
Minor Impact of A258D Mutation on Biochemical and Enzymatic Properties of Leishmania infantum GDP-Mannose Pyrophosphorylase

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Media
2.2. Identification of Leishmanial GDP-MP Sequences
2.3. DNA Extraction, PCR and Cloning Strategy
2.4. Production of Recombinant LiGDP-MP
2.5. Purification of Recombinant LiGDP-MP
2.6. Enzymatic Assays
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Leishmaniasis Fact Sheet, 20 May 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/leishmaniasis (accessed on 29 October 2021).
- Franssen, S.U.; Durrant, C.; Stark, O.; Moser, B.; Downing, T.; Imamura, H.; Dujardin, J.-C.; Sanders, M.J.; Mauricio, I.; Miles, M.A.; et al. Global genome diversity of the Leishmania donovani complex. Elife 2020, 9, 51243. [Google Scholar] [CrossRef]
- Ready, P. Epidemiology of visceral leishmaniasis. Clin. Epidemiol. 2014, 6, 147–154. [Google Scholar] [CrossRef] [Green Version]
- Maia, C.; Cardoso, L. Spread of Leishmania infantum in Europe with dog travelling. Vet. Parasitol. 2015, 213, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Le Rutte, E.A.; van Straten, R.; Overgaauw, P.A. Awareness and control of canine leishmaniosis: A survey among Spanish and French veterinarians. Vet. Parasitol. 2018, 253, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Roatt, B.M.; Cardoso, J.M.D.O.; De Brito, R.C.F.; Coura-Vital, W.; Aguiar-Soares, R.D.D.O.; Reis, A.B. Recent advances and new strategies on leishmaniasis treatment. Appl. Microbiol. Biotechnol. 2020, 104, 8965–8977. [Google Scholar] [CrossRef]
- Freeze, H.H.; Elbein, A.D. Glycosylation precursors. In Essentials of Glycobiology, 2nd ed.; Varki, A., Cummings, R.D., Esko, J.D., Freeze, H.H., Stanley, P., Bertozzi, C.R., Hart, G.W., Etzler, M.E., Eds.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2009; pp. 47–61. [Google Scholar]
- Pomel, S.; Loiseau, P.M. GDP-mannose: A key-point for target identification and drug design in kinetoplastids. In Trypanosomatid Diseases: Molecular Routes to Drug Discoveries; Jäger, T., Koch, O., Flohe, L., Eds.; Wiley-VCH Verlag GmbH and Co. KGaA: Weinheim, Germany, 2013; pp. 315–334. [Google Scholar]
- Garami, A.R.; Ilg, T. Disruption of mannose activation in Leishmania mexicana: GDP-mannose pyrophosphorylase is required for virulence, but not for viability. EMBO J. 2001, 20, 3657–3666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, J.; Curtis, J.; Spurck, T.P.; Ilg, T.; Garami, A.R.; Baldwin, T.; Courret, N.; McFadden, G.I.; Davis, A.; Handman, E. Characterisation of a Leishmania mexicana knockout lacking guanosine diphosphate-mannose pyrophosphorylase. Int. J. Parasitol. 2005, 35, 861–873. [Google Scholar] [CrossRef]
- Pomel, S.; Rodrigo, J.; Hendra, F.; Cavé, C.; Loiseau, P. In silicoanalysis of a therapeutic target in Leishmania infantum: The guanosine-diphospho-D-mannose pyrophosphorylase. Parasite 2012, 19, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Daligaux, P.; Bernadat, G.; Tran, L.; Cavé, C.; Loiseau, P.M.; Pomel, S.; Ha-Duong, T. Comparative study of structural models of Leishmania donovani and human GDP-mannose pyrophosphorylases. Eur. J. Med. Chem. 2016, 107, 109–118. [Google Scholar] [CrossRef]
- Levaique, H.; Pamlard, O.; Apel, C.; Bignon, J.; Arriola, M.; Kuhner, R.; Awang, K.; Loiseau, P.; Litaudon, M.; Pomel, S. Alkyl-resorcinol derivatives as inhibitors of GDP-mannose pyrophosphorylase with antileishmanial activities. Molecules 2021, 26, 1551. [Google Scholar] [CrossRef]
- Carnielli, J.B.; de Andrade, H.M.; Pires, S.F.; Chapeaurouge, A.D.; Perales, J.; Monti-Rocha, R.; Carvalho, S.F.; Ribeiro, L.P.; Dietze, R.; Figueiredo, S.G.; et al. Proteomic analysis of the soluble proteomes of miltefosine-sensitive and -resistant Leishmania infantum chagasi isolates obtained from Brazilian patients with different treatment outcomes. J. Proteom. 2014, 108, 198–208. [Google Scholar] [CrossRef] [Green Version]
- Pountain, A.W.; Barrett, M.P. Untargeted metabolomics to understand the basis of phenotypic differences in amphotericin B-resistant Leishmania parasites. Wellcome Open Res. 2019, 4, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, W.; Daligaux, P.; Lazar, N.; Ha-Duong, T.; Cavé, C.; Van Tilbeurgh, H.; Loiseau, P.; Pomel, S. Biochemical analysis of leishmanial and human GDP-Mannose Pyrophosphorylases and selection of inhibitors as new leads. Sci. Rep. 2017, 7, 751. [Google Scholar] [CrossRef] [Green Version]
- Pomel, S.; Mao, W.; Ha-Duong, T.; Cavé, C.; Loiseau, P. GDP-mannose pyrophosphorylase: A biologically validated target for drug development against leishmaniasis. Front. Cell. Infect. Microbiol. 2019, 9, 186. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Pelissier, M.-C.; Lesley, S.A.; Kuhn, P.; Bourne, Y. Structural insights into the catalytic mechanism of bacterial guanosine-diphospho-d-mannose pyrophosphorylase and its regulation by divalent ions. J. Biol. Chem. 2010, 285, 27468–27476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, L.; Liu, Z.; Wang, Y.; Yang, F.; Wang, J.; Huang, W.; Qin, J.; Tian, M.; Cai, X.; Liu, X.; et al. Cryo-EM structures of human GMPPA–GMPPB complex reveal how cells maintain GDP-mannose homeostasis. Nat. Struct. Mol. Biol. 2021, 28. [Google Scholar] [CrossRef]
- Rudolf, F.B.; Fromm, H.J. Plotting methods for analyzing enzyme rate data. In Contemporary Enzyme Kinetics and Mechanism; Purich, D.L., Ed.; Academic Press: San Diego, CA, USA, 1983; pp. 53–74. [Google Scholar]
- Davis, A.J.; Perugini, M.; Smith, B.; Stewart, J.; Ilg, T.; Hodder, A.N.; Handman, E. Properties of GDP-mannose pyrophosphorylase, a critical enzyme and drug target in Leishmania mexicana. J. Biol. Chem. 2004, 279, 12462–12468. [Google Scholar] [CrossRef] [Green Version]
- Szumilo, T.; Drake, R.; York, J.; Elbein, A. GDP-mannose pyrophosphorylase. Purification to homogeneity, properties, and utilization to prepare photoaffinity analogs. J. Biol. Chem. 1993, 268, 17943–17950. [Google Scholar] [CrossRef]
- Ning, B.; Elbein, A.D. Cloning, expression and characterization of the pig liver GDP-mannose pyrophosphorylase. Evidence that GDP-mannose and GDP-Glc pyrophosphorylases are different proteins. Eur. J. Biochem. 2000, 267, 6866–6874. [Google Scholar] [CrossRef] [PubMed]
- Führing, J.; Damerow, S.; Fedorov, R.; Schneider, J.; Münster-Kühnel, A.-K.; Gerardy-Schahn, R. Octamerization is essential for enzymatic function of human UDP-glucose pyrophosphorylase. Glycobiology 2013, 23, 426–437. [Google Scholar] [CrossRef] [Green Version]
- Führing, J.I.; Cramer, J.T.; Schneider, J.; Baruch, P.; Gerardy-Schahn, R.; Fedorov, R. A quaternary mechanism enables the complex biological functions of octameric human UDP-glucose pyrophosphorylase, a key enzyme in cell metabolism. Sci. Rep. 2015, 5, 9618. [Google Scholar] [CrossRef] [Green Version]
- Ning, B.; Elbein, A. Purification and properties of mycobacterial GDP–mannose pyrophosphorylase. Arch. Biochem. Biophys. 1999, 362, 339–345. [Google Scholar] [CrossRef]
- Denton, H.; Fyffe, S.; Smith, T.K. GDP-mannose pyrophosphorylase is essential in the bloodstream form of Trypanosoma brucei. Biochem. J. 2010, 425, 603–614. [Google Scholar] [CrossRef] [Green Version]
- Shinabarger, D.; Berry, A.; May, T.B.; Rothmel, R.; Fialho, A.; Chakrabarty, A.M. Purification and characterization of phosphomannose isomerase-guanosine diphospho-D-mannose pyrophosphorylase. A bifunctional enzyme in the alginate biosynthetic pathway of Pseudomonas aeruginosa. J. Biol. Chem. 1991, 266, 2080–2088. [Google Scholar] [CrossRef]
- May, T.; Shinabarger, D.; Boyd, A.; Chakrabarty, A. Identification of amino acid residues involved in the activity of phosphomannose isomerase-guanosine 5′-diphospho-D-mannose pyrophosphorylase. A bifunctional enzyme in the alginate biosynthetic pathway of Pseudomonas aeruginosa. J. Biol. Chem. 1994, 269, 4872–4877. [Google Scholar] [CrossRef]
- Elling, L.; Ritter, J.E.; Verseck, S. Expression, purification and characterization of recombinant phosphomannomutase and GDP-α-D-mannose pyrophosphorylase from Salmonella enterica, group B, for the synthesis of GDP-α-D-mannose from D-mannose. Glycobiology 1996, 6, 591–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fey, S.; Elling, L.; Kragl, U. The cofactor Mg2+—A key switch for effective continuous enzymatic production of GDP-mannose using recombinant GDP-mannose pyrophosphorylase. Carbohydr. Res. 1997, 305, 475–481. [Google Scholar] [CrossRef]
- Zmuda, F.; Shepherd, S.M.; Ferguson, M.; Gray, D.W.; Torrie, L.S.; De Rycker, M. Trypanosoma cruzi Phosphomannomutase and Guanosine Diphosphate-Mannose Pyrophosphorylase Ligandability Assessment. Antimicrob. Agents Chemother. 2019, 63, e01082-19. [Google Scholar] [CrossRef] [Green Version]
- Mizanur, R.M.; Pohl, N.L.B. Phosphomannose isomerase/GDP-mannose pyrophosphorylase from Pyrococcus furiosus: A thermostable biocatalyst for the synthesis of guanidinediphosphate-activated and mannose-containing sugar nucleotides. Org. Biomol. Chem. 2009, 7, 2135–2139. [Google Scholar] [CrossRef]
- Zou, L.; Zheng, R.B.; Lowary, T.L. Studies on the substrate specificity of a GDP-mannose pyrophosphorylase from Salmonella enterica. Beilstein J. Org. Chem. 2012, 8, 1219–1226. [Google Scholar] [CrossRef] [PubMed]
- Lindquist, L.; Kaiser, R.; Reeves, P.R.; Lindberg, A.A. Purification, characterization and HPLC assay of Salmonella glucose-1-phosphate thymidylyltransferase from the cloned rfbA gene. JBIC J. Biol. Inorg. Chem. 1993, 211, 763–770. [Google Scholar] [CrossRef] [PubMed]
- Boehlein, S.K.; Shaw, J.R.; Hwang, S.K.; Stewart, J.D.; Hannah, L.C. Deciphering the kinetic mechanisms controlling selected plant ADP-glucose pyrophosphorylases. Arch. Biochem. Biophys. 2013, 535, 215–226. [Google Scholar] [CrossRef] [PubMed]
- A Barton, W.; Lesniak, J.; Biggins, J.B.; Jeffrey, P.D.; Jiang, J.; Rajashankar, K.R.; Thorson, J.S.; Nikolov, D.B. Structure, mechanism and engineering of a nucleotidylyltransferase as a first step toward glycorandomization. Nat. Struct. Biol. 2001, 8, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Zuccotti, S.; Zanardi, D.; Rosano, C.; Sturla, L.; Tonetti, M.; Bolognesi, M. Kinetic and crystallographic analyses support a sequential-ordered bi bi catalytic mechanism for Escherichia coli glucose-1-phosphate thymidylyltransferase. J. Mol. Biol. 2001, 313, 831–843. [Google Scholar] [CrossRef]
- Persat, F.; Azzar, G.; Martel, M.-B.; Got, R. Properties of uridine diphosphate glucose pyrophosphorylase from Golgi apparatus of liver. Biochim. Biophys. Acta 1983, 749, 329–332. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
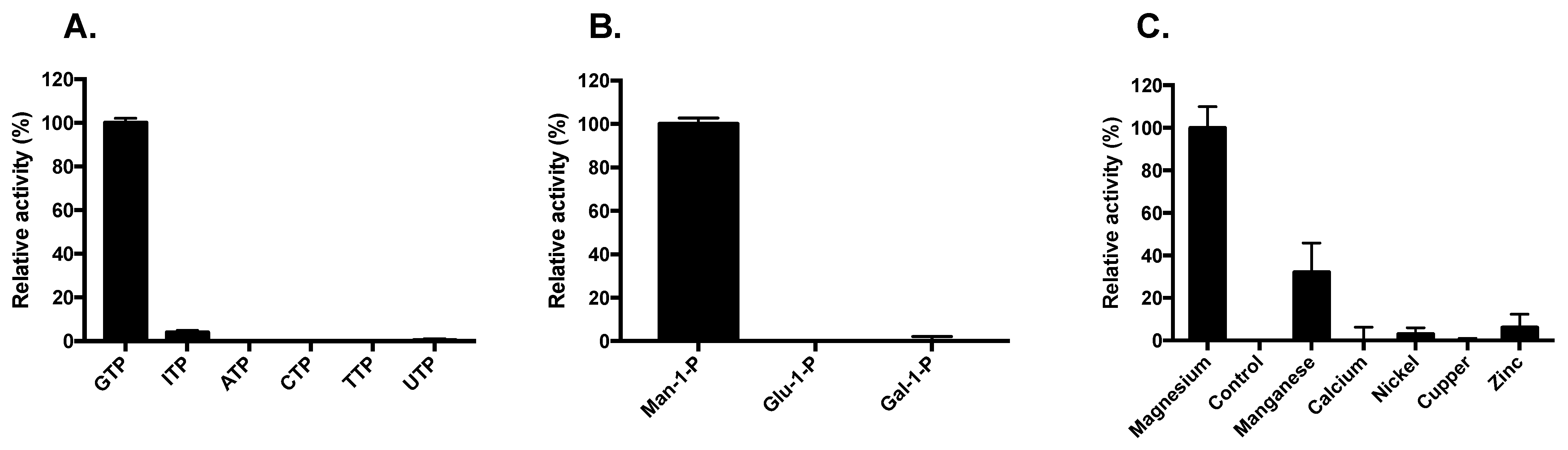{kind=link}
{kind=link}
| Substrate | Vm (µM·min−1) ± SD | Km (µM) ± SD | kcat (min−1) ± SD | kcat/Km (min−1µM−1) ± SD | Reference | |
|---|---|---|---|---|---|---|
| LiGDP-MP | Man-1-P | 0.60 ± 0.02 | 14.72 ± 1.45 | 12.47 ± 0.47 | 0.85 ± 0.06 | This study |
| GTP | 0.63 ± 0.02 | 17.95 ± 1.61 | 13.13 ± 0.43 | 0.73 ± 0.04 | ||
| LdGDP-MP | Man-1-P | 0.30 ± 0.06 | 9.98 ± 6.15 | 6.23 ± 1.24 | 0.75 ± 0.33 | [14] |
| GTP | 0.39 ± 0.03 | 16.84 ± 3.29 | 8.05 ± 0.71 | 0.48 ± 0.05 | ||
| hGDP-MP | Man-1-P | 0.47 ± 0.01 | 6.98 ± 0.64 | 9.30 ± 0.19 | 1.34 ± 0.11 | [14] |
| GTP | 0.44 ± 0.01 | 7.08 ± 0.57 | 8.74 ± 0.14 | 1.24 ± 0.10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mao, W.; Lazar, N.; van Tilbeurgh, H.; Loiseau, P.M.; Pomel, S. Minor Impact of A258D Mutation on Biochemical and Enzymatic Properties of Leishmania infantum GDP-Mannose Pyrophosphorylase. Microorganisms 2022, 10, 231. https://doi.org/10.3390/microorganisms10020231
Mao W, Lazar N, van Tilbeurgh H, Loiseau PM, Pomel S. Minor Impact of A258D Mutation on Biochemical and Enzymatic Properties of Leishmania infantum GDP-Mannose Pyrophosphorylase. Microorganisms. 2022; 10(2):231. https://doi.org/10.3390/microorganisms10020231
Chicago/Turabian StyleMao, Wei, Noureddine Lazar, Herman van Tilbeurgh, Philippe M. Loiseau, and Sébastien Pomel. 2022. "Minor Impact of A258D Mutation on Biochemical and Enzymatic Properties of Leishmania infantum GDP-Mannose Pyrophosphorylase" Microorganisms 10, no. 2: 231. https://doi.org/10.3390/microorganisms10020231