
Novel Methanobacterium Strain Induces Severe Corrosion by Retrieving Electrons from Fe0 under a Freshwater Environment

 , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
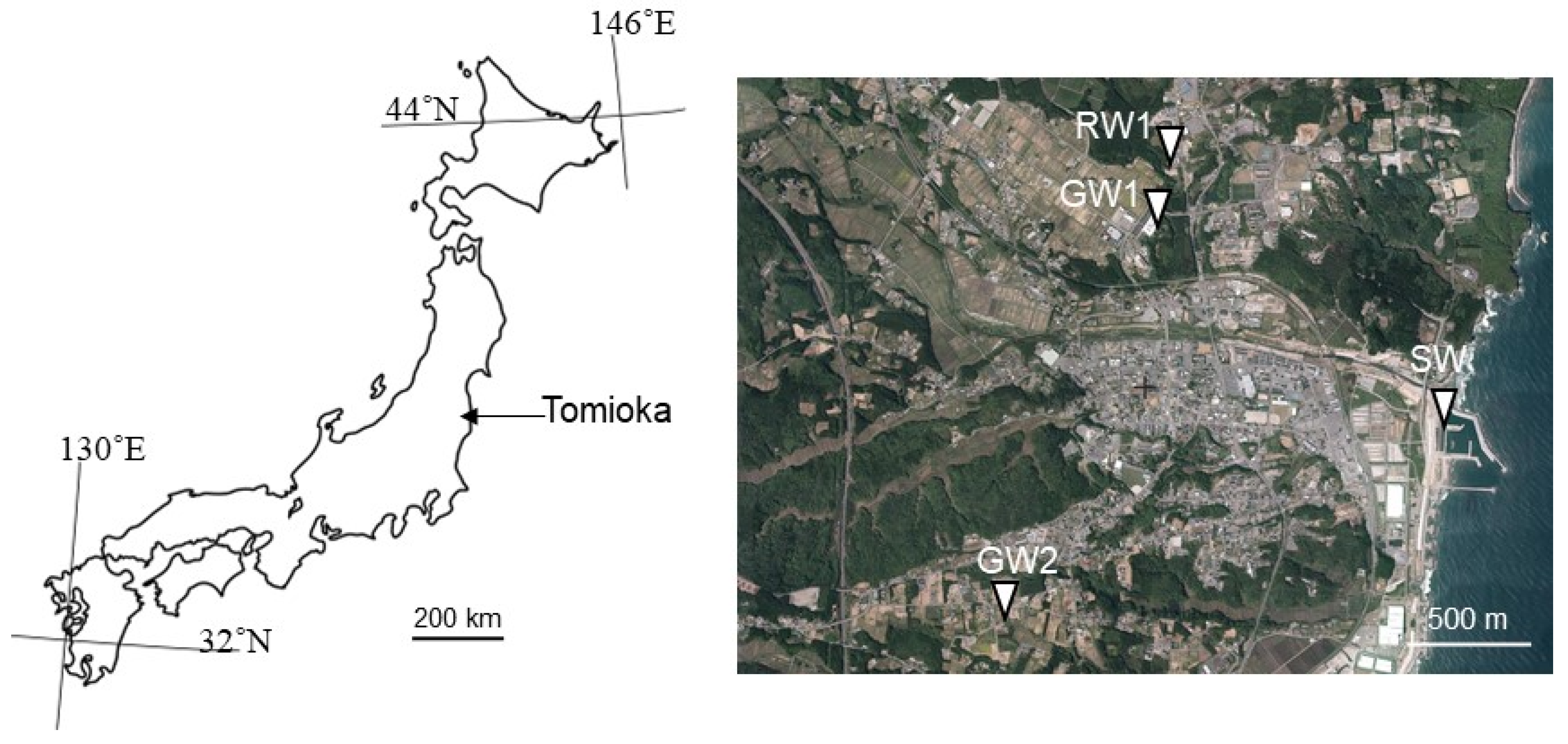
2.1. Site and Sample Description
2.2. Analytical Methods
2.3. Cultivation Methods for Enrichment and Isolation of Microorganisms with EMIC Activity
2.3.1. Enrichment Method of Iron-Corroding Microorganisms from Collected Samples
2.3.2. Isolation of Methanogens with EMIC Activity
2.4. Electrical Incubation and Cyclic Voltammetry Measurement of Cells
3. Results
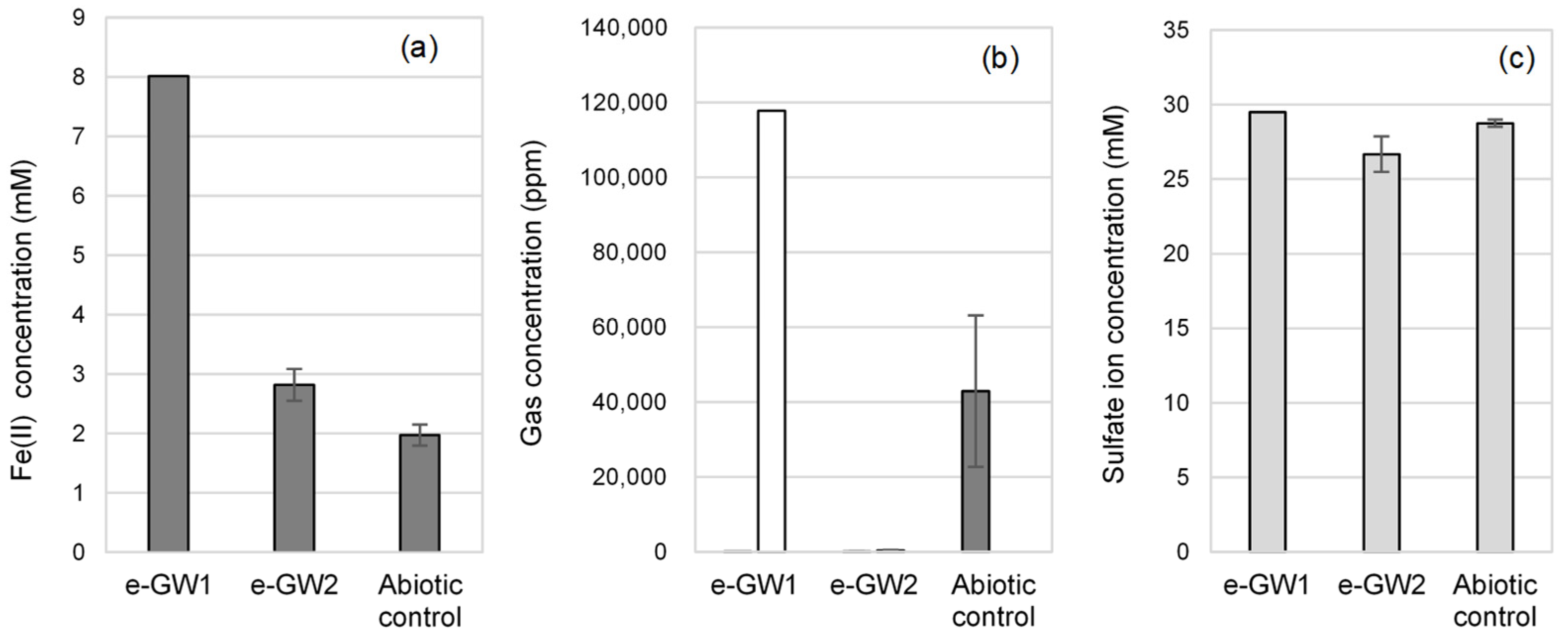
3.1. Water Chemistry
3.2. Enrichment and Isolation of Microorganisms with EMIC Activity
3.2.1. Enrichment of Iron-Corroding Microbial Communities
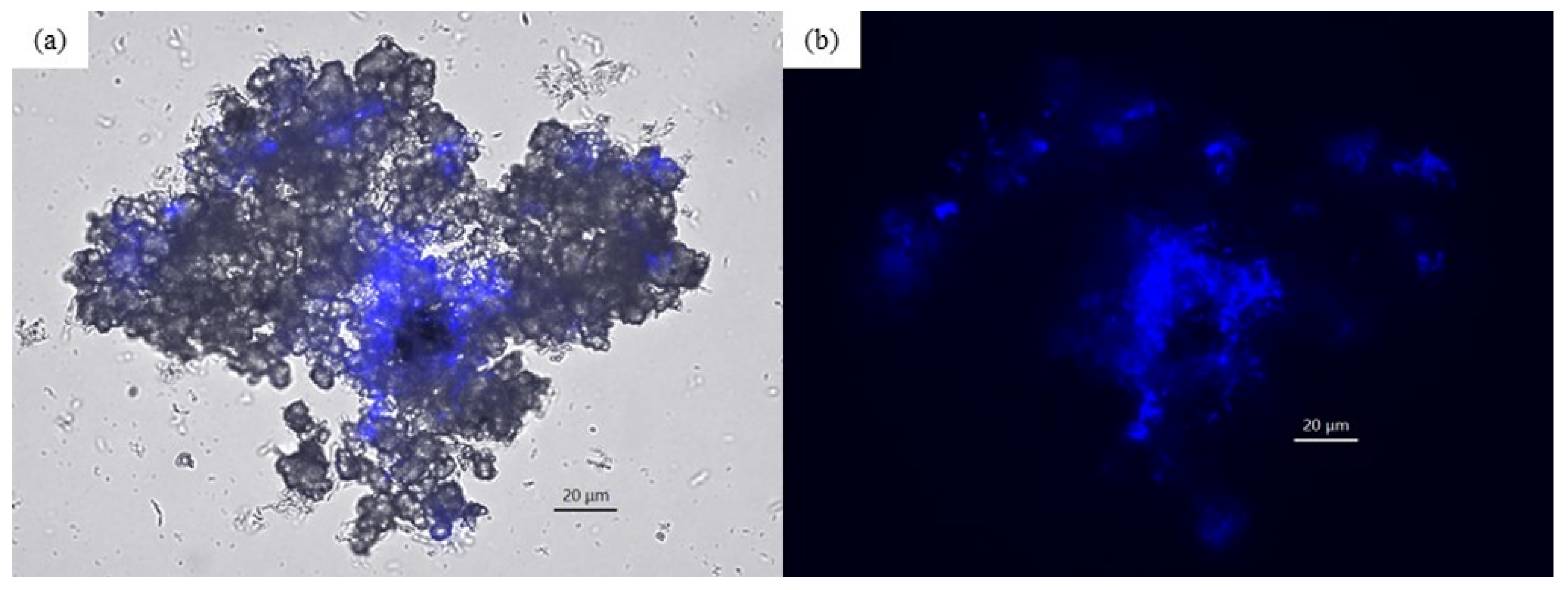
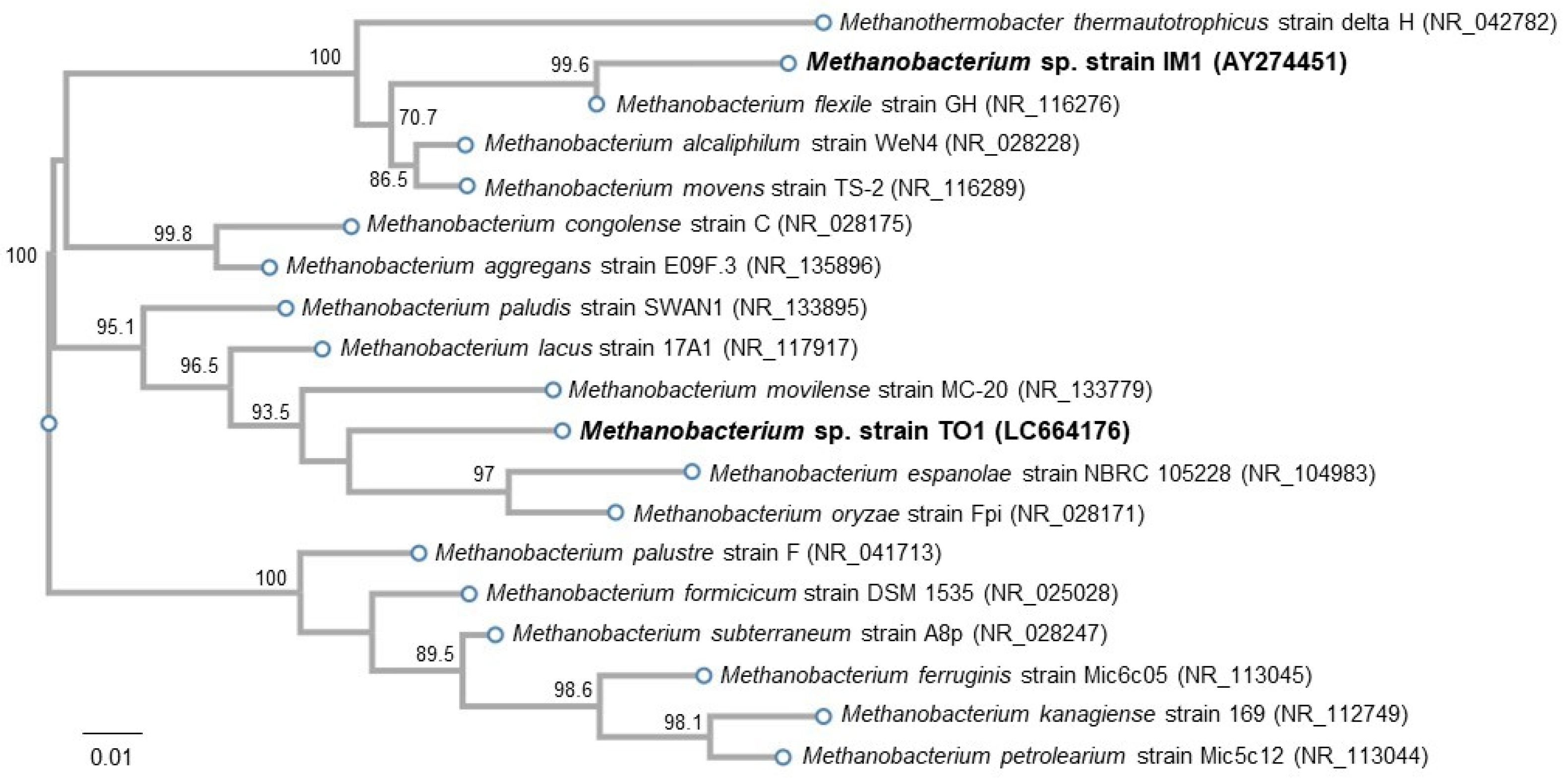
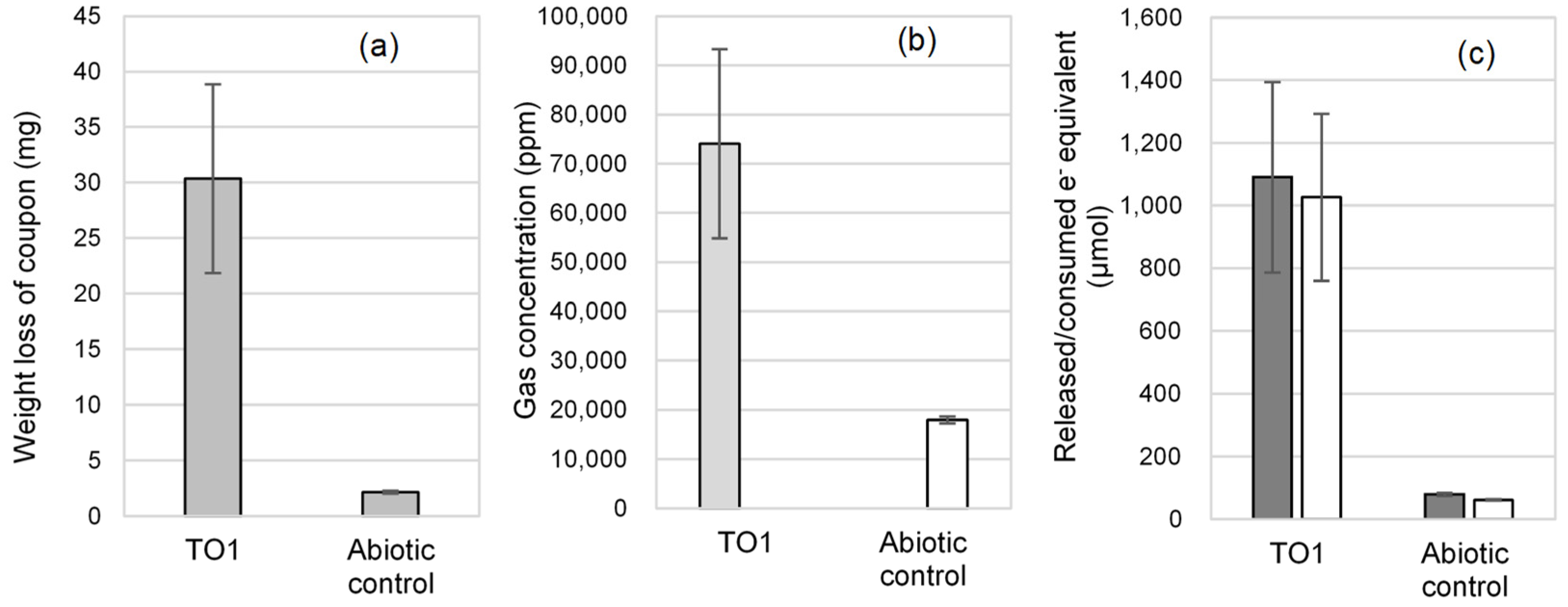
3.2.2. Isolation of Microorganisms with EMIC Activity
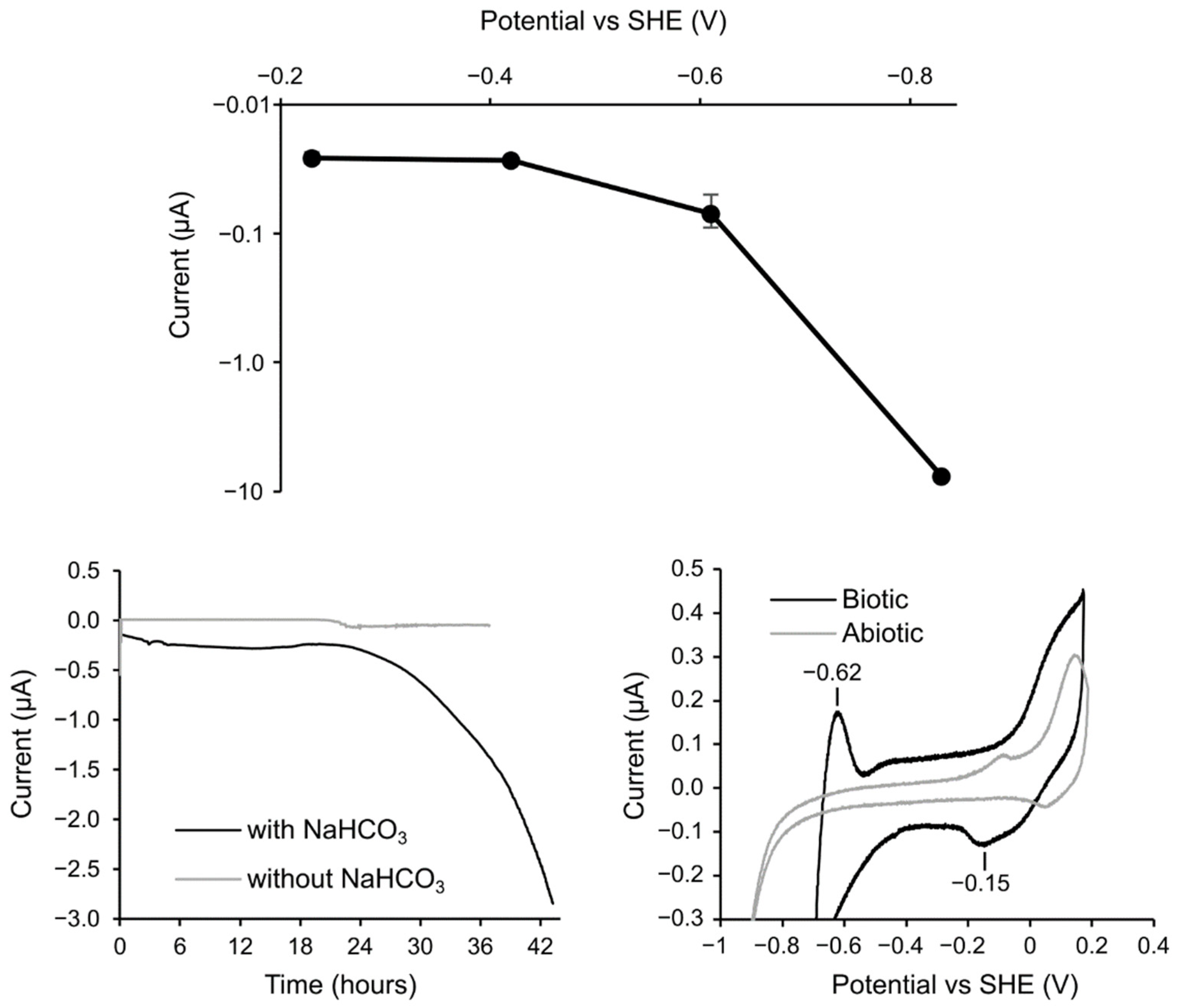
3.3. Electrochemical Incubation and CV Measurement of Methanobacteirum sp. TO1
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Koch, G.; Vaney, J.; Thomopson, N.; Moghissi, O.; Gould, M.; Payer, J. International Measures of Prevention Application, and Economics of Corrosion Technologies Study; NACE Impact: Houston, TX, USA, 2016. [Google Scholar]
- Cord-Ruwisch, R. Microbially Influenced Corrosion of Steel. Environ. Microbe-Met. Interact. 2000, 170, 159–173. [Google Scholar]
- Jia, R.; Unsal, T.; Xu, D.; Lekbach, Y.; Gu, T. Microbiologically Influenced Corrosion and Current Mitigation Strategies: A State of the Art Review. Int. Biodeterior. Biodegrad. 2019, 137, 42–58. [Google Scholar] [CrossRef]
- Dinh, H.T.; Kuever, J.; Mussmann, M.; Hassel, A.W.; Stratmann, M.; Widdel, F. Iron Corrosion by Novel Anaerobic Microorganisms. Nature 2004, 427, 829–832. [Google Scholar] [CrossRef] [PubMed]
- Little, B.; Blackwood, D.; Hinks, J.; Lauro, F.; Marsili, E.; Okamoto, A.; Rice, S.; Wade, S.; Flemming, H.-C. Microbially influenced corrosion—Any progress? Corros. Sci. 2020, 170, 108641. [Google Scholar] [CrossRef]
- Lee, W.; Lewandowski, Z.P.H.; Nielsen, P.H.; Hamilton, W.A. Role of Sulfate-Reducing Bacteria in Corrosion of Mild Steel: A Review. Biofouling 1995, 8, 165–194. [Google Scholar] [CrossRef]
- Enning, D.; Garrelfs, J. Corrosion of Iron by Sulfate-Reducing Bacteria: New Views of an Old Problem. Appl. Environ. Microbiol. 2014, 80, 1226–1236. [Google Scholar] [CrossRef] [Green Version]
- Kato, S.; Yumoto, I.; Kamagata, Y. Isolation of Acetogenic Bacteria That Induce Biocorrosion by Utilizing Metallic Iron as the Sole Electron Donor. Appl. Environ. Microbiol. 2015, 81, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.; Dohmae, N.; Nealson, K.H.; Hashimoto, K.; Okamoto, A. Multi-Heme Cytochromes Provide a Pathway for Survival in Energy-Limited Environments. Sci. Adv. 2018, 4, eaao5682. [Google Scholar] [CrossRef] [Green Version]
- Enning, D.; Venzlaff, H.; Garrelfs, J.; Dinh, H.T.; Meyer, V.; Mayrhofer, K.; Hassel, A.W.; Stratmann, M.; Widdel, F. Marine Sulfate-Reducing Bacteria Cause Serious Corrosion of Iron Under Electroconductive Biogenic Mineral Crust. Environ. Microbiol. 2012, 14, 1772–1787. [Google Scholar] [CrossRef] [Green Version]
- Venzlaff, H.; Enning, D.; Srinivasan, J.; Mayrhofer, K.J.J.; Hassel, A.W.; Widdel, F.; Stratmann, M. Accelerated Cathodic Reaction in Microbial Corrosion of Iron Due to Direct Electron Uptake by Sulfate-Reducing Bacteria. Corros. Sci. 2013, 66, 88–96. [Google Scholar] [CrossRef] [Green Version]
- Uchiyama, T.; Ito, K.; Mori, K.; Tsurumaru, H.; Harayama, S. Iron-Corroding Methanogen Isolated from a Crude-Oil Storage Tank. Appl. Environ. Microbiol. 2010, 76, 1783–1788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palacios, P.A.; Snoeyenbos-West, O.; Löscher, C.R.; Thamdrup, B.; Rotaru, A.E. Baltic Sea Methanogens Compete with Acetogens for Electrons from Metallic Iron. ISME J. 2019, 13, 3011–3023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsurumaru, H.; Ito, N.; Mori, K.; Wakai, S.; Uchiyama, T.; Iino, T.; Hosoyama, A.; Ataku, H.; Nishijima, K.; Mise, M.; et al. An Extracellular [NiFe] Hydrogenase Mediating Iron Corrosion Is Encoded in a Genetically Unstable Genomic Island in Methanococcus maripaludis. Sci. Rep. 2018, 8, 15149. [Google Scholar] [CrossRef] [PubMed]
- Lahme, S.; Mand, J.; Longwell, J.; Smith, R.; Enning, D. Severe Corrosion of Carbon Steel in Oil Field Produced Water Can Be Linked to Methanogenic Archaea Containing a Special Type of [NiFe] Hydrogenase. Appl. Environ. Microbiol. 2021, 87. [Google Scholar] [CrossRef] [PubMed]
- Lovley, D.R.; Phillips, E.J. Rapid Assay for Microbially Reducible Ferric Iron in Aquatic Sediments. Appl. Environ. Microbiol. 1987, 53, 1536–1540. [Google Scholar] [CrossRef] [Green Version]
- Hirano, S.; Nagaoka, T.; Matsumoto, N. Microbial Community Dynamics in a Crust Formed on Carbon Steel SS400 During Corrosion. Corros. Eng. Sci. Technol. 2020, 55, 685–692. [Google Scholar] [CrossRef]
- Skillman, L.C.; Evans, P.N.; Naylor, G.E.; Morvan, B.; Jarvis, G.N.; Joblin, K.N. 16S Ribosomal DNA-Directed PCR Primers for Ruminal Methanogens and Identification of Methanogens Colonising Young Lambs. Anaerobe 2004, 10, 277–285. [Google Scholar] [CrossRef]
- Huerta-Cepas, J.; Serra, F.; Bork, P. ETE 3: Reconstruction, Analysis, and Visualization of Phylogenomic Data. Mol. Biol. Evol. 2016, 33, 1635–1638. [Google Scholar] [CrossRef] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree: Computing large minimum evolution trees with profiles instead of a distance matrix. Mol. Biol. Evol. 2009, 26, 1641–1650. [Google Scholar] [CrossRef]
- Mori, K.; Tsurumaru, H.; Harayama, S. Iron Corrosion Activity of Anaerobic Hydrogen-Consuming Microorganisms Isolated from Oil Facilities. J. Biosci. Bioeng. 2010, 110, 426–430. [Google Scholar] [CrossRef]
- Joulian, C.; Patel, B.K.; Ollivier, B.; Garcia, J.L.; Roger, P.A. Methanobacterium oryzae sp. nov., a Novel Methanogenic Rod Isolated from a Philippines Ricefield. Int. J. Syst. Evol. Microbiol. 2000, 50, 525–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kip, N.; Jansen, S.; Leite, M.F.A.; de Hollander, M.; Afanasyev, M.; Kuramae, E.E.; Veen, J.A.V. Methanogens Predominate in Natural Corrosion Protective Layers on Metal Sheet Piles. Sci. Rep. 2017, 7, 11899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- In ’t Zandt, M.H.; Kip, N.; Frank, J.; Jansen, S.; van Veen, J.A.; Jetten, M.S.M.; Welte, C.U. High-Level Abundances of Methanobacteriales and Syntrophobacterales M. Help To Prevent Corrosion of Metal Sheet Piles. Appl. Environ. Microbiol. 2019, 85. [Google Scholar] [CrossRef] [PubMed]
- An, B.A.; Kleinbub, S.; Ozcan, O.; Koerdt, A. Iron to Gas: Versatile Multiport Flow-Column Revealed Extremely High Corrosion Potential by Methanogen-Induced Microbiologically Influenced Corrosion (Mi-MIC). Front. Microbiol. 2020, 11, 527. [Google Scholar] [CrossRef]
- Beese-Vasbender, P.F.; Grote, J.P.; Garrelfs, J.; Stratmann, M.; Mayrhofer, K.J. Selective Microbial Electrosynthesis of Methane by a Pure Culture of a Marine Lithoautotrophic Archaeon. Bioelectrochemistry 2015, 102, 50–55. [Google Scholar] [CrossRef]
- Deutzmann, J.S.; Sahin, M.; Spormann, A.M. Extracellular Enzymes Facilitate Electron Uptake in Biocorrosion and Bioelectrosynthesis. mBio 2015, 6, e00496-15. [Google Scholar] [CrossRef] [Green Version]
- Lohner, S.T.; Deutzmann, J.S.; Logan, B.E.; Leigh, J.; Spormann, A.M. Hydrogenase-Independent Uptake and Metabolism of Electrons by the Archaeon Methanococcus maripaludis. ISME J. 2014, 8, 1673–1681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowe, A.R.; Xu, S.; Gardel, E.; Bose, A.; Girguis, P.; Amend, J.P.; El-Naggar, M.Y. Methane-Linked Mechanisms of Electron Uptake from Cathodes by Methanosarcina barkeri. mBio 2019, 10, e02448-18. [Google Scholar] [CrossRef] [Green Version]
- Kato, S.; Hashimoto, K.; Watanabe, K. Methanogenesis Facilitated by Electric Syntrophy via (Semi)Conductive Iron–Oxide Minerals. Environ. Microbiol. 2012, 14, 1646–1654. [Google Scholar] [CrossRef]
- Rotaru, A.E.; Calabrese, F.; Stryhanyuk, H.; Musat, F.; Shrestha, P.M.; Weber, H.S.; Snoeyenbos-West, O.L.O.; Hall, P.O.J.; Richnow, H.H.; Musat, N.; et al. Conductive Particles Enable Syntrophic Acetate Oxidation Between Geobacter and Methanosarcina from Coastal Sediments. mBio 2018, 9, e00226-18. [Google Scholar] [CrossRef] [Green Version]
- Lovley, D.R.; Holmes, D.E. Electromicrobiology: The Ecophysiology of Phylogenetically Diverse Electroactive Microorganisms. Nat. Rev. Microbiol. 2022, 20, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Rajala, P.; Carpén, L.; Vepsäläinen, M.; Raulio, M.; Sohlberg, E.; Bomberg, M. Microbially Induced Corrosion of Carbon Steel in Deep Groundwater Environment. Front. Microbiol. 2015, 6, 647. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | pH | ORP (mv) | TOC (mg/L) | COD (mg/L) | Cl (mg/L) | NO2 (mg/L) | Br (mg/L) | NO3 (mg/L) | SO4 (mg/L) |
| GW1 | 8.4 | 71.8 | 2.9 | 4.4 | 10.1 | ND | 0.15 | ND | 10.5 |
| GW2 | 7.1 | −18.9 | 4.4 | 15.2 | 7.5 | 0.022 | ND | 0.28 | 14.2 |
| RW | 8.4 | 14.7 | 2.3 | 2.5 | 5.7 | 0.012 | 0.002 | 1.89 | 18.4 |
| SW | 7.9 | −107.2 | 3.8 | 9.8 | 19,198.2 | ND | 69.4 | ND | 2628.6 |
| Sample Name | Fe (mM) | Li (mg/L) | Na (mg/L) | NH4 (mg/L) | K (mg/L) | Mg (mg/L) | Ca (mg/L) | ||
| GW1 | 41 | ND | 23.5 | 1.2 | 7.6 | 4.5 | 14.3 | ||
| GW2 | 32 | ND | 9.1 | 3.0 | 2.8 | 3.1 | 7.5 | ||
| RW | 5 | ND | 9.2 | 0.54 | 1.7 | 4.1 | 15.3 | ||
| SW | 23 | 1.78 | 10,293.4 | ND | 543.3 | 1303.5 | 395.5 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hirano, S.-i.; Ihara, S.; Wakai, S.; Dotsuta, Y.; Otani, K.; Kitagaki, T.; Ueno, F.; Okamoto, A. Novel Methanobacterium Strain Induces Severe Corrosion by Retrieving Electrons from Fe0 under a Freshwater Environment. Microorganisms 2022, 10, 270. https://doi.org/10.3390/microorganisms10020270
Hirano S-i, Ihara S, Wakai S, Dotsuta Y, Otani K, Kitagaki T, Ueno F, Okamoto A. Novel Methanobacterium Strain Induces Severe Corrosion by Retrieving Electrons from Fe0 under a Freshwater Environment. Microorganisms. 2022; 10(2):270. https://doi.org/10.3390/microorganisms10020270
Chicago/Turabian StyleHirano, Shin-ichi, Sota Ihara, Satoshi Wakai, Yuma Dotsuta, Kyohei Otani, Toru Kitagaki, Fumiyoshi Ueno, and Akihiro Okamoto. 2022. "Novel Methanobacterium Strain Induces Severe Corrosion by Retrieving Electrons from Fe0 under a Freshwater Environment" Microorganisms 10, no. 2: 270. https://doi.org/10.3390/microorganisms10020270