1 | −3.15 ± 1.49 | + |
| H2-7 | 97.51 ± 0.07 * | - | GLSH04 | 41.65 ± 1.99 * | - | GCH09 | 20.61 ± 2.20 * | + | JC55 | −3.82 ± 3.06 | + |
| 5GH 41-08 | 97.39 ± 0.02 * | + | GLSH01 | 41.60 ± 0.68 * | - | HN02 | 18.61 ± 2.86 * | - | JC12 | −3.84 ± 0.02 | - |
| H24-9 | 96.82 ± 0.15 * | + | H18-10 | 41.30 ± 1.81 * | - | HN09 | 18.29 ± 3.71 * | - | HN20 | −4.48 ± 3.90 | + |
| TS3-1 | 95.76 ± 0.26 * | - | GLCH06 | 40.31 ± 0.75 * | - | HN11 | 17.73 ± 3.58 * | - | JC26 | −12.20 ± 4.67 | - |
| HN05 | 95.31 ± 0.06 * | - | JC41 | 37.56 ± 1.21 * | - | MB7-1 | 16.88 ± 5.41 * | - | HN29 | −12.50 ± 6.38 | - |
| GTH01 | 92.78 ± 1.51 * | + | GLSH06 | 36.89 ± 2.63 * | - | GLSH10 | 16.83 ± 1.02 * | + | 5GH31-15 | −14.71 ± 5.85 | + |
| JC33 | 90.61 ± 0.81 * | + | TS7-2 | 34.92 ± 1.19 * | + | JC16 | 16.73 ± 3.39 * | - | H12-10 | −15.01 ± 5.40 | + |
| H8-5 | 76.80 ± 1.92 * | + | H6-4 | 34.65 ± 3.70 * | - | GLC09 | 16.66 ± 2.18 * | - | H1-2 | −17.21 ± 3.61 | + |
| H33-8 | 74.94 ± 1.91 * | + | GCH06 | 34.58 ± 1.57 * | + | GCH05 | 16.24 ± 0.88 * | + | JTL02 | −17.66 ± 0.42 | - |
| TS6-1 | 71.41 ± 0.76 * | + | JC53 | 33.38 ± 3.67 * | - | H6-7 | 15.86 ± 5.30 | - | H1-1 | −23.42 ± 1.84 | - |
| K204 | 69.73 ± 0.19 * | + | GLSH08 | 32.97 ± 4.06 * | - | GLCH04 | 14.14 ± 3.99 | + | JTL05 | −23.48 ± 5.89 | - |
| 5GH 41-07 | 69.16 ± 0.47 * | + | H23-8 | 32.76 ± 3.50 * | + | H20-5 | 13.95 ± 3.62 | + | JTL03 | −26.26 ± 3.37 | - |
| H30-3 | 68.37 ± 1.49 * | + | HN10 | 29.51 ± 0.34 * | - | HN24 | 12.37 ± 5.06 | + | H1-8 | −29.74 ± 2.09 | + |
| MB5-1 | 67.85 ± 0.76 * | + | JC08 | 28.35 ± 3.20 * | - | GLSH09 | 12.29 ± 4.50 | - | JC27 | −31.48 ± 4.00 | + |
| GLC02 | 63.71 ± 1.89 * | + | GLCH09 | 26.76 ± 5.73 * | + | HN22 | 10.03 ± 4.10 | - | JC28 | −40.70 ± 3.22 | + |
| H30-6 | 60.23 ± 1.82 * | + | HN03 | 26.18 ± 3.15 * | + | GLCH03 | 10.01 ± 3.98 | + | JTL06 | −55.43 ± 6.10 | - |
| MB7-5 | 59.51 ± 0.66 * | + | HN08 | 26.18 ± 1.49 * | + | JC59 | 9.82 ± 1.73 | + | JTR09 | −55.44 ± 1.89 | + |
| K185 | 58.63 ± 2.05 * | + | HN25 | 24.06 ± 4.56 * | + | JTR01 | 9.48 ± 4.28 | - | GLSH11 | −72.94 ± 5.66 | - |
| H15-2 | 58.19 ± 2.66 * | + | H5-9 | 23.58 ± 3.64 * | - | GC03 | 6.58 ± 1.68 | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jang, H.; Kim, S.T.; Sang, M.K. Suppressive Effect of Bioactive Extracts of Bacillus sp. H8-1 and Bacillus sp. K203 on Tomato Wilt Caused by Clavibacter michiganensis subsp. michiganensis. Microorganisms 2022, 10, 403. https://doi.org/10.3390/microorganisms10020403
Jang H, Kim ST, Sang MK. Suppressive Effect of Bioactive Extracts of Bacillus sp. H8-1 and Bacillus sp. K203 on Tomato Wilt Caused by Clavibacter michiganensis subsp. michiganensis. Microorganisms. 2022; 10(2):403. https://doi.org/10.3390/microorganisms10020403
Chicago/Turabian StyleJang, Hwajin, Sang Tae Kim, and Mee Kyung Sang. 2022. "Suppressive Effect of Bioactive Extracts of Bacillus sp. H8-1 and Bacillus sp. K203 on Tomato Wilt Caused by Clavibacter michiganensis subsp. michiganensis" Microorganisms 10, no. 2: 403. https://doi.org/10.3390/microorganisms10020403