
Diversity of Microbial Communities of Pinus sylvestris var. mongolica at Spatial Scale

Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Information
2.2. Soil Sampling
2.3. Environmental Variables
2.4. Soil DNA Extraction and Amplification
2.5. Sequence Analysis
2.6. Statistical Analysis
3. Results
3.1. Soil Properties
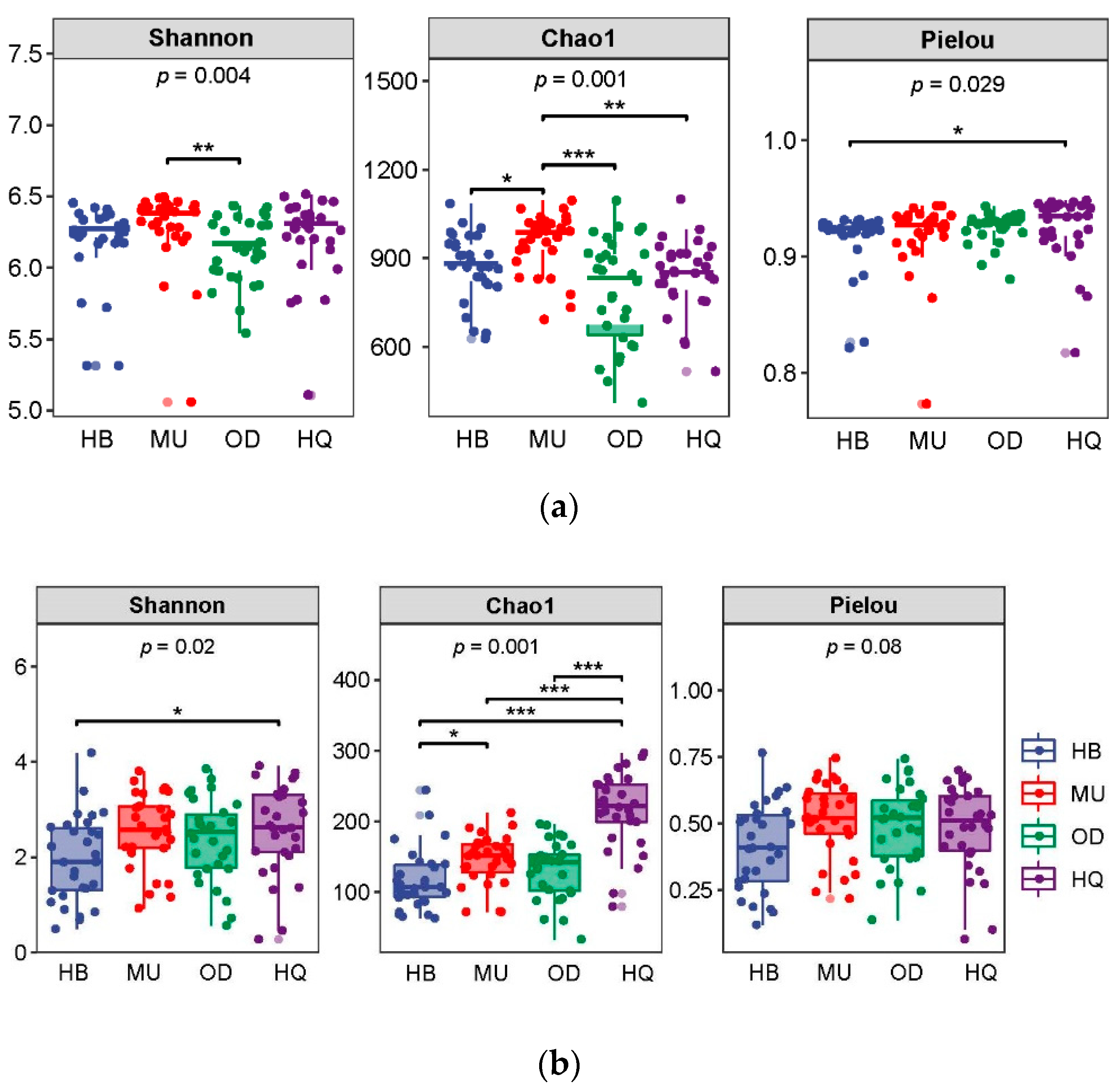
3.2. Diversity of Bacterial and Fungal Communities
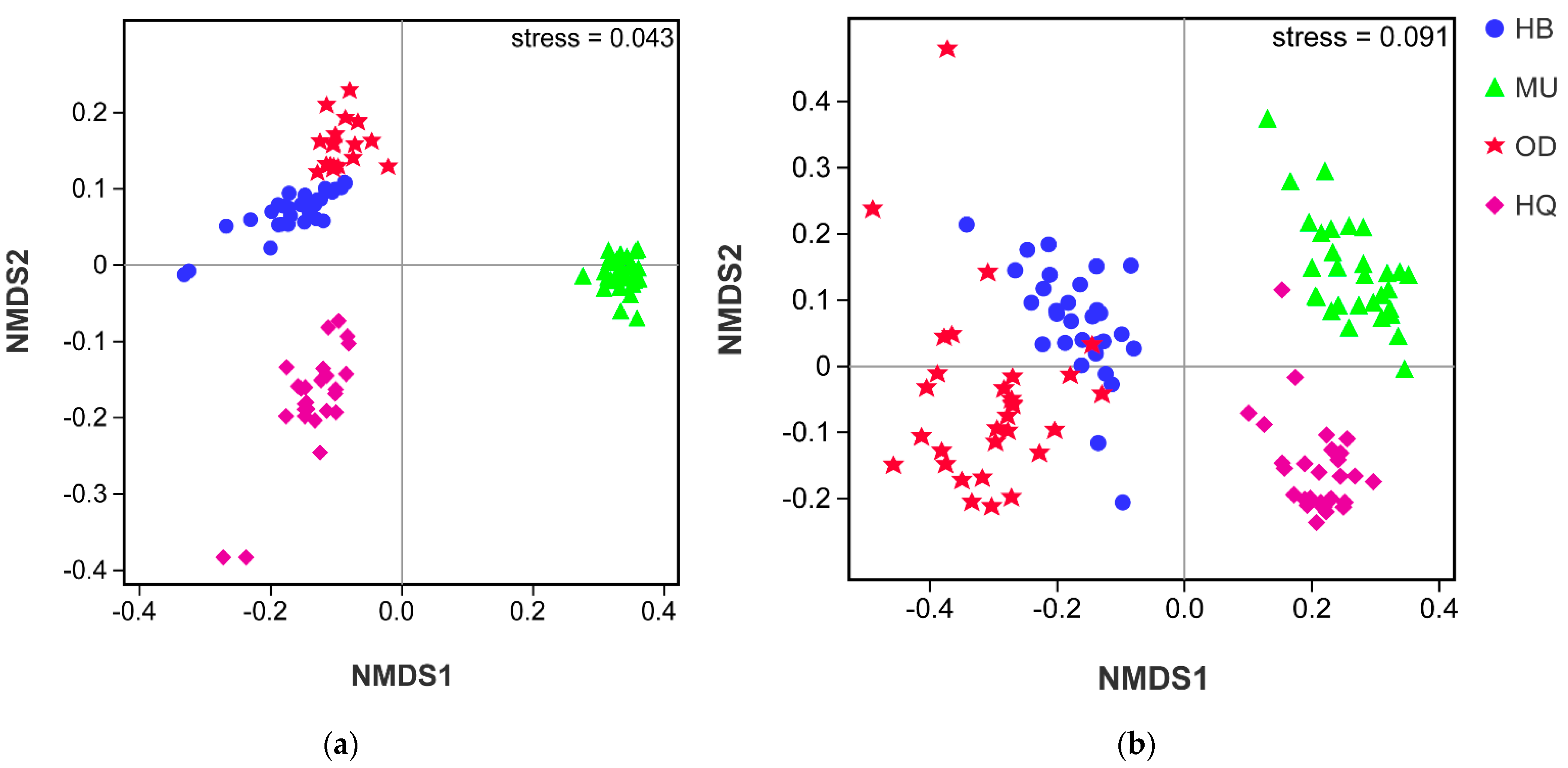
3.3. Bacterial and Fungal Community Composition
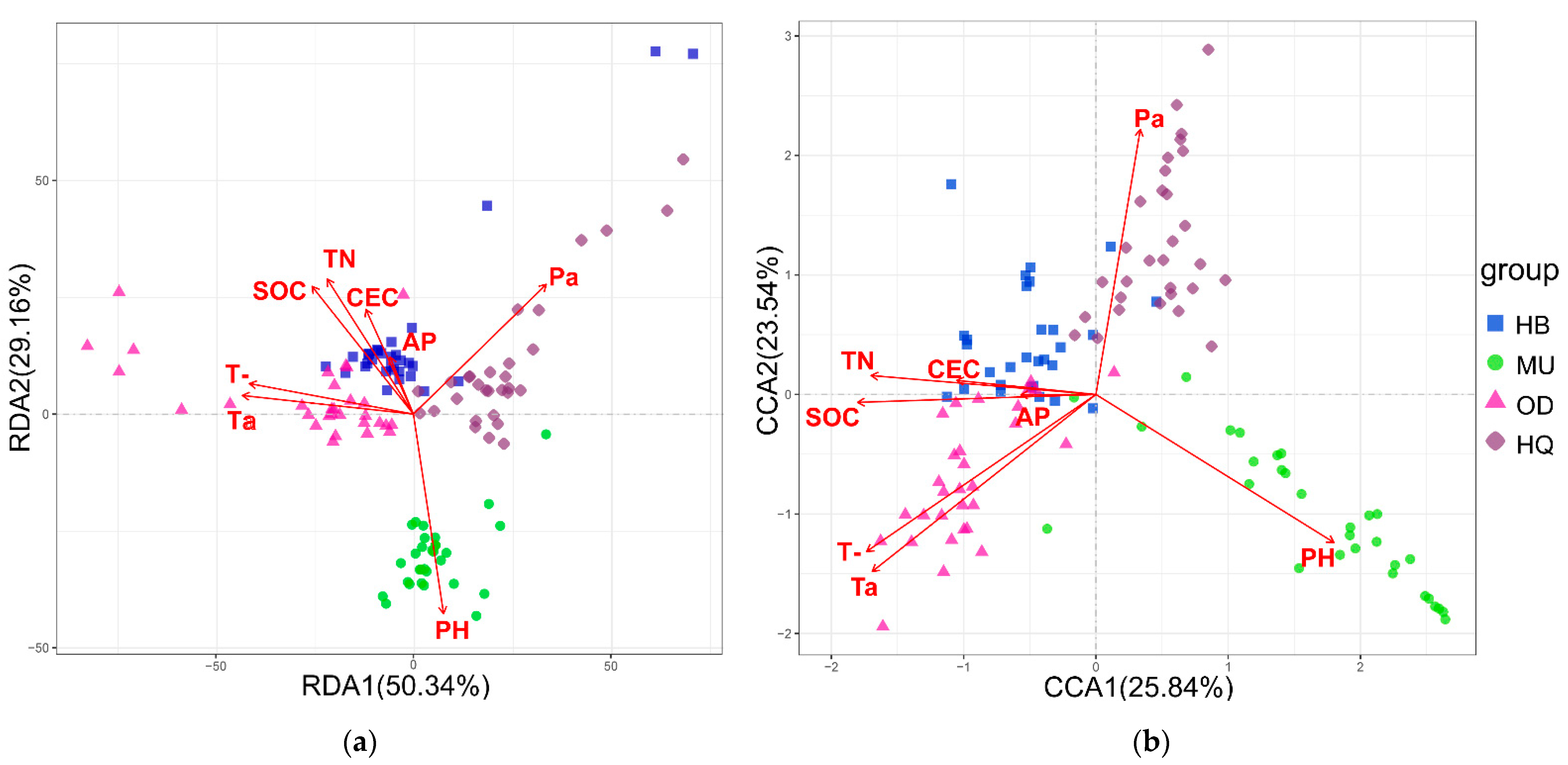
3.4. The Response of Bacterial and Fungal Communities to Environmental Variation
4. Discussion
4.1. Variation in Bacterial and Fungal Composition
4.2. Variation in Bacterial and Fungal Community Structure
4.3. Response of Bacterial and Fungal Communities to Environmental Variation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roesch, L.F.W.; Fulthorpe, R.R.; Riva, A.; Casella, G.; Hadwin, A.K.W.; Kent, A.D.; Daroub, S.H.; Camargo, F.A.O.; Farmerie, W.G.; Triplett, E.W. Pyrosequencing enumerates and contrasts soil microbial diversity. ISME J. 2007, 1, 283–290. [Google Scholar] [CrossRef]
- Zhu, Y.G.; Shen, R.F.; He, J.Z.; Wang, Y.F.; Han, X.G.; Jia, Z.J. China soil microbiome initiative: Progress and perspective. Bull. Chin. Acad. Sci. 2017, 32, 554–565. [Google Scholar]
- Falkowski, P.G.; Fenchel, T.; Delong, E.F. The microbial engines that drive earth’s biogeochemical cycles. Science 2008, 320, 1034–1039. [Google Scholar] [CrossRef] [Green Version]
- Raymond, P.A.; Hartmann, J.; Lauerwald, R.; Sobek, S.; McDonald, C.; Hoover, M.; Butman, D.; Striegl, R.; Mayorga, E.; Humborg, C.; et al. Global carbon dioxide emissions from inland waters. Nature 2013, 503, 355–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, B.; Dai, Z.M.; Wang, H.Z.; Dsouza, M.; Liu, X.M.; He, Y.; Wu, J.J.; Rodrigues, J.L.M.; Gibert, J.A.; Brookes, P.C.; et al. Distinct biogeographic patterns for archaea, bacteria, and fungi along the vegetation gradient at the continental scale in eastern China. mSystems 2017, 2, e00174-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morriën, E.; Hannula, S.E.; Snoek, L.B.; Helmsing, N.R.; Zweers, H.; de Hollander, M.; Soto, R.L.; Bouffaud, M.; Buée, M.; Dimmers, W.; et al. Soil networks become more connected and take up more carbon as nature restoration progresses. Nat. Commun. 2017, 8, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Bardgett, R.D.; Wardle, D.A. Aboveground-Belowground Linkages: Biotic Interactions, Ecosystem Processes, and Global Change; Oxford University Press: Oxford, UK, 2010; 301p. [Google Scholar]
- Friesen, M.L.; Porter, S.S.; Stark, S.C.; von Wettberg, E.J.; Sachs, J.L.; Martinez-Romero, E. Microbially mediated plant functional traits. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 23–46. [Google Scholar] [CrossRef] [Green Version]
- Fierer, N. Embracing the unknown: Disentangling the complexities of the soil microbiome. Nat. Rev. Microbiol. 2017, 15, 579–590. [Google Scholar] [CrossRef]
- Lauber, C.L.; Ramirez, K.S.; Aanderud, Z.; Lennon, J.; Fierer, N. Temporal variability in soil microbial communities across land-use types. ISME J. 2013, 7, 1641–1650. [Google Scholar] [CrossRef]
- Li, J.; Li, Z.A.; Wang, F.M.; Zou, B.; Chen, Y.; Zhao, J.; Mo, Q.F.; Li, Y.W.; Li, X.B.; Xia, H.P. Effects of nitrogen and phosphorus addition on soil microbial community in a secondary tropical forest of China. Biol. Fertil. Soils 2015, 51, 207–215. [Google Scholar] [CrossRef]
- Ding, G.C.; Wan, X.H.; Yang, Q.F.; Lin, Q.X.; Huang, Z.Q. Effects of tree species transition on soil microbial community composition and functions in subtropical China. Chin. J. Appl. Ecol. 2017, 28, 3751–3758. [Google Scholar]
- Wardle, D.A. Ecological linkages between aboveground and belowground biota. Science 2004, 304, 1629–1633. [Google Scholar] [CrossRef] [PubMed]
- Klimek, B.; Chodak, M.; Jaźwa, M.; Niklińska, M. Functional diversity of soil microbial communities in boreal and temperate Scots pine forests. Eur. J. For. Res. 2016, 135, 731–742. [Google Scholar] [CrossRef] [Green Version]
- Shade, A.; Caporaso, J.G.; Handelsman, J.; Knight, R.; Fierer, N. A meta-analysis of changes in bacterial and archaeal communities with time. ISME J. 2013, 7, 1493–1506. [Google Scholar] [CrossRef] [PubMed]
- Green, J.L.; Bohannan, B.J.M.; Whitaker, R.J. Microbial biogeography: From taxonomy to traits. Science 2008, 320, 1039–1043. [Google Scholar] [CrossRef] [Green Version]
- Martiny, J.B.H.; Bohannan, B.J.M.; Brown, J.H.; Colwell, R.K.; Fuhrman, J.A.; Green, J.L.; Horner-Devine, M.C.; Kane, M.; Krumins, J.A.; Kuske, C.R.; et al. Microbial biogeography: Putting microorganisms on the map. Nat. Rev. Microbiol. 2006, 4, 102–112. [Google Scholar] [CrossRef]
- Ramette, A.; Tiedje, J.M. Biogeography: An emerging cornerstone for understanding prokaryotic diversity, ecology, and evolution. Microb. Ecol. 2007, 53, 197–207. [Google Scholar] [CrossRef] [Green Version]
- Yin, H.J.; Zhang, Z.L.; Liu, Q. Root exudates and their ecological consequences in forest ecosystems: Problems and perspective. Acta Phytoecol. Sin. 2018, 42, 1055–1070. [Google Scholar] [CrossRef]
- Waring, G.B. Exploring relationships between enzyme activities and leaf litter decomposition in a wet tropical forest. Soil Biol. Biochem. 2013, 64, 89–95. [Google Scholar] [CrossRef]
- Su, X.L.; Li, Y.B.; Yang, B.; Li, Q. Effects of plant diversity on soil microbial community in a subtropical forest. Chin. J. Ecol. 2018, 37, 2254–2261. [Google Scholar]
- Gourmelon, V.; Maggia, L.; Powell, J.R.; Gigante, S.; Hortal, S.; Gueunier, C.; Letellier, K.; Carriconde, F.; Daffonchio, D. Environmental and geographical factors structure soil microbial diversity in new Caledonian ultramafic substrates: A metagenomic approach. PLoS ONE 2016, 11, e0167405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagova-Mareckova, M.; Cermak, L.; Omelka, M.; Kyselkova, M.; Kopecky, J. Bacterial diversity and abundance of a creek valley sites reflected soil pH and season. Open Life Sci. 2015, 10, 61–70. [Google Scholar] [CrossRef]
- Shen, C.; Xiong, J.; Zhang, H.; Feng, Y.; Lin, X.; Li, X.; Liang, W.; Chu, H. Soil pH drives the spatial distribution of bacterial communities along elevation on Changbai Mountain. Soil Biol. Biochem. 2013, 57, 204–211. [Google Scholar] [CrossRef]
- Geyer, K.M.; Altrichter, A.E.; Takacs-Vesbach, C.D.; Van Horn, D.J.; Gooseff, M.N.; Barrett, J.E. Bacterial community composition of divergent soil habitats in a polar desert. FEMS Microbiol. Ecol. 2014, 89, 490–494. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, R.I.; Thomson, B.C.; James, P.; Bell, T.; Thomson, B.; Bailey, M.; Whiteley, A.S. The bacterial biogeography of British soils. Environ. Microbiol. 2011, 13, 1642–1654. [Google Scholar] [CrossRef]
- Donnarumma, F.; Bazzicalupo, M.; Blažinkov, M.; Mengoni, A.; Sikora, S.; Babić, K.H. Biogeography of Sinorhizobium meliloti nodulating alfalfa in different Croatian regions. Res. Microbiol. 2014, 165, 508–516. [Google Scholar] [CrossRef]
- Pérez-Jaramillo, J.E.; Mendes, R.; Raaijmakers, J.M. Impact of plant domestication on rhizosphere microbiome assembly and functions. Plant Mol. Biol. 2016, 90, 635–644. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Helgason, B.; Wang, L.F.; Winsley, T.; Ferrari, B.C.; Siciliano, S.D. Legacy effects of soil moisture on microbial community structure and N2O emissions. Soil Biol. Biochem. 2016, 95, 40–50. [Google Scholar] [CrossRef]
- Carson, J.K.; Gonzalez-Quiñones, V.; Murphy, D.V.; Hinz, C.; Shaw, J.A.; Gleeson, D.B. Low pore connectivity increases bacterial diversity in soil. Appl. Environ. Microbiol. 2010, 76, 3936–3942. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.T.; Jia, Z.J.; Wang, D.M.; Chiu, C.Y. Effects of temperature on the composition and diversity of bacterial communities in bamboo soils at different elevations. Biogeosciences 2017, 14, 4879–4889. [Google Scholar] [CrossRef] [Green Version]
- Van Horn, D.J.; Van Horn, M.L.; Barrett, J.E.; Gooseff, M.N.; Altrichter, A.E.; Geyer, K.M.; Zeglin, L.H.; Takacs-Vesbaach, C.D.; Gilbert, J.A. Factors controlling soil microbial biomass and bacterial diversity and community composition in a cold desert ecosystem: Role of geographic scale. PLoS ONE 2013, 8, e66103. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.L.; Zhu, J.J.; Kang, H.Z.; Xu, A.H.; Zhang, J.X.; Li, F.Q. Optimum conditions for pure culture of major ectomycorrhizal fungi obtained from Pinus sylvestris var. mongolica plantations in southeastern Keerqin sandy lands, China. J. For. Res. 2008, 19, 113–118. [Google Scholar] [CrossRef]
- Li, M.M.; Ding, G.D.; Gao, G.L.; Zhao, Y.Y.; Wang, D.Y. Introduction suitability of Pinus sylvestris var. mongholica in 10 northern provinces of China. J. Desert Res. 2016, 36, 1021–1028. [Google Scholar]
- Guo, M.S.; Gao, G.L.; Ding, G.D.; Zhang, Y.; Zhao, Y.Y.; Ren, Y. Diversity of ectomycorrhizal fungi associated with Pinus sylvestris var. mongolica in Hulunbuir Sandy Land. Mycosystema 2018, 37, 1133–1142. [Google Scholar]
- QIN, P.Y.; Yang, H.J.; Jiang, F.L.; Zhang, S.B.; Tian, X.M.; Huang, X.R.; Zhang, Z.D. Quantitative classification of natural plant communities in the Saihanba National Nature Reserve, Hebei Province, China. Chin. J. Appl. Ecol. 2016, 27, 1383–1392. [Google Scholar]
- Zhao, P.S.; Guo, M.S.; Gao, G.L.; Zhang, Y.; Ding, G.D.; Ren, Y.; Akhtar, M. Community structure and functional group of root-associated Fungi of Pinus sylvestris var. mongolica across stand ages in the Mu Us Desert. Ecol. Evol. 2020, 10, 3032–3042. [Google Scholar] [CrossRef] [Green Version]
- Han, X.S. Soil Microbial Diversity of Different Sand-Fixation Plantations in Horqin Sandy Land. Master’s Thesis, Northeastern University, Shenyang, China, 2010. [Google Scholar]
- Cao, H.Y.; Gao, G.L.; Ding, G.D.; Zhang, Y.; Zhao, Y.Y.; Ren, Y.; Chen, Y.X.; Guo, M.S. Community Structure and Diversity of Soil Fungi in Four Habitats in Hulun Buir Sandy Land. Sci. Silva Sin. 2019, 55, 118–127. [Google Scholar]
- Bao, S.D. Soil Agrochemical Analysis; China Agriculture Press: Beijing, China, 2000. [Google Scholar]
- Ding, Y.P.; Du, Y.J.; Gao, G.L.; Zhang, Y.; Cao, H.Y.; Zhu, B.B.; Yang, S.Y.; Zhang, J.X.; Qiu, Y.; Liu, H.L. Soil bacterial community structure and functional prediction of Pinus sylvestris var. mongolica plantations in the Hulun Buir Sandy Land. Acta Ecol. Sin. 2021, 41, 4131–4139. [Google Scholar]
- Guo, M.S.; Gao, G.L.; Ding, G.D.; Zhang, Y. Drivers of ectomycorrhizal fungal community structure associated with Pinus sylvestris var. mongolica differ at regional vs. local spatial scales in northern China. Forests 2020, 11, 323. [Google Scholar] [CrossRef] [Green Version]
- Marcel, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet 2011, 17, 10–12. [Google Scholar]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. Dada2: High-resolution sample inference from illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. Mafft: A novel method for rapid multiple sequence alignment based on fast fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree: Computing large minimum evolution trees with profiles instead of a distance matrix. Mol. Biol. Evol. 2009, 26, 1641–1650. [Google Scholar] [CrossRef] [PubMed]
- Urbanová, M.; Šnajdr, J.; Baldrian, P. Composition of fungal and bacterial communities in forest litter and soil is largely determined by dominant trees. Soil Biol. Biochem. 2015, 84, 53–64. [Google Scholar] [CrossRef]
- Dickie, I.A.; Moyersoen, B. Towards a global view of ectomycorrhizal ecology. New Phytol. 2008, 180, 263–265. [Google Scholar] [CrossRef]
- Zhao, P.Y. Assembly mechanism of soil microbial community in Larix principis-rupprechtii Mayr forest of subalpine north China in Shanxi Province. Ph.D. Thesis, Shanxi University, Taiyuan, China, 2019. [Google Scholar]
- Kang, B.T.; Hou, F.J.; Bowatte, S. Characterization of soil bacterial communities in alpine and desert grasslands in the Qilian Mountain range. Pratac. Sci. 2020, 37, 10–19. [Google Scholar]
- Lynd, L.R.; Weimer, P.J.; Van Zyl, W.H.; Pretorius, I.S. Microbial cellulose utilization: Fundamentals and biotechnology. Microbiol. Mol. Biol. Rev. 2002, 66, 506–577. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.F.; Chen, J.H.; Pan, G.X.; Wang, G.M.; Liu, X.Y.; Zhang, X.H.; Li, L.Q.; Bian, R.J.; Cheng, K.; Zheng, J.W. A long-term hybrid poplar plantation on cropland reduces soil organic carbon mineralization and shifts microbial community abundance and composition. Appl. Soil Ecol. 2017, 111, 94–104. [Google Scholar] [CrossRef]
- Stevenson, A.; Hallsworth, J.E. Water and temperature relations of soil Actinobacteria. Env. Microbiol. Rep. 2014, 6, 744–755. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.J.; Gao, G.L.; Chen, L.H.; Ding, G.D.; Zhang, Y.; Cao, H.Y. Soil bacteria community structure and function prediction in the Hulun Buir Sandy Area. China Environ. Sci. 2019, 39, 4840–4848. [Google Scholar]
- Xian, W.D.; Zhang, X.T.; Li, W.J. Research status and prospect on bacterial phylum Chloroflexi. Acta Microbiol. Sin. 2020, 60, 1801–1820. [Google Scholar]
- Xiong, J.B.; Liu, Y.Q.; Lin, X.G.; Zhang, H.Y.; Zeng, J.; Hou, J.Z.; Yang, Y.P.; Yao, T.D.; Knight, R.; Chu, H.Y. Geographic distance and pH drive bacterial distribution in alkaline lake sediments across Tibetan Plateau. Environ. Microbiol. 2012, 14, 2457–2466. [Google Scholar] [CrossRef] [Green Version]
- Pankratov, T.A.; Ivanova, A.O.; Dedysh, S.N.; Liesack, W. Bacterial populations and environmental factors controlling cellulose degradation in an acidic Sphagnum peat. Environ. Microbiol. 2011, 13, 1800–1814. [Google Scholar] [CrossRef]
- Bi, B.Y.; Zhang, H.; Yuan, Y.; Wu, Z.H.; Wang, Y.; Han, F.P. Dynamic changes of soil microbial community in Pinus sylvestris var. mongolica plantations in the Mu Us Sandy Land. J. Environ. Manag. 2021, 287, 112306. [Google Scholar] [CrossRef]
- Wu, Y.T.; Wubet, T.; Trogisch, S.; Both, S.; Scholten, T.; Bruelheide, H.; Buscot, F. Forest age and plant species composition determine the soil fungal community composition in a Chinese subtropical forest. PLoS ONE 2013, 8, e66829. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.B.; Zhu, D.Q.; Zhao, C.C.; Zhang, L.L.; Chen, L.X.; Duan, W.B. Community composition and diversity of fungi in soils under different types of Pinus koraiensis forests. Acta Pedol. Sin. 2019, 56, 1221–1234. [Google Scholar]
- Shen, Z.Z.; Ruan, Y.Z.; Chao, X.; Zhang, J.; Li, R.; Shen, Q.R. Rhizosphere microbial community manipulated by 2 years of consecutive biofertilizer application associated with banana Fusarium wilt disease suppression. Biol. Fertil. Soils 2015, 51, 553–562. [Google Scholar] [CrossRef]
- Paungfoo-Lonhienne, C.; Yeoh, Y.K.; Kasinadhuni, N.R.P.; Lonhienne, T.G.A.; Robinson, N.; Hugenholtz, P.; Ragan, M.A.; Schmidt, S. Nitrogen fertilizer dose alters fungal communities in sugarcane soil and rhizosphere. Sci. Rep. 2015, 5, 8678. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.S.; Wang, L.; Jia, J.W.; Fu, X.H.; Le, Y.Q.; Chen, X.Z.; Sun, Y. Response of soil microbial community in Jiuduansha Wetland to different successional stages and its implications for soil microbial respiration and carbon turnover. Soil Biol. Biochem. 2011, 43, 638–646. [Google Scholar] [CrossRef]
- O’Brien, S.L.; Gibbons, S.M.; Owens, S.M.; Hampton-Marcell, J.; Johnston, E.R.; Jastrow, J.D.; Gilbert, J.A.; Meyer, F.; Antonopoulos, D.A. Spatial scale drives patterns in soil bacterial diversity. Environ. Microbiol. 2016, 18, 2039–2051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lladó, S.; López-Mondéjar, R.; Baldrian, P. Drivers of microbial community structure in forest soils. Appl. Microbiol. Biotechnol. 2018, 102, 4331–4338. [Google Scholar] [CrossRef] [PubMed]
- Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing-based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale. Appl. Environ. Microbiol. 2009, 75, 5111–5120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talbot, J.M.; Bruns, T.D.; Taylor, J.W.; Smith, D.P.; Branco, S.; Glassman, S.I.; Erlandson, S.; Vilgalys, R.; Liao, H.L.; Smith, M.E.; et al. Endemism and functional convergence across the North American soil mycobiome. Proc. Natl. Acad. Sci. USA 2014, 111, 6341–6346. [Google Scholar] [CrossRef] [Green Version]
- Allison, S.; Treseder, K. Warming and drying suppress microbial activity and carbon cycling in boreal forest soils. Glob. Chang. Biol. 2008, 14, 2898–2909. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Deng, Y.; Shen, L.; Wen, C.; Yan, Q.; Ning, D.; Qin, Y.; Xue, K.; Wu, L.; He, Z.; et al. Temperature mediates continental-scale diversity of microbes in forest soils. Nat. Commun. 2016, 7, 12083. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.H.; Gillooly, J.F.; Allen, A.P.; Savage, V.M.; West, G.B. Toward a metabolic theory of ecology. Ecology 2004, 85, 1771–1789. [Google Scholar] [CrossRef]
- Wang, Z.; Brown, J.H.; Tang, Z.; Fang, J. Temperature dependence, spatial scale, and tree species diversity in eastern Asia and North America. Proc. Natl. Acad. Sci. USA 2009, 106, 13388–13392. [Google Scholar] [CrossRef] [Green Version]
- Segura, A.M.; Calliari, D.; Kruk, C.; Fort, H.; Izaguirre, I.; Saad, J.F.; Arim, M. Metabolic dependence of phytoplankton species richness. Glob. Ecol. Biogeogr. 2015, 24, 472–482. [Google Scholar] [CrossRef]
- Lange, M.; Eisenhauer, N.; Sierra, C.A.; Bessler, H.; Engels, C.; Griffiths, R.I.; Mellado-Vázquez, P.G.; Malik, A.A.; Roy, J.; Scheu, S.; et al. Plant diversity increases soil microbial activity and soil carbon storage. Nat. Commun. 2015, 6, 6707. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, M.; Brunner, I.; Hagedorn, F.; Bardgett, R.D.; Stierli, B.; Herzog, C.; Chen, X.; Zingg, A.; Graf-Pannatier, E.B.; Rigling, A.; et al. A decade of irrigation transforms the soil microbiome of a semi-arid pine forest. Mol. Ecol. 2017, 26, 1190–1206. [Google Scholar] [CrossRef] [PubMed]
- Barnard, R.L.; Osborne, C.A.; Firestone, M.K. Responses of soil bacterial and fungal communities to extreme desiccation and rewetting. ISME J. 2013, 7, 2229–2241. [Google Scholar] [CrossRef] [PubMed]
- de Vries, F.T.; Liiri, M.E.; Bjørnlund, L.; Bowker, M.A.; Christensen, S.; Setälä, H.M.; Bardgett, R.D. Land use alters the resistance and resilience of soil food webs to drought. Nat. Clim. Change 2012, 2, 276–280. [Google Scholar] [CrossRef]
- He, J.Z.; Xu, Z.H.; Hughes, J. Molecular bacterial diversity of a forest soil under residue management regimes in subtropical Australia. FEMS Microbiol. Ecol. 2006, 55, 38–47. [Google Scholar] [CrossRef] [Green Version]
- Rousk, J.; Bååth, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef]
- Zhang, K.; Bao, W.K.; Yang, B.; Hu, B. The effects of understory vegetation on soil microbial community composition and structure. Chin. J. Appl. Environ. Biol. 2017, 23, 1178–1184. [Google Scholar]
- Richter, D.D.; Markewitz, D.; Trumbore, S.E.; Wells, C.G. Rapid accumulation and turnover of soil carbon in a re-establishing forest. Nature 1999, 400, 56–58. [Google Scholar] [CrossRef] [Green Version]
- Sul, W.J.; Asuming-Brempong, S.; Wang, Q.; Tourlousse, D.M.; Penton, C.R.; Deng, Y.; Rodrigues, J.L.M.; Adiku, S.G.K.; Jones, J.W.; Zhou, J. Tropical agricultural land management influences on soil microbial communities through its effect on soil organic carbon. Soil Biol. Biochem. 2013, 65, 33–38. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Jiang, Y.; Wang, F.; Wen, C.; Ye, D.; Xue, K.; Qin, Y.; Yang, Y.; Wu, L.; Zhou, J. Long-term soil transplant simulating climate change with latitude significantly alters microbial temporal turnover. ISME J. 2015, 9, 2561–2572. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.J.; Sui, Y.Y.; Yu, Z.H.; Shi, Y.; Chu, H.Y.; Jin, J.; Liu, X.B.; Wang, G.H. High throughput sequencing analysis of biogeographical distribution of bacterial communities in the black soils of northeast China. Soil Biol. Biochem. 2014, 70, 113–122. [Google Scholar] [CrossRef]
- Hollister, E.B.; Engledow, A.S.; Hammett, A.J.M.; Provin, T.L.; Wilkinson, H.H.; Gentry, T.J. Shifts in microbial community structure along an ecological gradient of hypersaline soils and sediments. ISME J. 2010, 4, 829–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | HB | OD | MU | HQ |
|---|---|---|---|---|
| GC | 48°14′–48°18′ N 119°58′–120°1′ E | 42°22′–42°24′ N 117°15′–117°17′ E | 38°18′–38°22′ N 109°40′–109°46′ E | 42°42′–42°43′ N 122°29′–122°30′ E |
| Ele (m) | 790–850 | 1520–1530 | 1100–1130 | 220–240 |
| Ta (°C) | 11.6 | 14.7 | 11.0 | 9.9 |
| T+ (mm) | 18.4 | 21.9 | 18.6 | 16.6 |
| T− (mm) | 4.4 | 7.6 | 3.9 | 3.5 |
| Pa (mm) | 22.7 | 18.0 | 18.7 | 24.8 |
| Soil type | aeolian sandy soil | aeolian sandy soil | aeolian sandy soil | aeolian sandy soil |
| Stand status | normal growth | normal growth | part of the degradation | part of the degradation |
| Site | TN | SOC | AP | pH | CEC |
|---|---|---|---|---|---|
| HB | 0.11 ± 0.01 a | 1.35 ± 0.11 a | 8.61 ± 1.05 a | 5.88 ± 0.02 c | 8.66 ± 0.72 a |
| HQ | 0.04 ± 0.00 b | 0.42 ± 0.04 b | 2.78 ± 0.25 c | 6.29 ± 0.11 b | 2.65 ± 0.13 c |
| MU | 0.01 ± 0.01 c | 0.18 ± 0.01 c | 4.32 ± 0.18 bc | 8.31 ± 0.04 a | 2.88 ± 0.06 c |
| OD | 0.10 ± 0.01 a | 1.39 ± 0.08 a | 5.39± 0.24 b | 6.15 ± 0.02 b | 5.35 ± 0.33 b |
| TN | SOC | AP | pH | CEC | Pa | T− | Ta | T+ | |
|---|---|---|---|---|---|---|---|---|---|
| Shannon | −0.261 ** | −0.287 ** | −0.385 ** | 0.344 ** | −0.19 * | 0.105 | −0.246 ** | −0.246 ** | −0.105 |
| Chao1 | −0.306 ** | −0.312 ** | −0.226 * | 0.353 ** | −0.159 | −0.026 | −0.134 | −0.134 | 0.026 |
| Pielou | 0.016 | −0.025 | −0.313 ** | 0.181 * | −0.05 | 0.055 | −0.133 | −0.133 | −0.055 |
| TN | SOC | AP | pH | CEC | Pa | T− | Ta | T+ | |
|---|---|---|---|---|---|---|---|---|---|
| Shannon | −0.160 | −0.166 | −0.257 ** | 0.201 * | −0.215 * | 0.015 | −0.162 | −0.162 | −0.015 |
| Chao1 | −0.132 | −0.131 | −0.177 | 0.196 * | −0.152 | −0.075 | −0.066 | −0.066 | 0.075 |
| Pielou | −0.211 * | −0.238 ** | −0.484 ** | 0.126 | −0.363 ** | 0.372 ** | −0.519 ** | −0.519 ** | −0.372 ** |
| TN | SOC | AP | pH | CEC | Pa | T− | Ta | T+ | |
|---|---|---|---|---|---|---|---|---|---|
| r | 0.15346 | 0.16540 | 0.00052 | 0.07494 | 0.03636 | 0.32330 | 0.36799 | 0.44175 | 0.47109 |
| p | 0.002 | 0.001 | 0.465 | 0.042 | 0.209 | 0.001 | 0.001 | 0.001 | 0.001 |
| TN | SOC | AP | pH | CEC | Pa | T− | Ta | T+ | |
|---|---|---|---|---|---|---|---|---|---|
| r | 0.0344 | 0.02711 | 0.01592 | 0.08055 | 0.04986 | 0.14459 | 0.14554 | 0.17623 | 0.19962 |
| p | 0.157 | 0.205 | 0.364 | 0.018 | 0.138 | 0.001 | 0.002 | 0.001 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, D.-D.; Zhao, W.; Reyila, M.; Huang, K.-C.; Liu, S.; Cui, B.-K. Diversity of Microbial Communities of Pinus sylvestris var. mongolica at Spatial Scale. Microorganisms 2022, 10, 371. https://doi.org/10.3390/microorganisms10020371
Wang D-D, Zhao W, Reyila M, Huang K-C, Liu S, Cui B-K. Diversity of Microbial Communities of Pinus sylvestris var. mongolica at Spatial Scale. Microorganisms. 2022; 10(2):371. https://doi.org/10.3390/microorganisms10020371
Chicago/Turabian StyleWang, Dan-Dan, Wen Zhao, Mumin Reyila, Kai-Chuan Huang, Shun Liu, and Bao-Kai Cui. 2022. "Diversity of Microbial Communities of Pinus sylvestris var. mongolica at Spatial Scale" Microorganisms 10, no. 2: 371. https://doi.org/10.3390/microorganisms10020371