
The Antimicrobial Effect of Various Single-Strain and Multi-Strain Probiotics, Dietary Supplements or Other Beneficial Microbes against Common Clinical Wound Pathogens


Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbial Strains and Clinical Isolates
2.2. Molecular Methods for the Detection of Probiotic Strains
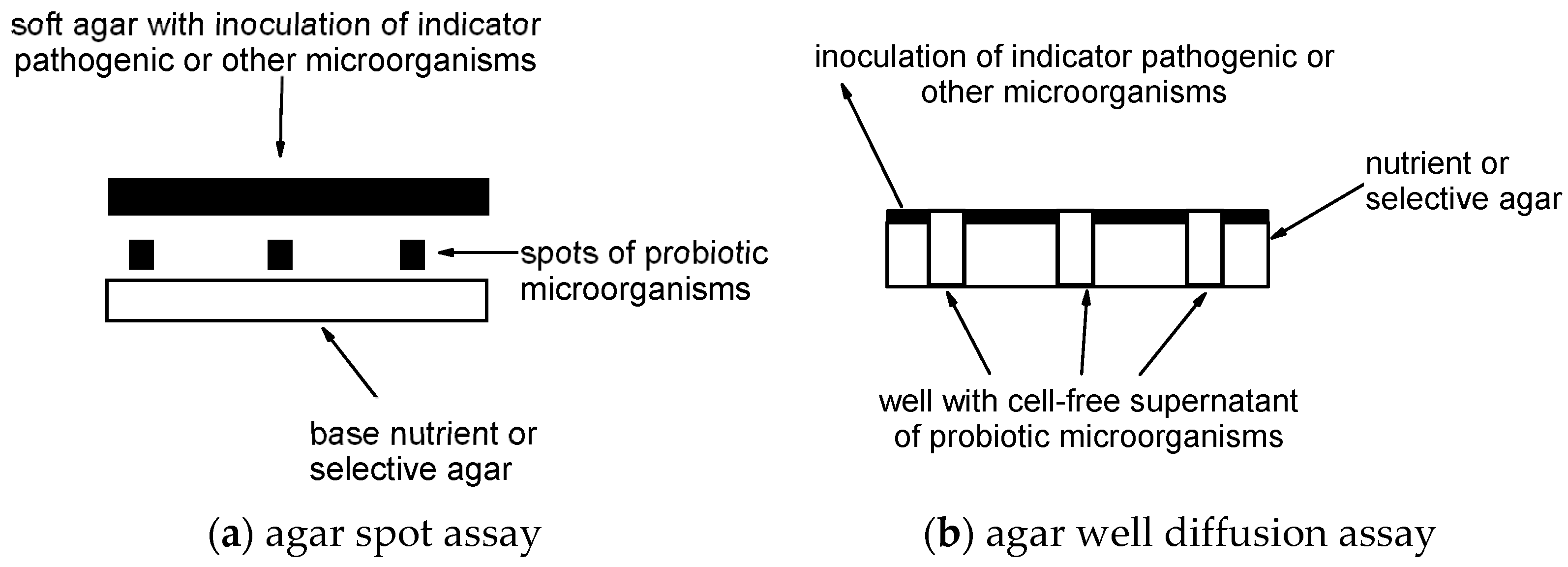
2.3. Agar Spot Assay
2.4. Co-Culturing for Microbial Competition Assay
2.5. Agar Well Diffusion Assay
2.6. Statistics
3. Results
3.1. Identification of Species and Genera of Microbial Strains Using Molecular Detection Methods
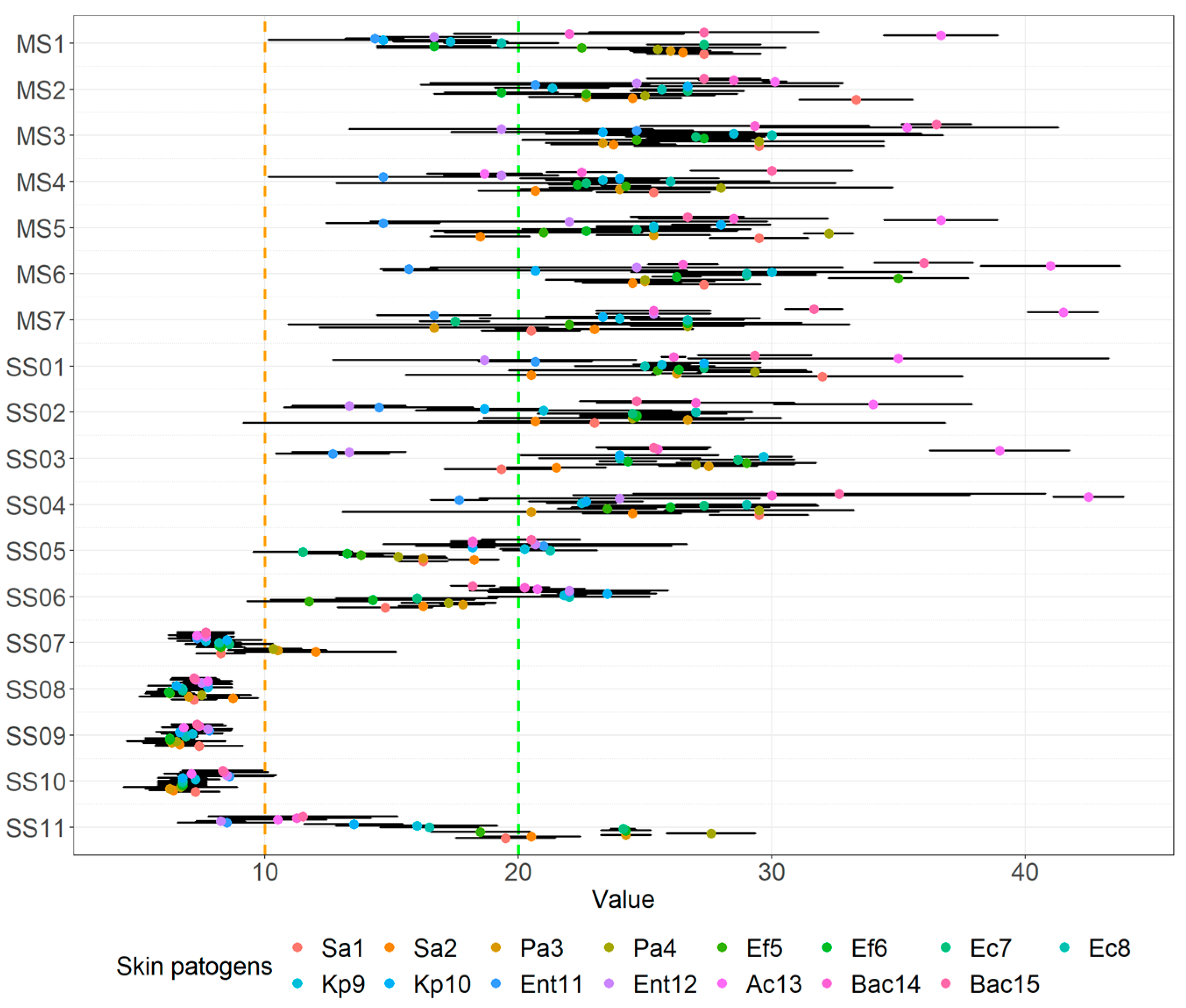
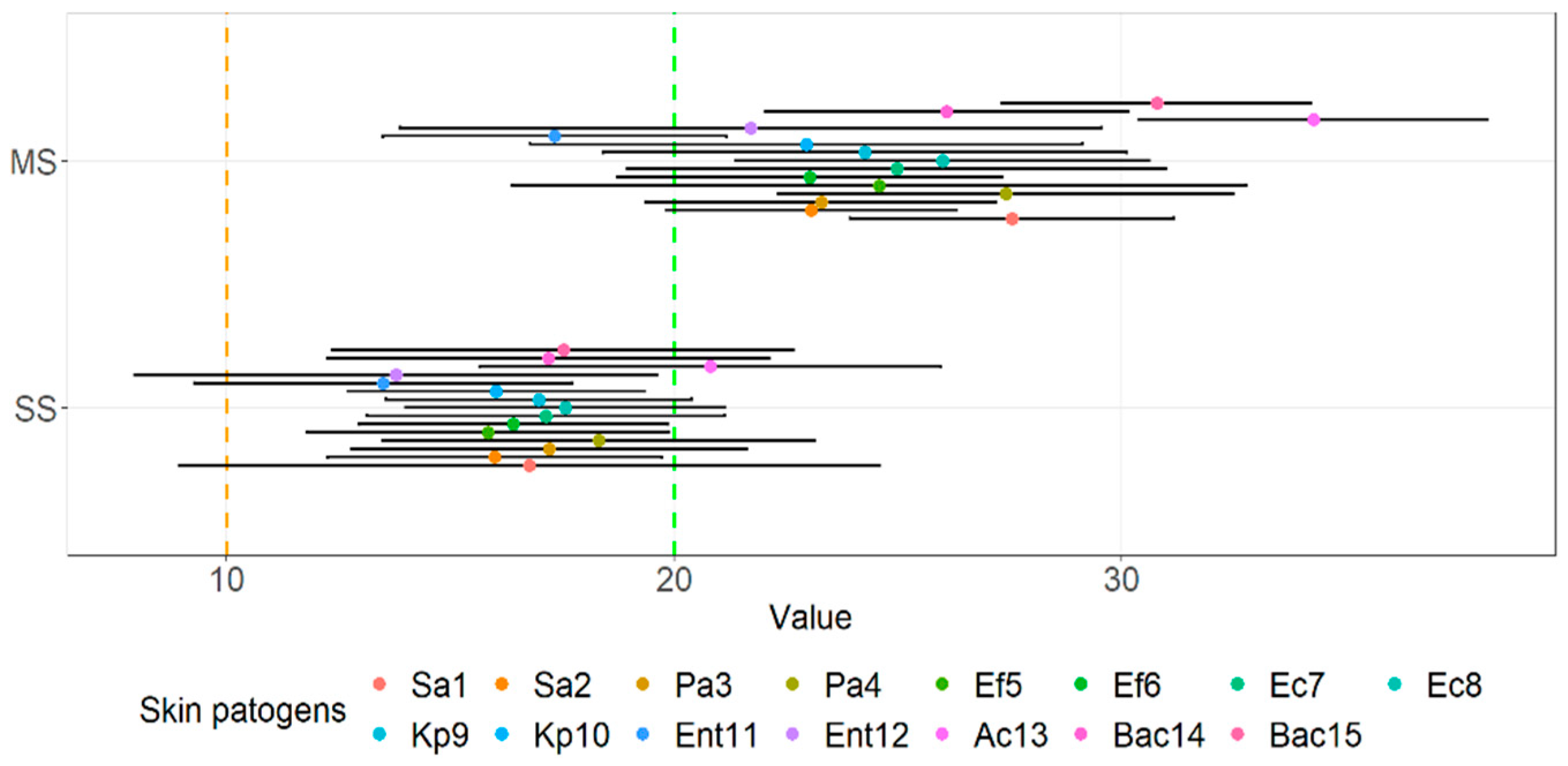
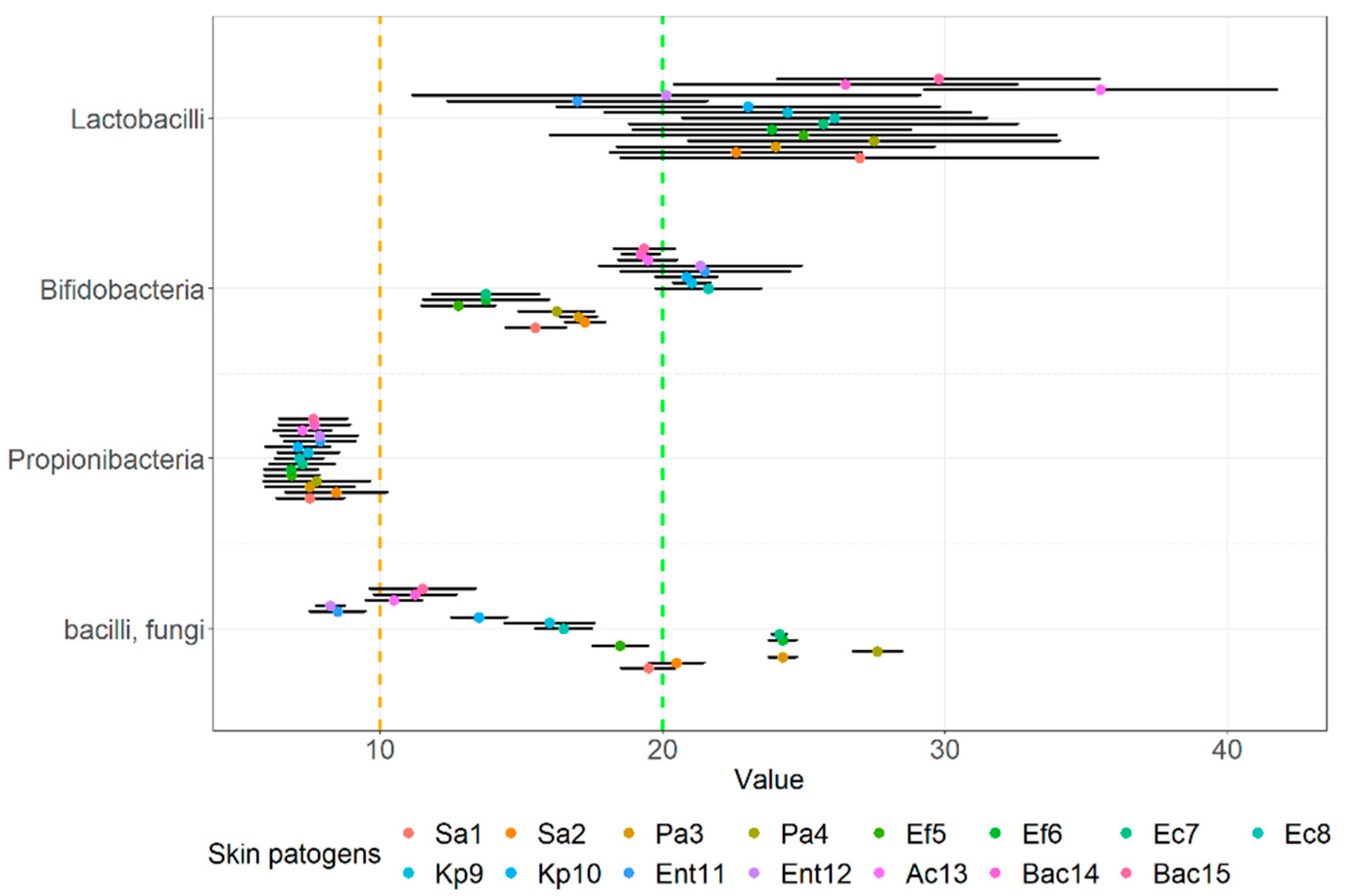
3.2. Agar Spot Assay
3.3. Co-Culturing for Microbial Competition Assay
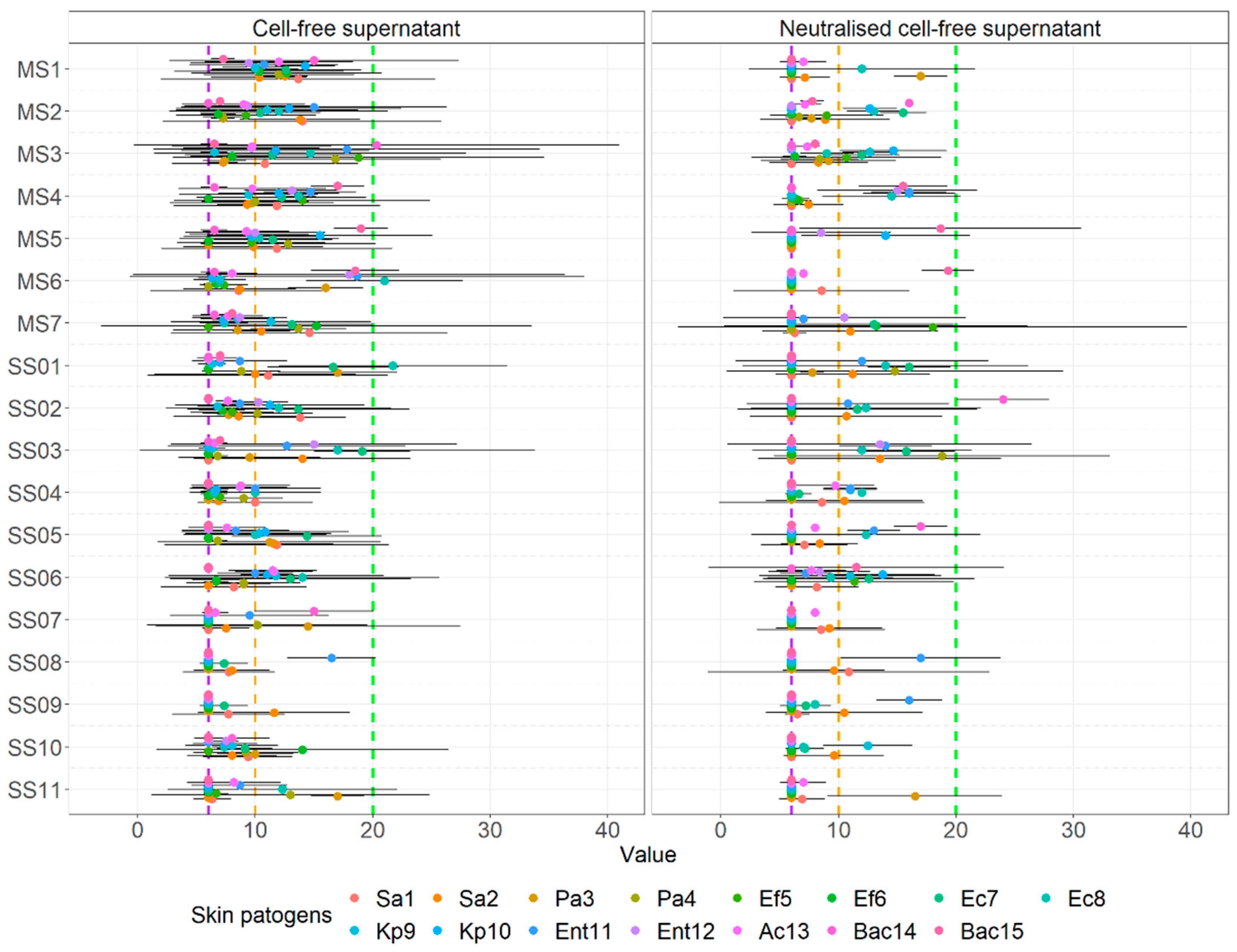
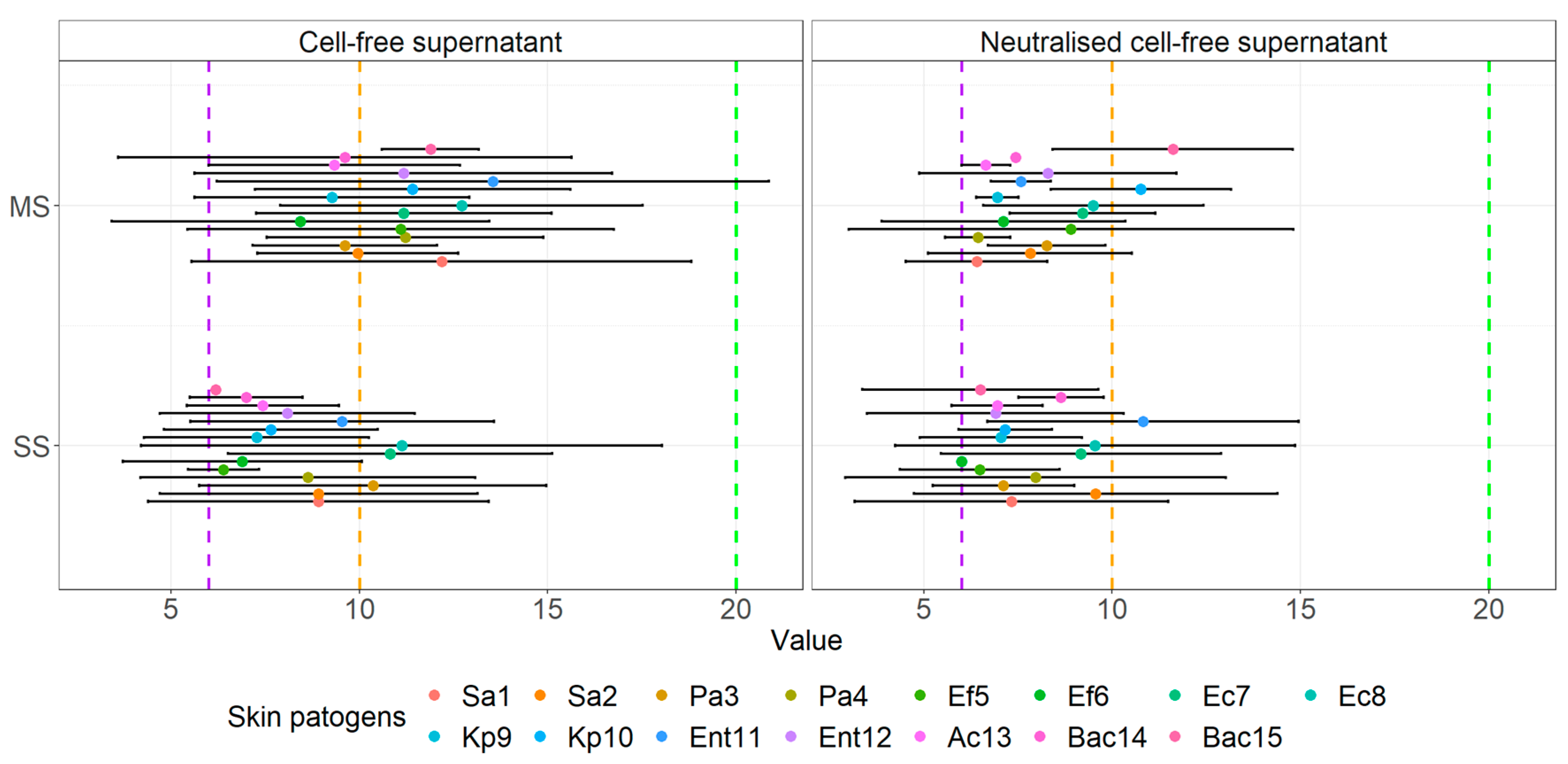
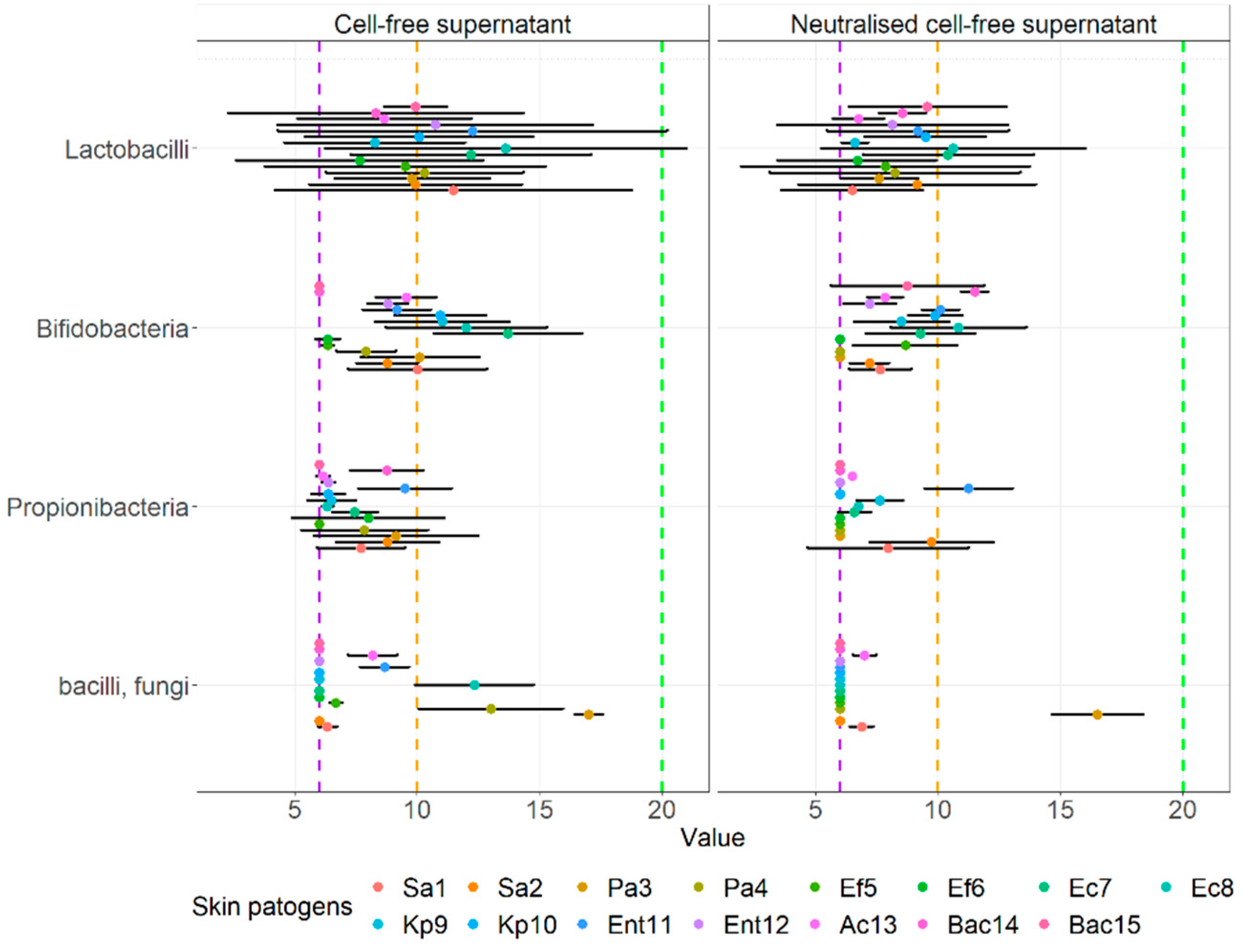
3.4. Agar Well Diffusion Assay
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- McLoughlin, I.J.; Wright, E.M.; Tagg, J.R.; Jain, R.; Hale, J.D.F. Skin microbiome—The next frontier for probiotic intervention. Probiotics Antimicrob. Proteins 2022, 14, 630–647. [Google Scholar] [CrossRef] [PubMed]
- Perez Perez, G.I.; Gao, Z.; Jourdain, R.; Ramirez, J.; Gany, F.; Clavaud, C.; Demaude, J.; Breton, L.; Blaser, M.J. Body site is a more determinant factor than human population diversity in the healthy skin microbiome. PLoS ONE 2016, 11, e0151990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mihai, M.M.; Preda, M.; Lungu, I.; Gestal, M.C.; Popa, M.I.; Holban, A.M. Nanocoatings for chronic wound repair—Modulation of microbial colonization and biofilm formation. Int. J. Mol. Sci. 2018, 19, 1179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scalise, A.; Bianchi, A.; Tartaglione, C.; Bolletta, E.; Pierangeli, M.; Torresetti, M.; Marazzi, M.; Di Benedetto, G. Microenvironment and microbiology of skin wounds: The role of bacterial biofilms and related factors. Semin. Vasc. Surg. 2015, 28, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Peral, M.C.; Huaman Martinez, M.A.; Valdez, J.C. Bacteriotherapy with Lactobacillus plantarum in burns. Int. Wound J. 2009, 6, 73–81. [Google Scholar] [CrossRef]
- Daeschlein, G. Antimicrobial and antiseptic strategies in wound management. Int. Wound J. 2013, 10, 9–14. [Google Scholar] [CrossRef]
- Bowler, P.G.; Duerden, B.I.; Armstrong, D.G. Wound microbiology and associated approaches to wound management. Clin. Microbiol. Rev. 2001, 14, 244–269. [Google Scholar] [CrossRef] [Green Version]
- Mahnic, A.; Breznik, V.; Bombek Ihan, M.; Rupnik, M. Comparison between cultivation and sequencing based approaches for microbiota analysis in swabs and biopsies of chronic wounds. Front. Med. 2021, 8, 607255. [Google Scholar] [CrossRef]
- Fijan, S.; Frauwallner, A.; Langerholc, T.; Krebs, B.; Ter Haar Née Younes, J.A.; Heschl, A.; Mičetić Turk, D.; Rogelj, I. Efficacy of using probiotics with antagonistic activity against pathogens of wound infections: An integrative review of literature. Biomed Res. Int. 2019, 2019, 7585486. [Google Scholar] [CrossRef] [Green Version]
- Martinengo, L.; Olsson, M.; Bajpai, R.; Soljak, M.; Upton, Z.; Schmidtchen, A.; Car, J.; Järbrink, K. Prevalence of chronic wounds in the general population: Systematic review and meta-analysis of observational studies. Ann. Epidemiol. 2019, 29, 8–15. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The international scientific association for probiotics and prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Reid, G.; Younes, J.A.; Van der Mei, H.C.; Gloor, G.B.; Knight, R.; Busscher, H.J. Microbiota restoration: Natural and supplemented recovery of human microbial communities. Nat. Rev. Microbiol. 2011, 9, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Cecchini, M.; Langer, J.; Slawomirski, L. Antimicrobial Resistance in G7 Countries and beyond,” Economic Issues, Policies and Options for Action: Oecd Report; OECD: Paris, France, 2015. [Google Scholar]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The international scientific association of probiotics and prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef] [PubMed]
- Ramos, A.N.; Sesto Cabral, M.E.; Noseda, D.; Bosch, A.; Yantorno, O.M.; Valdez, J.C. Antipathogenic properties of Lactobacillus plantarum on Pseudomonas aeruginosa: The potential use of its supernatants in the treatment of infected chronic wounds. Wound Repair Regen. 2012, 20, 552–562. [Google Scholar] [CrossRef]
- Valdéz, J.C.; Peral, M.C.; Rachid, M.; Santana, M.; Perdigón, G. Interference of Lactobacillus plantarum with Pseudomonas aeruginosa in vitro and in infected burns: The potential use of probiotics in wound treatment. Clin. Microbiol. Infect. 2005, 11, 472–479. [Google Scholar] [CrossRef] [Green Version]
- Jones, M.L.; Ganopolsky, J.G.; Labbé, A.; Prakash, S. A novel nitric oxide producing probiotic patch and its antimicrobial efficacy: Preparation and in vitro analysis. Appl. Microbiol. Biotechnol. 2010, 87, 509–516. [Google Scholar] [CrossRef]
- Chan, A.P.; Choi, Y.; Brinkac, L.M.; Krishnakumar, R.; Depew, J.; Kim, M.; Hinkle, M.K.; Lesho, E.P.; Fouts, D.E. Multidrug resistant pathogens respond differently to the presence of co-pathogen, commensal, probiotic and host cells. Sci. Rep. 2018, 8, 8656. [Google Scholar] [CrossRef]
- Mohammedsaeed, W.; McBain, A.J.; Cruickshank, S.M.; O’Neill, C.A.; Schaffner, D.W. Lactobacillus rhamnosus GG inhibits the toxic effects of Staphylococcus aureus on epidermal keratinocytes. Appl. Environ. Microbiol. 2014, 80, 5773–5781. [Google Scholar] [CrossRef] [Green Version]
- Shu, M.; Wang, Y.; Yu, J.; Kuo, S.; Coda, A.; Jiang, Y.; Gallo, R.L.; Huang, C.-M. Fermentation of Propionibacterium acnes, a commensal bacterium in the human skin microbiome, as skin probiotics against methicillin-resistant Staphylococcus aureus. PLoS ONE 2013, 8, e55380. [Google Scholar] [CrossRef]
- Al-Malkey, M.K.; Ismeeal, M.C.; Al-Hur, F.J.; Mohammed, S.W.; Nayyel, H.J. Antimicrobial effect of probiotic Lactobacillus spp. on Pseudomonas aeruginosa. J. Contemp. Med. Sci. 2017, 3, 218–223. [Google Scholar] [CrossRef]
- Prince, T.; McBain, A.J.; O’Neill, C.A. Lactobacillus reuteri protects epidermal keratinocytes from Staphylococcus aureus-induced cell death by competitive exclusion. Appl. Environ. Microbiol. 2012, 78, 5119–5126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, J.G.; Motlagh, H.; Povey, S.B.; Percival, S.L. 2—The role of micro-organisms and biofilms in dysfunctional wound healing. In Advanced Wound Repair Therapies; Farrar, D., Ed.; Woodhead Publishing: Sawston, UK, 2011; pp. 39–76. [Google Scholar]
- Lopes, E.G.; Moreira, D.A.; Gullón, P.; Gullón, B.; Cardelle-Cobas, A.; Tavaria, F.K. Topical application of probiotics in skin: Adhesion, antimicrobial and antibiofilm in vitro assays. J. Appl. Microbiol. 2017, 122, 450–461. [Google Scholar] [CrossRef]
- Moghadam, S.S.; Khodaii, Z.; Zadeh, S.F.; Ghooshchian, M.; Aghmiyuni, Z.F.; Shabestari, T.M. Synergistic or antagonistic effects of probiotics and antibiotics- alone or in combination- on antimicrobial-resistant Pseudomonas aeruginosa isolated from burn wounds. Arch. Clin. Infect. Dis. 2018, 13, e63121. [Google Scholar] [CrossRef] [Green Version]
- Argenta, A.; Satish, L.; Gallo, P.; Liu, F.; Kathju, S. Local application of probiotic bacteria prophylaxes against sepsis and death resulting from burn wound infection. PLoS ONE 2016, 11, e0165294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satish, L.; Gallo, P.H.; Johnson, S.; Yates, C.C.; Kathju, S. Local probiotic therapy with Lactobacillus plantarum mitigates scar formation in rabbits after burn injury and infection. Surg. Infect. 2017, 18, 119–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sürmeli, M.; Maçin, S.; Akyön, Y.; Kayikçioğlu, A.U. The protective effect of Lactobacillus plantarum against meticillin-resistant Staphylococcus aureus infections: An experimental animal model. J. Wound Care 2019, 28, s29–s34. [Google Scholar] [CrossRef]
- Brachkova, M.I.; Marques, P.; Rocha, J.; Sepodes, B.; Duarte, M.A.; Pinto, J.F. Alginate films containing Lactobacillus plantarum as wound dressing for prevention of burn infection. J. Hosp. Infect. 2011, 79, 375–377. [Google Scholar] [CrossRef]
- Ong, J.S.; Taylor, T.D.; Yong, C.C.; Khoo, B.Y.; Sasidharan, S.; Choi, S.B.; Ohno, H.; Liong, M.T. Lactobacillus plantarum USM8613 aids in wound healing and suppresses Staphylococcus aureus infection at wound sites. Probiotics Antimicrob. Proteins 2020, 12, 125–137. [Google Scholar] [CrossRef]
- Jones, M.; Ganopolsky, J.G.; Labbé, A.; Gilardino, M.; Wahl, C.; Martoni, C.; Prakash, S. Novel nitric oxide producing probiotic wound healing patch: Preparation and in vivo analysis in a new zealand white rabbit model of ischaemic and infected wounds. Int. Wound J. 2012, 9, 330–343. [Google Scholar] [CrossRef]
- Binda, S.; Hill, C.; Johansen, E.; Obis, D.; Pot, B.; Sanders, M.E.; Tremblay, A.; Ouwehand, A.C. Criteria to qualify microorganisms as probiotic in foods and dietary supplements. Front. Microbiol. 2020, 11, 1662. [Google Scholar] [CrossRef]
- Peral, M.C.; Rachid, M.M.; Gobbato, N.M.; Martinez, M.A.H.; Valdez, J.C. Interleukin-8 production by polymorphonuclear leukocytes from patients with chronic infected leg ulcers treated with Lactobacillus plantarum. Clin. Microbiol. Infect. 2010, 16, 281–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komatsu, S.; Sakamoto, E.; Norimizu, S.; Shingu, Y.; Asahara, T.; Nomoto, K.; Nagino, M. Efficacy of perioperative synbiotics treatment for the prevention of surgical site infection after laparoscopic colorectal surgery: A randomized controlled trial. Surg. Today 2016, 46, 479–490. [Google Scholar] [CrossRef] [PubMed]
- El-Ghazely, M.H.; Mahmoud, W.H.; Atia, M.A.; Eldip, E.M. Effect of probiotic administration in the therapy of pediatric thermal burn. Ann. Burn. Fire Disasters 2016, 29, 268–272. [Google Scholar]
- Mayes, T.; Gottschlich, M.M.; James, L.E.; Allgeier, C.; Weitz, J.; Kagan, R.J. Clinical safety and efficacy of probiotic administration following burn injury. J. Burn Care Res. 2015, 36, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Kotzampassi, K.; Stavrou, G.; Damoraki, G.; Georgitsi, M.; Basdanis, G.; Tsaousi, G.; Giamarellos-Bourboulis, E.J. A four-probiotics regimen reduces postoperative complications after colorectal surgery: A randomized, double-blind, placebo-controlled study. World J. Surg. 2015, 39, 2776–2783. [Google Scholar] [CrossRef]
- Aisu, N.; Tanimura, S.; Yamashita, Y.; Yamashita, K.; Maki, K.; Yoshida, Y.; Sasaki, T.; Takeno, S.; Hoshino, S. Impact of perioperative probiotic treatment for surgical site infections in patients with colorectal cancer. Exp. Ther. Med. 2015, 10, 966–972. [Google Scholar] [CrossRef] [Green Version]
- Moraffah, F.; Kiani, M.; Abdollahi, M.; Yoosefi, S.; Vatanara, A.; Samadi, N. In vitro-in vivo correlation for the antibacterial effect of Lactiplantibacillus plantarum as a topical healer for infected burn wound. Probiotics Antimicrob. Proteins 2022, 14, 675–689. [Google Scholar] [CrossRef]
- Bücher, C.; Burtscher, J.; Domig, K.J. Propionic acid bacteria in the food industry: An update on essential traits and detection methods. Compr. Rev. Food Sci. Food Saf. 2021, 20, 4299–4323. [Google Scholar] [CrossRef]
- Dubernet, S.; Desmasures, N.; Guéguen, M. A PCR-based method for identification of lactobacilli at the genus level. FEMS Microbiol. Lett. 2002, 214, 271–275. [Google Scholar] [CrossRef]
- Sul, S.Y.; Kim, H.J.; Kim, T.W.; Kim, H.Y. Rapid identification of Lactobacillus and Bifidobacterium in probiotic products using multiplex PCR. J. Microbiol. Biotechnol. 2007, 17, 490–495. [Google Scholar] [PubMed]
- Walter, J.; Tannock, G.W.; Tilsala-Timisjarvi, A.; Rodtong, S.; Loach, D.M.; Munro, K.; Alatossava, T. Detection and identification of gastrointestinal Lactobacillus species by using denaturing gradient gel electrophoresis and species-specific PCR primers. Appl. Environ. Microbiol. 2000, 66, 297–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, H.; Fujita, T.; Suzuki, Y.; Benno, Y. Monitoring and survival of Lactobacillus gasseri SBT2055 in the human intestinal tract. Microbiol. Immunol. 2006, 50, 867–870. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, C.; Palmeira-de-Oliveira, R.; Martinez-de-Oliveira, J.; Neves, J.D.; Pestana, P.G.; Rolo, J.; Donders, G.; Palmeira-de-Oliveira, A. Development and validation of a new one step multiplex-pcr assay for the detection of ten Lactobacillus species. Anaerobe 2019, 59, 192–200. [Google Scholar] [CrossRef]
- Bernhard, A.E.; Field, K.G. Identification of nonpoint sources of fecal pollution in coastal waters by using host-specific 16S ribosomal DNA genetic markers from fecal anaerobes. Appl. Environ. Microbiol. 2000, 66, 1587–1594. [Google Scholar] [CrossRef] [Green Version]
- Sheu, S.J.; Hwang, W.Z.; Chiang, Y.C.; Lin, W.H.; Chen, H.C.; Tsen, H.Y. Use of Tuf gene-based primers for the PCR detection of probiotic Bifidobacterium species and enumeration of bifidobacteria in fermented milk by cultural and quantitative real-time pcr methods. J. Food Sci. 2010, 75, M521–M527. [Google Scholar] [CrossRef]
- Deasy, B.M.; Rea, M.C.; Fitzgerald, G.F.; Cogan, T.M.; Beresford, T.P. A rapid PCR based method to distinguish between Lactococcus and Enterococcus. Syst. Appl. Microbiol. 2000, 23, 510–522. [Google Scholar] [CrossRef]
- Rossi, F.; Torriani, S.; Dellaglio, F. Genus- and species-specific PCR-based detection of dairy propionibacteria in environmental samples by using primers targeted to the genes encoding 16S rRNA. Appl. Environ. Microbiol. 1999, 65, 4241–4244. [Google Scholar] [CrossRef] [Green Version]
- Majeed, M.; Nagabhushanam, K.; Arumugam, S.; Alli, F. Novel PCR Primers and Methods Thereof for the Identification of Bacillus coagulans. WO 2017/058741, 6 April 2017. [Google Scholar]
- Wattiau, P.; Renard, M.E.; Ledent, P.; Debois, V.; Blackman, G.; Agathos, S.N. A PCR test to identify Bacillus subtilis and closely related species and its application to the monitoring of wastewater biotreatment. Appl. Microbiol. Biotechnol. 2001, 56, 816–819. [Google Scholar] [CrossRef]
- Cheng, S.; McCleskey, F.K.; Gress, M.J.; Petroziello, J.M.; Liu, R.; Namdari, H.; Beninga, K.; Salmen, A.; DelVecchio, V.G. A PCR assay for identification of Enterococcus faecium. J. Clin. Microbiol. 1997, 35, 1248–1250. [Google Scholar] [CrossRef] [Green Version]
- Mitterdorfer, G.; Mayer, H.K.; Kneifel, W.; Viernstein, H. Clustering of Saccharomyces boulardii strains within the species S. cerevisiae using molecular typing techniques. J. Appl. Microbiol. 2002, 93, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Fijan, S.; Šulc, D.; Steyer, A. Study of the in vitro antagonistic activity of various single-strain and multi-strain probiotics against Escherichia coli. Int. J. Environ. Res. Public Health 2018, 15, 1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igbafe, J.; Kilonzo-Nthenge, A.; Nahashon, S.N.; Mafiz, A.I.; Nzomo, M. Probiotics and antimicrobial effect of Lactiplantibacillus plantarum, Saccharomyces cerevisiae, and Bifidobacterium longum against common foodborne pathogens in poultry. Agriculture 2020, 10, 368. [Google Scholar] [CrossRef]
- Shokryazdan, P.; Sieo, C.C.; Kalavathy, R.; Liang, J.B.; Alitheen, N.B.; Faseleh Jahromi, M.; Ho, Y.W. Probiotic potential of lactobacillus strains with antimicrobial activity against some human pathogenic strains. Biomed Res. Int. 2014, 2014, 927268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, W.W.; Stout, T.R. Disc plate method of microbiological antibiotic assay. I. Factors influencing variability and error. Appl. Microbiol. 1971, 22, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Fijan, S. Antimicrobial effect of probiotics against common pathogens. In Probiotics and Prebiotics in Human Nutrition and Health; Rao, V., Rao, L., Eds.; InTech: Rijeka, Croatia, 2016; ISBN 978-953-51-2476-4. [Google Scholar] [CrossRef] [Green Version]
- Tranberg, A.; Klarin, B.; Johansson, J.; Påhlman, L.I. Efficacy of Lactiplantibacillus plantarum 299 and 299v against nosocomial oropharyngeal pathogens in vitro and as an oral prophylactic treatment in a randomized, controlled clinical trial. MicrobiologyOpen 2021, 10, e1151. [Google Scholar] [CrossRef] [PubMed]
- Holder, I.A.; Boyce, S.T. Agar well diffusion assay testing of bacterial susceptibility to various antimicrobials in concentrations non-toxic for human cells in culture. Burns 1994, 20, 426–429. [Google Scholar] [CrossRef]
- Varela-Trinidad, G.U.; Domínguez-Díaz, C.; Solórzano-Castanedo, K.; Íñiguez-Gutiérrez, L.; Hernández-Flores, T.D.J.; Fafutis-Morris, M. Probiotics: Protecting our health from the gut. Microorganisms 2022, 10, 1428. [Google Scholar] [CrossRef]
- Millette, M.; Luquet, F.M.; Lacroix, M. In vitro growth control of selected pathogens by Lactobacillus acidophilus- and Lactobacillus casei-fermented milk. Lett. Appl. Microbiol. 2007, 44, 314–319. [Google Scholar] [CrossRef]
- Saxelin, M.; Tynkkynen, S.; Mattila-Sandholm, T.; de Vos, W.M. Probiotic and other functional microbes: From markets to mechanisms. Curr. Opin. Biotechnol. 2005, 16, 204–211. [Google Scholar] [CrossRef]
- Wang, H.; Jiang, X. Isolation and characterization of competitive exclusion microorganisms from animal wastes-based composts against Listeria monocytogenes. J. Appl. Microbiol. 2022, 132, 4531–4543. [Google Scholar] [CrossRef] [PubMed]
- Dowarah, R.; Verma, A.K.; Agarwal, N.; Singh, P.; Singh, B.R. Selection and characterization of probiotic lactic acid bacteria and its impact on growth, nutrient digestibility, health and antioxidant status in weaned piglets. PLoS ONE 2018, 13, e0192978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Kholy, A.M.; El-Shinawy, S.H.; Meshref, A.M.S.; Korny, A.M. Screening of antagonistic activity of probiotic bacteria against some food-borne pathogens. J. Appl. Environ. Microbiol. 2014, 2, 53–60. [Google Scholar] [CrossRef]
- Varma, P.; Nisha, N.; Dinesh, K.R.; Kumar, A.V.; Biswas, R. Anti-infective properties of Lactobacillus fermentum against Staphylococcus aureus and Pseudomonas aeruginosa. J. Mol. Microbiol. Biotechnol. 2011, 20, 137–143. [Google Scholar] [CrossRef]
- Jeanson, S.; Floury, J.; Gagnaire, V.; Lortal, S.; Thierry, A. Bacterial colonies in solid media and foods: A review on their growth and interactions with the micro-environment. Front. Microbiol. 2015, 6, 1284. [Google Scholar] [CrossRef] [Green Version]
- Fijan, S. Influence of the growth of Pseudomonas aeruginosa in milk fermented by multispecies probiotics and kefir microbiota. J. Probiotics Health 2015, 4, 1–6. [Google Scholar] [CrossRef]
- Skandamis, P.N.; Jeanson, S. Colonial vs. Planktonic type of growth: Mathematical modeling of microbial dynamics on surfaces and in liquid, semi-liquid and solid foods. Front. Microbiol. 2015, 6, 1178. [Google Scholar] [CrossRef] [Green Version]
- Chapman, C.M.; Gibson, G.R.; Todd, S.; Rowland, I. Comparative in vitro inhibition of urinary tract pathogens by single- and multi-strain probiotics. Eur. J. Nutr. 2013, 52, 1669–1677. [Google Scholar] [CrossRef]
- Monteiro, C.; Do Carmo, M.; Melo, B.; Alves, M.; Dos Santos, C.; Monteiro, S.; Bomfim, M.; Fernandes, E.; Monteiro-Neto, V. In vitro antimicrobial activity and probiotic potential of Bifidobacterium and Lactobacillus against species of Clostridium. Nutrients 2019, 11, 448. [Google Scholar] [CrossRef] [Green Version]
- Tejero-Sariñena, S.; Barlow, J.; Costabile, A.; Gibson, G.R.; Rowland, I. In vitro evaluation of the antimicrobial activity of a range of probiotics against pathogens: Evidence for the effects of organic acids. Anaerobe 2012, 18, 530–538. [Google Scholar] [CrossRef]
- Favaro, L.; Barretto Penna, A.L.; Todorov, S.D. Bacteriocinogenic lab from cheeses—Application in biopreservation? Trends Food Sci. Technol. 2015, 41, 37–48. [Google Scholar] [CrossRef]
- Piwowarek, K.; Lipińska, E.; Hać-Szymańczuk, E.; Kieliszek, M.; Ścibisz, I. Propionibacterium spp.—Source of propionic acid, vitamin b12, and other metabolites important for the industry. Appl. Microbiol. Biotechnol. 2018, 102, 515–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Issaoui, K.; Khay, E.O.; Abrini, J.; Zinebi, S.; Amajoud, N.; Senhaji, N.S.; Abriouel, H. Molecular identification and antibiotic resistance of bacteriocinogenic lactic acid bacteria isolated from table olives. Arch. Microbiol. 2021, 203, 597–607. [Google Scholar] [CrossRef] [PubMed]
- Fredua-Agyeman, M.; Gaisford, S. Assessing inhibitory activity of probiotic culture supernatants against Pseudomonas aeruginosa: A comparative methodology between agar diffusion, broth culture and microcalorimetry. World J. Microbiol. Biotechnol. 2019, 35, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FDA. Content and Format of Premarket Notification [510(k)] Submissions for Liquid Chemical Sterilants/ High Level Disinfectants; U.S. Department of Health And Human Services, Food and Drug Administration: Washington, DC, USA, 2000.
- Matijašić, B.B.; Obermajer, T.; Rogelj, I. Quantification of Lactobacillus gasseri, Enterococcus faecium and Bifidobacterium infantis in a probiotic otc drug by real-time pcr. Food Control 2010, 21, 419–425. [Google Scholar] [CrossRef]
- Goldstein, E.J.C.; Tyrrell, K.L.; Citron, D.M. Lactobacillus species: Taxonomic complexity and controversial susceptibilities. Clin. Infect. Dis. 2015, 60, S98–S107. [Google Scholar] [CrossRef] [Green Version]
- Patro, J.N.; Ramachandran, P.; Lewis, J.L.; Mammel, M.K.; Barnaba, T.; Pfeiler, E.A.; Elkins, C.A. Development and utility of the fda ‘gutprobe’ DNA microarray for identification, genotyping and metagenomic analysis of commercially available probiotics. J. Appl. Microbiol. 2015, 118, 1478–1488. [Google Scholar] [CrossRef]
- Otaka, M.; Kikuchi-Hayakawa, H.; Ogura, J.; Ishikawa, H.; Yomogida, Y.; Ota, M.; Hidese, S.; Ishida, I.; Aida, M.; Matsuda, K.; et al. Effect of Lacticaseibacillus paracasei strain Shirota on improvement in depressive symptoms, and its association with abundance of actinobacteria in gut microbiota. Microorganisms 2021, 9, 1026. [Google Scholar] [CrossRef]
- Kim, E.; Yang, S.M.; Cho, E.J.; Kim, H.Y. Novel real-time PCR assay for Lactobacillus casei group species using comparative genomics. Food Microbiol. 2020, 90, 103485. [Google Scholar] [CrossRef]
- Szajewska, H.; Kołodziej, M. Systematic review with meta-analysis: Lactobacillus rhamnosus GG in the prevention of antibiotic-associated diarrhoea in children and adults. Aliment. Pharmacol. Ther. 2015, 42, 1149–1157. [Google Scholar] [CrossRef]
- Li, Y.-T.; Xu, H.; Ye, J.-Z.; Wu, W.-R.; Shi, D.; Fang, D.-Q.; Liu, Y.; Li, L.-J. Efficacy of Lactobacillus rhamnosus GG in treatment of acute pediatric diarrhea: A systematic review with meta-analysis. World J. Gastroenterol. 2019, 25, 4999–5016. [Google Scholar] [CrossRef] [PubMed]
- Szajewska, H.; Skórka, A.; Ruszczyński, M.; Gieruszczak-Białek, D. Meta-analysis: Lactobacillus GG for treating acute gastroenteritis in children—Updated analysis of randomised controlled trials. Aliment. Pharmacol. Ther. 2013, 38, 467–476. [Google Scholar] [CrossRef] [PubMed]
- Laursen, R.P.; Hojsak, I. Probiotics for respiratory tract infections in children attending day care centers—A systematic review. Eur. J. Pediatr. 2018, 177, 979–994. [Google Scholar] [CrossRef] [PubMed]
- Capurso, L. Thirty years of Lactobacillus rhamnosus GG: A review. J. Clin. Gastroenterol. 2019, 53 (Suppl. 1), S1–S41. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.J.; Lee, N.-K.; Paik, H.-D. Probiotic characterization of Lactobacillus brevis Ku15153 showing antimicrobial and antioxidant effect isolated from kimchi. Food Sci. Biotechnol. 2019, 28, 1521–1528. [Google Scholar] [CrossRef]
- Wang, Y.; Jia, D.; Wang, J.-H.; Li, H.-H.; Liu, J.-L.; Liu, A.-H.; Wang, J.-M.; Guan, G.-Q.; Luo, J.-X.; Yin, H.; et al. Assessment of probiotic adhesion and inhibitory effect on Escherichia coli and Salmonella adhesion. Arch. Microbiol. 2021, 203, 6267–6274. [Google Scholar] [CrossRef]
- Papadimitriou, K.; Zoumpopoulou, G.; Foligné, B.; Alexandraki, V.; Kazou, M.; Pot, B.; Tsakalidou, E. Discovering probiotic microorganisms: In vitro, in vivo, genetic and omics approaches. Front. Microbiol. 2015, 6, 58. [Google Scholar] [CrossRef] [Green Version]
- Vinderola, G.; Gueimonde, M.; Gomez-Gallego, C.; Delfederico, L.; Salminen, S. Correlation between in vitro and in vivo assays in selection of probiotics from traditional species of bacteria. Trends Food Sci. Technol. 2017, 68, 83–90. [Google Scholar] [CrossRef]
- Li, X.; Liu, Y.; Guo, X.; Ma, Y.; Zhang, H.; Liang, H. Effect of Lactobacillus casei on lipid metabolism and intestinal microflora in patients with alcoholic liver injury. Eur. J. Clin. Nutr. 2021, 75, 1227–1236. [Google Scholar] [CrossRef]
- Mai, T.T.; Thi Thu, P.; Thi Hang, H.; Trang, T.T.T.; Yui, S.; Shigehisa, A.; Tien, V.T.; Dung, T.V.; Nga, P.B.; Hung, N.T.; et al. Efficacy of probiotics on digestive disorders and acute respiratory infections: A controlled clinical trial in young vietnamese children. Eur. J. Clin. Nutr. 2021, 75, 513–520. [Google Scholar] [CrossRef]
- Macnaughtan, J.; Figorilli, F.; García-López, E.; Lu, H.; Jones, H.; Sawhney, R.; Suzuki, K.; Fairclough, S.; Marsden, J.; Moratella, A.; et al. A double-blind, randomized placebo-controlled trial of probiotic Lactobacillus casei Shirota in stable cirrhotic patients. Nutrients 2020, 12, 1651. [Google Scholar] [CrossRef] [PubMed]
- Paniágua, A.L.; Correia, A.F.; Pereira, L.C.; de Alencar, B.M.; Silva, F.B.A.; Almeida, R.M.; de Medeiros Nóbrega, Y.K. Inhibitory effects of Lactobacillus casei Shirota against both Candida auris and Candida spp. Isolates that cause vulvovaginal candidiasis and are resistant to antifungals. BMC Complement. Med. Ther. 2021, 21, 237. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liang, H. Effects of Lactobacillus casei on iron metabolism and intestinal microflora in rats exposed to alcohol and iron. Turk. J. Gastroenterol. 2022, 33, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Szajewska, H.; Urbańska, M.; Chmielewska, A.; Weizman, Z.; Shamir, R. Meta-analysis: Lactobacillus reuteri strain DSM 17938 (and the original strain ATCC 55730) for treating acute gastroenteritis in children. Benef. Microbes 2014, 5, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Liévin-Le Moal, V.; Servin, A.L. Anti-infective activities of lactobacillus strains in the human intestinal microbiota: From probiotics to gastrointestinal anti-infectious biotherapeutic agents. Clin. Microbiol. Rev. 2014, 27, 167–199. [Google Scholar] [CrossRef] [Green Version]
- Moreno Márquez, C.; Fernández Álvarez, P.; Valdés Delgado, T.; Castro Laria, L.; Argüelles Arias, F.; Caunedo Álvarez, A.; Gómez Rodríguez, B.J. Randomized, double-blind, placebo-controlled clinical trial on the usefulness of probiotic Lactobacillus reuteri in bismuth-containing quadruple eradication therapy for infection with Helicobacter pylori. Rev. Española Enferm. Dig. 2021. [Google Scholar] [CrossRef]
- Invernici, M.M.; Furlaneto, F.A.C.; Salvador, S.L.; Ouwehand, A.C.; Salminen, S.; Mantziari, A.; Vinderola, G.; Ervolino, E.; Santana, S.I.; Silva, P.H.F.; et al. Bifidobacterium animalis subsp lactis HN019 presents antimicrobial potential against periodontopathogens and modulates the immunological response of oral mucosa in periodontitis patients. PLoS ONE 2020, 15, e0238425. [Google Scholar] [CrossRef]
- Arunachalam, K.; Gill, H.; Chandra, R. Enhancement of natural immune function by dietary consumption of Bifidobacterium lactis (HN019). Eur. J. Clin. Nutr. 2000, 54, 263–267. [Google Scholar] [CrossRef] [Green Version]
- Fredua-Agyeman, M.; Stapleton, P.; Basit, A.W.; Beezer, A.E.; Gaisford, S. In vitro inhibition of Clostridium difficile by commercial probiotics: A microcalorimetric study. Int. J. Pharm. 2017, 517, 96–103. [Google Scholar] [CrossRef]
- Taipale, T.; Pienihäkkinen, K.; Isolauri, E.; Larsen, C.; Brockmann, E.; Alanen, P.; Jokela, J.; Söderling, E. Bifidobacterium animalis subsp. lactis BB-12 in reducing the risk of infections in infancy. Br. J. Nutr. 2011, 105, 409–416. [Google Scholar] [CrossRef]
- Dyshlyuk, L.S.; Milentyeva, I.S.; Asyakina, L.K.; Ostroumov, L.A.; Osintsev, A.M.; Pozdnyakova, A.V. Using Bifidobacterium and Propionibacterium strains in probiotic consortia to normalize the gastrointestinal tract. Braz. J. Biol. 2022, 84, e256945. [Google Scholar] [CrossRef] [PubMed]
- Koskinen, P.; Deptula, P.; Smolander, O.P.; Tamene, F.; Kammonen, J.; Savijoki, K.; Paulin, L.; Piironen, V.; Auvinen, P.; Varmanen, P. Complete genome sequence of Propionibacterium freudenreichii DSM 20271(T). Stand. Genom. Sci. 2015, 10, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyra, A.; Krogius-Kurikka, L.; Nikkilä, J.; Malinen, E.; Kajander, K.; Kurikka, K.; Korpela, R.; Palva, A. Effect of a multispecies probiotic supplement on quantity of irritable bowel syndrome-related intestinal microbial phylotypes. BMC Gastroenterol. 2010, 10, 110. [Google Scholar] [CrossRef] [Green Version]
- Myllyluoma, E.; Ahlroos, T.; Veijola, L.; Rautelin, H.; Tynkkynen, S.; Korpela, R. Effects of anti-Helicobacter pylori treatment and probiotic supplementation on intestinal microbiota. Int. J. Antimicrob. Agents 2007, 29, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.-K.; Kim, W.-S.; Paik, H.-D. Bacillus strains as human probiotics: Characterization, safety, microbiome, and probiotic carrier. Food Sci. Biotechnol. 2019, 28, 1297–1305. [Google Scholar] [CrossRef] [PubMed]
- Ahire, J.J.; Neelamraju, J.; Madempudi, R.S. Behavior of Bacillus coagulans unique IS2 spores during passage through the simulator of human intestinal microbial ecosystem (shime) model. LWT 2020, 124, 109196. [Google Scholar] [CrossRef]
- Madempudi, R.S.; Neelamraju, J.; Ahire, J.J.; Gupta, S.K.; Shukla, V.K. Bacillus coagulans unique IS2 in constipation: A double-blind, placebo-controlled study. Probiotics Antimicrob. Proteins 2020, 12, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Ratna Sudha, M.; Yelikar, K.A.; Deshpande, S. Clinical study of Bacillus coagulans unique IS-2 (ATCC PTA-11748) in the treatment of patients with bacterial vaginosis. Indian J. Microbiol. 2012, 52, 396–399. [Google Scholar] [CrossRef] [Green Version]
- Abid, R.; Waseem, H.; Ali, J.; Ghazanfar, S.; Muhammad Ali, G.; Elasbali, A.M.; Alharethi, S.H. Probiotic yeast Saccharomyces: Back to nature to improve human health. J. Fungi 2022, 8, 444. [Google Scholar] [CrossRef]
- Dinleyici, E.C.; Kara, A.; Ozen, M.; Vandenplas, Y. Saccharomyces boulardii CNCM I-745 in different clinical conditions. Expert Opin. Biol. Ther. 2014, 14, 1593–1609. [Google Scholar] [CrossRef]
- Szajewska, H.; Guarino, A.; Hojsak, I.; Indrio, F.; Kolacek, S.; Shamir, R.; Vandenplas, Y.; Weizman, Z. Use of probiotics for management of acute gastroenteritis: A position paper by the espghan working group for probiotics and prebiotics. J. Pediatr. Gastroenterol. Nutr. 2014, 58, 531–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guarino, A.; Ashkenazi, S.; Gendrel, D.; Lo Vecchio, A.; Shamir, R.; Szajewska, H. European society for pediatric gastroenterology, hepatology, and nutrition/european society for pediatric infectious diseases evidence-based guidelines for the management of acute gastroenteritis in children in Europe: Update 2014. J. Pediatr. Gastroenterol. Nutr. 2014, 59, 132–152. [Google Scholar] [CrossRef] [PubMed]
- Ishaque, S.M.; Khosruzzaman, S.M.; Ahmed, D.S.; Sah, M.P. A randomized placebo-controlled clinical trial of a multi-strain probiotic formulation (Bio-Kult®) in the management of diarrhea-predominant irritable bowel syndrome. BMC Gastroenterol. 2018, 18, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reininghaus, E.Z.; Wetzlmair, L.C.; Fellendorf, F.T.; Platzer, M.; Queissner, R.; Birner, A.; Pilz, R.; Hamm, C.; Maget, A.; Rieger, A.; et al. Probiotic treatment in individuals with euthymic bipolar disorder: A pilot-study on clinical changes and compliance. Neuropsychobiology 2020, 79, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Moser, A.M.; Spindelboeck, W.; Halwachs, B.; Strohmaier, H.; Kump, P.; Gorkiewicz, G.; Högenauer, C. Effects of an oral synbiotic on the gastrointestinal immune system and microbiota in patients with diarrhea-predominant irritable bowel syndrome. Eur. J. Nutr. 2019, 58, 2767–2778. [Google Scholar] [CrossRef] [Green Version]
- Kienesberger, B.; Obermüller, B.; Singer, G.; Arneitz, C.; Gasparella, P.; Klymiuk, I.; Horvath, A.; Stadlbauer, V.; Magnes, C.; Zügner, E.; et al. Insights into the composition of a co-culture of 10 probiotic strains (Omni Biotic® AAD10) and effects of its postbiotic culture supernatant. Nutrients 2022, 14, 1194. [Google Scholar] [CrossRef]
- Sprunt, K.; Leidy, G. The use of bacterial interference to prevent infection. Can. J. Microbiol. 1988, 34, 332–338. [Google Scholar] [CrossRef] [Green Version]
- Florey, H.W. Use of micro-organisms for therapeutic purposes. Br. Med. J. 1945, 2, 635–642. [Google Scholar] [CrossRef] [Green Version]
- Howard, J.C.; Reid, G.; Gan, B.S. Probiotics in surgical wound infections: Current status. Clin. Investig. Med. 2004, 27, 274–281. [Google Scholar]
- Bekiaridou, A.; Karlafti, E.; Oikonomou, I.M.; Ioannidis, A.; Papavramidis, T.S. Probiotics and their effect on surgical wound healing: A systematic review and new insights into the role of nanotechnology. Nutrients 2021, 13, 4265. [Google Scholar] [CrossRef]
- Reid, G.; Jass, J.; Sebulsky, M.T.; McCormick, J.K. Potential uses of probiotics in clinical practice. Clin. Microbiol. Rev. 2003, 16, 658–672. [Google Scholar] [CrossRef] [PubMed]
- Siddharthan, R.; Chapek, M.; Warren, M.; Martindale, R. Probiotics in prevention of surgical site infections. Surg. Infect. (Larchmt) 2018, 19, 781–784. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Label | Supplement | Lactobacilli 1 | Bifidobacteria | Other Bacteria or Fungi |
|---|---|---|---|---|
| MS1 | OMNi-BiOTiC® HETOX, Institut Allergosan, Austria | Lacticaseibacillus casei W56 Lactobacillus acidophilus W37 Levilactobacillus brevis W63 Lactobacillus salivarius W24 | Bifidobacterium lactis W52 Bifidobacterium bifidum W23 | Lactococcus lactis W58 Lactococcus lactis W19 |
| MS 2 | OMNi-BiOTiC® STRESS Repair, Institut Allergosan, Austria | Lacticaseibacillus casei W56 Lactobacillus acidophilus W22 Lacticaseibacillus paracasei W20 Lactiplantibacillus plantarum W62 Ligilactobacillus salivarius W24 | Bifidobacterium lactis W52 Bifidobacterium lactis W51 Bifidobacterium bifidum W23 | Lactococcus lactis W19 |
| MS 3 | OMNi-BiOTiC® 6, Institut Allergosan, Austria | Lactobacillus acidophilus W55 Ligilactobacillus salivarius W57 Lacticaseibacillus casei W56 | Bifidobacterium animalis W53 | Enterococcus faecium W54 Lactococcus lactis W58 |
| MS 4 | OMNi-BiOTiC® FLORA plus+, Institut Allergosan, Austria | Lactobacillus crispatus LBV88 Lacticaseibacillus rhamnosus LBV96 Lactobacillus gasseri LBV150N Lactobacillus jensenii LBV116 | / | / |
| MS 5 | OMNi-BiOTiC® Activ, Institut Allergosan, Austria | Lacticaseibacillus casei W56 Lactobacillus acidophilus W37, Levilactobacillus brevis W63, Ligilactobacillus salivarius W24 | Bifidobacterium lactis W52, Bifidobacterium longum W108, Bifidobacterium breve W25, Bifidobacterium lactis W51, Bifidobacterium bifidum W23 | Lactococcus lactis W58, Lactococcus lactis W19, |
| MS 6 | NutriVital Ultra SB, NutriVital Ply Ltd., Australia | Lactobacillus acidophilus La14 Lactiplantibacillus plantarum Lp-115 | Bifidobacterium animalis subsp. lactis BI-04 | Saccharomyces cerevisiae var. boulardii |
| MS 7 | (Bio-Kult®), Protexin Lopsen Head, UK, | Lacticaseibacillus casei PXN 37, Lactiplantibacillus plantarum PXN 47, Lacticaseibacillus rhamnosus PXN 54, Lactobacillus acidophilus PXN 35, Lactobacillus delbrueckii subsp. bulgaricus PXN 39, Lactobacillus helveticus PXN 45, Ligilactobacillus salivarius PXN 57 | Bifidobacterium bifidum PXN 23, Bifidobacterium breve PXN 25, Bifidobacterium longum PXN 30, Bifidobacterium infantis PXN 27 | Bacillus subtilis PXN 21, Lactococcus lactis subsp. lactis PXN 63, Streptococcus thermophilus PXN 66 |
| Label | Supplement | Strains |
|---|---|---|
| SS01 | Waya® LGG® forte, Medis GmbH, Austria | Lacticaseibacillus rhamnosus LGG |
| SS02 | Yakult®, Yakult Honsha Co, Ltd., Yakult Europe, Italy | Lacticaseibacillus paracasei Shirota |
| SS03 | BioGaia®, TwoPac AB, Sweden | Limosilactobacillus reuteri DSM 17938 |
| SS04 | German Collection of Microorganisms and Cell Cultures GmbH | Lactiplantibacillus plantarum subsp. plantarum DSM 2601 |
| SS05 | Probactiol® senior, Metagenics Italia S.r.l., Italia | Bifidobacterium animalis subsp. lactis HN019 |
| SS06 | Baby Linbi®, Lek Pharmaceutical company d.d., Slovenia | Bifidobacterium animalis subsp. lactis BB-12 |
| SS07 | ProLife® sporogenes, Zeta Farmaceutici, S.p.a., Italy | Bacillus coagulans MTCC 5260 |
| SS08 | German Collection of Microorganisms and Cell Cultures GmbH | Propionibacterium freudenreichii subsp. freudenreichii DSM 20271 |
| SS09 | German Collection of Microorganisms and Cell Cultures GmbH | Acidipropionibacterium acidipropionici DSM 20272 |
| SS10 | Optim PropioniBacter, Laboratoire Optim, Bionoto sprl, Belgium | Propionibacterium freudenreichii subsp. shermanii |
| SS11 | SB Probiotic, Blooms, Phytologic Pty Ltd., Australia | Saccharomyces cerevisiae var. boulardii |
| Label | Pathogen | Origin |
|---|---|---|
| 1 | Staphylococcus aureus | Patient with leg ulcer infection |
| 2 | Patient with diabetic ulcer infection | |
| 3 | Pseudomonas aeruginosa | Patient with inguinal infection after cardio intervention |
| 4 | Patient with gastrostomy site infection | |
| 5 | Enterococcus faecalis | Patient with infection at central venous catheterization |
| 6 | Patient with sternal wound infection | |
| 7 | Escherichia coli | Patient with surgical wound infection and dehiscence |
| 8 | Patient with surgical wound infection | |
| 9 | Klebsiella pneumoniae | Patient with sternal wound infection |
| 10 | Patient with surgical wound infection | |
| 11 | Enterobacter spp. | Patient with leg ulcer infection |
| 12 | Patient with inguinal infection after cardio intervention | |
| 13 | Acinetobacter spp. | Patient with bedsore (pressure ulcer) infection |
| 14 | Bacteroides spp. | Patient with perianal infection |
| 15 | Patient with bedsore (pressure ulcer) infection |
| Microorganism | Primer Pairs (5′–3′) | Product Size | Reference | |
|---|---|---|---|---|
| Lactobacilli spp. | ||||
| Lactobacilli spp. | LbLMA1-rev | CTC AAA ACT AAA CAA AGT TTC | 220 bp | Dubernet et al., 2002 [42] |
| R-16-1 | CTT GTA CAC ACC GCC CGT C | |||
| Lacticaseibacillus rhamnosus | Rham 1 | GTC GAA CGA GTT CTG ATT ATT G | 158 bp | Sul et al., 2007 [43] |
| RhamR | GAA CCA TGC GGT TCT TGG AT | |||
| Lactobacillus acidophilus | LacidoF | CAC TTC GGT GAT GAC GTT GG | 575 bp | |
| LacidoR | CGA TGC AGT TCC TCG GTT AAG C | |||
| Lacticaseibacillus casei | PrI | CAG ACT GAA AGT CTG ACG G | 200 bp | Walter et al., 2000 [44] |
| CasII | GCG ATG CGA ATT TCT TTT TC | |||
| Limosilactobacillus reuteri | Lfpr | GCC GCC TAA GGT GGG ACA GAT | 350 bp | |
| Reu | AAC ACT CAA GGA TTG TCT GA | |||
| Lactobacillus gasseri | Lgas-3 | AGC GAC CGA GAG AGA GAG A | 360 bp | Takahashi et al., 2006 [45] |
| Lgas-2 | TGC TAT CGC TTC AAG TGC TT | |||
| Lactiplantibacillus plantarum | LplanF | CGA GAC AGC AAT TCC TGC ACT CG | 176 bp | Gaspar et al., 2019 [46] |
| LplanR | CCT CAG AAA CAG TCC GGT TGA C | |||
| Bifidobacteria spp. | ||||
| Bifidobacterium spp. | Bif164F | GGG TGG TAA TGC CGG ATG | 453 bp | Bernhard et al., 2000 [47] |
| Bif601R | TAA GCC ATG GAC TTT CAC ACC | |||
| Bifidobacterium bifidum | BifF | ATT TGA GCC ACT GTC TGG TG | 431 bp | Sul et al., 2007 [43] |
| BifR | CAT CCG GGA ACG TCG GGA AA | |||
| Bifidobacterium longum | BiflongF | TTC CAG TTG ATC GCA TGG TC | 831 bp | |
| BiflongR | GGG AAG CCG TAT CTC TAC GA | |||
| Bifidobacterium animalis | Bani-tF | TCA CGA CAA GTG GGT TGC CA | 178 bp | Sheu et al., 2010 [48] |
| Bani-tR | GTT GAT CGG CAG CTT GCC G | |||
| Other bacteria and fungi | ||||
| Lactococcus spp. | L1 | AAC TCT GTT GTT AGA G | 570 bp | Deasy et al., 2000 [49] |
| L2 | ATC TCT AGG AAT AGC AC | |||
| Propionibacterium spp. | PB1 | AGT GGC GAA GGC GGT TCT CTG GA | 865 bp | Rossi et al., 1999 [50] |
| PB2 | TGG GGT CGA GTT GCA GAC CCC AAT | |||
| Bacillus coagulans | BC1-F | ACA GGG CTT TCA GAT ACC CG | 990 bp | Majeed et al., 2017 2017 [51] |
| BC1-R | CGG GGA TCC GTC CAT CAA AA | |||
| Bacillus subtilis | Bsub5F | AAG TCG AGC GGA CAG ATG G | 595 bp | Wattiau et al., 2001 [52] |
| Bsub5R | CCA GTT TCC AAT GAC CCT CCC C | |||
| Enterococcus faecium | EM1F | TTG AGG CAG ACC AGA TTG ACG | 658 bp | Cheng et al., 1997 [53] |
| EM1R | TAT GAC AGC GAC TCC GAT TCC | |||
| Saccharomyces cerevisiae | SC1 | AAC GGT GAG AGA TTT CTG TGC | 1170 bp | Mitterdorfer et al., 2002 [54] |
| SC2 | AGC TGG CAG TAT TCC CAC AG | |||
| PCR Program | Denaturation 1 | Annealing | Extension | No. of Cycles | Reference/Modified Program |
|---|---|---|---|---|---|
| Lactobacilli spp. | 30 s at 94 °C | 30 s at 55 °C | 30 s at 72 °C | 20 | Dubernet, et al., 2002 [42] |
| Lacticaseibacillus casei, Limosilalactobacillus reuteri | 30 | Walter et al., 2000 [44] | |||
| Lacticaseibacillus rhamnosus, Lactobacillus acidophilus, Bifidobacterium longum | 30 s at 94 °C | 30 s at 63 °C | 30 s at 72 °C | 30 | Sul, et al., 2007 [43] |
| Lactobacillus gasseri | 30 s at 94 °C | 120 s at 65 °C | 120 s at 72 °C | 35 | Takahashi et al., 2006 [45] |
| Lactiplantibacillus plantarum | 15 s at 94 °C | 30 s at 60 °C | 60 s at 72 °C | 40 | Gaspar et al., 2019 [46] |
| Bifidobacterium spp. | 30 s at 94 °C | 60 s at 53 °C | 120 s at 72 °C | 35 | Bernhard et al., 2000 [47] |
| Bifidobacterium bifidum | 30 s at 94 °C | 45 s at 57 °C | 30 s at 72 °C | 35 | Modified in this study |
| Bifidobacterium animalis | 35 s at 94 °C | 35 s at 60 °C | 40 s at 72 °C | 35 | Sheu et al., 2010 [48] |
| Propionibacterium spp. | 30 s at 94 °C | 15 s at 70 °C | 60 s at 72 °C | 40 | Rossi et al., 1999 [50] |
| Bacillus coagulans | 30 s at 94 °C | 30 s at 60 °C | 60 s at 72 °C | 30 | Majeed et al., 2017 [51] |
| Bacillus subtilis | 30 s at 94 °C | 120 s at 65 °C | 120 s at 72 °C | 30 | Wattiau et al., 2001 [52] |
| Enterococcus faecium | 60 s at 94 °C | 60 s at 54 °C | 60 s at 72 °C | 40 | Fijan et al., 2018 [55] |
| Lactococcus spp. | 60 s at 94 °C | 60 s at 50 °C | 60 s at 72 °C | 30 | Modified in this study |
| Saccharomyces cerevisiae | 60 s at 94 °C | 60 s at 50 °C | 60 s at 72 °C | 30 | Mitterdorfer et al., 2000 [54] |
| Sample | Confirmed Lactobacilli | Confirmed Bifidobacteria | Confirmed Other Bacteria or Fungi | |||
|---|---|---|---|---|---|---|
| Genus-Specific PCR | Species-Specific PCR | Genus-Specific PCR | Species-Specific PCR | Genus-Specific PCR | Species-Specific PCR | |
| MS1 | Lactobacilli 1 spp. | L. acidophilus, L. casei | Bifidobacterium | B. animalis, B. bifidum | Lactococcus | / |
| MS2 | Lactobacilli spp. | L. acidophilus, L. casei, L. plantarum | Bifidobacterium | B. animalis, B. bifidum | Lactococcus | / |
| MS3 | Lactobacilli spp. | L. acidophilus, L. casei | Bifidobacterium | B. animalis | Lactococcus | E. faecium |
| MS4 | Lactobacilli spp. | L. gasseri, L. rhamnosus | / | / | / | / |
| MS5 | Lactobacilli spp. | L. acidophilus, L. casei | Bifidobacterium | B. animalis, B. bifidum, B. longum | Lactococcus | / |
| MS6 | Lactobacilli spp. | L. acidophilus, L. plantarum | Bifidobacterium | B. animalis | / | Saccharomyces cerevisiae |
| MS7 | Lactobacilli spp. | L. acidophilus, L. casei, L. plantarum, L. rhamnosus | Bifidobacterium | B. animalis, B. bifidum, B. longum | Lactococcus | B. subtilis |
| Confirmed Genus-Specific PCR | Confirmed Species-Specific PCR | |
|---|---|---|
| SS01 | Lactobacilli spp.1 | Lacticaseibacillus rhamnosus |
| SS02 | Lactobacilli spp. | Lacticaseibacillus casei |
| SS03 | Lactobacilli spp. | Limosilactobacillus reuteri |
| SS04 | Lactobacilli spp. | Lactiplantibacillus plantarum |
| SS05 | Bifidobacterium genus | Bifidobacterium animalis |
| SS06 | Bifidobacterium spp. | Bifidobacterium animalis |
| SS07 | (Not conducted) | Bacillus coagulans |
| SS08 | Propionibacterium spp. | (Not conducted) |
| SS09 | Propionibacterium spp. | (Not conducted) |
| SS10 | Propionibacterium spp. | (Not conducted) |
| SS11 | (Not conducted) | Saccharomyces boulardii |
| Evaluation of Zone of Inhibition Using the Agar Spot Assay * | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sa1 | Sa2 | Pa3 | Pa4 | Ef5 | Ef6 | Ec7 | Ec8 | Kp9 | Kp10 | Ent11 | Ent12 | Ac13 | Bac14 | Bac15 | |
| MS1 | 3+ | 3+ | 3+ | 3+ | 3+ | 2+ | 3+ | 2+ | 2+ | 2+ | 2+ | 2+ | 3+ | 3+ | 3+ |
| MS2 | 3+ | 3+ | 3+ | 3+ | 3+ | 2+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ |
| MS3 | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 2+ | 3+ | 3+ | 3+ |
| MS4 | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 2+ | 2+ | 2+ | 3+ | 3+ |
| MS5 | 3+ | 2+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 2+ | 3+ | 3+ | 3+ | 3+ |
| MS6 | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 2+ | 3+ | 3+ | 3+ | 3+ |
| MS7 | 3+ | 3+ | 2+ | 3+ | 3+ | 3+ | 2+ | 3+ | 3+ | 3+ | 2+ | 3+ | 3+ | 3+ | 3+ |
| SS01 | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 2+ | 3+ | 3+ | 3+ |
| SS02 | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 2+ | 2+ | 2+ | 3+ | 3+ | 3+ |
| SS03 | 2+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 2+ | 2+ | 3+ | 3+ | 3+ |
| SS04 | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 2+ | 3+ | 3+ | 3+ | 3+ |
| SS05 | 2+ | 2+ | 2+ | 2+ | 2+ | 2+ | 2+ | 3+ | 3+ | 2+ | 3+ | 3+ | 2+ | 2+ | 3+ |
| SS06 | 2+ | 2+ | 2+ | 2+ | 2+ | 2+ | 2+ | 3+ | 3+ | 3+ | 3+ | 3+ | 2+ | 3+ | 2+ |
| SS07 | + | 2+ | 2+ | 2+ | + | + | + | + | + | + | + | + | + | + | + |
| SS08 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| SS09 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| SS10 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| SS11 | 2+ | 2+ | 3+ | 3+ | 2+ | 3+ | 3+ | 2+ | 2+ | 2+ | + | + | 2+ | 2+ | 2+ |
| Evaluation of Log Step Reduction Using the Co-Culturing Assay * | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sa1 | Sa2 | Pa3 | Pa4 | Ef5 | Ef6 | Ec7 | Ec8 | Kp9 | Kp10 | Ent11 | Ent12 | Ac13 | Bac14 | Bac15 | |
| MS1 | 2+ | 2+ | 3+ | 3+ | 2+ | 2+ | 3+ | 2+ | + | 2+ | 3+ | 3+ | 3+ | 3+ | 3+ |
| MS2 | 2+ | 2+ | 3+ | 3+ | 2+ | 2+ | 2+ | 2+ | + | 2+ | 3+ | 3+ | 3+ | 3+ | 2+ |
| MS3 | 2+ | 2+ | 2+ | 3+ | 2+ | 2+ | 3+ | 3+ | + | 3+ | 3+ | 3+ | 3+ | 3+ | 2+ |
| MS4 | 2+ | 2+ | 3+ | 3+ | 2+ | 2+ | 3+ | 3+ | 3+ | 3+ | 2+ | 3+ | 3+ | 3+ | 3+ |
| MS5 | 2+ | 2+ | 3+ | 3+ | + | 2+ | 3+ | 3+ | 3+ | 2+ | 3+ | 3+ | 3+ | 2+ | 3+ |
| MS6 | 2+ | 2+ | 3+ | 2+ | + | 2+ | 2+ | + | + | 2+ | 2+ | 3+ | 3+ | 2+ | + |
| MS7 | 2+ | 2+ | 3+ | 3+ | 2+ | 2+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 2+ | 2+ |
| SS01 | 2+ | + | 3+ | 3+ | + | + | 2+ | 2+ | 2+ | + | 2+ | 3+ | + | 2+ | + |
| SS02 | 2+ | 2+ | 3+ | 3+ | 2+ | + | 2+ | 2+ | 2+ | + | 3+ | 3+ | 2+ | 2+ | + |
| SS03 | 2+ | 2+ | 3+ | 3+ | 2+ | 2+ | 2+ | 3+ | + | 2+ | 3+ | 3+ | 2+ | + | + |
| SS04 | 3+ | 3+ | 3+ | 2+ | 2+ | 2+ | 3+ | 3+ | 3+ | 2+ | 3+ | 3+ | 2+ | 3+ | 2+ |
| SS05 | 2+ | 2+ | 3+ | 2+ | 2+ | 2+ | 3+ | 3+ | + | 3+ | 3+ | 3+ | 2+ | 3+ | 2+ |
| SS06 | 3+ | 3+ | 3+ | 3+ | + | 2+ | 2+ | 3+ | 2+ | 3+ | 3+ | 3+ | 2+ | + | + |
| SS07 | 2+ | + | 3+ | 3+ | 2+ | + | 2+ | 2+ | 2+ | 2+ | 2+ | 2+ | 2+ | + | + |
| SS08 | + | + | + | + | + | + | + | + | + | + | + | + | 2+ | 2+ | + |
| SS09 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| SS10 | 2+ | + | 2+ | + | + | + | + | 2+ | 2+ | 2+ | 2+ | 2+ | 2+ | + | 3+ |
| SS11 | 2+ | 2+ | + | + | + | + | 2+ | + | + | + | + | 2+ | + | + | + |
| Evaluation of Zone of Inhibition Using the Agar Well Diffusion Assay * | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sa1 | Sa2 | Pa3 | Pa4 | Ef5 | Ef6 | Ec7 | Ec8 | Kp9 | Kp10 | Ent11 | Ent12 | Ac13 | Bac14 | Bac15 | |
| MS1 | 2+ | 2+ | 2+ | 2+ | 2+ | 2+ | 2+ | 2+ | 2+ | 2+ | 2+ | + | 2+ | 2+ | + |
| MS2 | 2+ | 2+ | + | + | + | + | 2+ | 2+ | 2+ | 2+ | 2+ | + | + | – | + |
| MS3 | 2+ | + | + | 2+ | 2+ | + | 2+ | 2+ | + | 2+ | 2+ | + | + | 2+ | + |
| MS4 | 2+ | + | + | 2+ | 2+ | – | 2+ | 2+ | + | 2+ | 2+ | 2+ | + | + | 2+ |
| MS5 | 2+ | + | – | 2+ | + | – | 2+ | 2+ | + | 2+ | + | 2+ | + | + | 2+ |
| MS6 | + | + | 2+ | – | + | + | + | 3+ | + | + | 2+ | 2+ | + | + | 2+ |
| MS7 | 2+ | 2+ | 2+ | – | 2+ | 2+ | + | 2+ | + | + | + | + | + | + | |
| SS01 | 2+ | 2+ | 2+ | + | – | – | 2+ | 3+ | + | + | + | – | + | – | + |
| SS02 | 2+ | + | + | 2+ | + | + | 2+ | 2+ | + | 2+ | + | 2+ | + | – | – |
| SS03 | – | 2+ | + | + | – | – | 2+ | 2+ | + | – | 2+ | 2+ | + | – | + |
| SS04 | 2+ | + | – | + | + | – | + | 2+ | + | + | 2+ | + | + | – | – |
| SS05 | 2+ | 2+ | 2+ | + | – | – | 2+ | 2+ | 2+ | 2+ | + | + | – | – | |
| SS06 | + | – | + | + | + | 2+ | 2+ | 2+ | 2+ | 2+ | 2+ | 2+ | – | – | |
| SS07 | – | + | 2+ | 2+ | – | – | – | – | – | – | + | – | + | 2+ | – |
| SS08 | + | + | – | – | – | – | + | – | – | – | 2+ | – | – | – | – |
| SS09 | + | 2+ | – | – | – | – | + | – | – | – | – | – | – | – | – |
| SS10 | + | + | 2+ | + | – | 2+ | + | + | + | + | – | + | – | + | – |
| SS11 | + | – | 2+ | 2+ | + | – | – | 2+ | – | – | + | – | + | + | – |
| Evaluation of Zone of Inhibition Using the Agar Well Diffusion Assay * | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sa1 | Sa2 | Pa3 | Pa4 | Ef5 | Ef6 | Ec7 | Ec8 | Kp9 | Kp10 | Ent11 | Ent12 | Ac13 | Bac14 | Bac15 | |
| MS1 | – | + | 2+ | – | – | – | – | 2+ | – | – | – | – | – | – | – |
| MS2 | – | + | + | + | + | – | 2+ | 2+ | – | 2+ | – | – | + | 2+ | + |
| MS3 | – | + | + | + | 2+ | + | 2+ | + | 2+ | 2+ | – | – | + | – | + |
| MS4 | – | + | – | – | + | + | – | 2+ | – | 2+ | 2+ | 2+ | – | – | 2+ |
| MS5 | – | – | – | – | – | – | – | – | – | 2+ | – | + | – | – | 2+ |
| MS6 | + | – | – | – | – | – | – | – | – | – | – | – | + | – | 2+ |
| MS7 | + | 2+ | – | – | 2+ | 2+ | 2+ | – | – | – | + | 2+ | – | – | – |
| SS01 | – | 2+ | + | 2+ | – | – | 2+ | 2+ | – | – | 2+ | – | – | – | – |
| SS02 | – | 2+ | – | – | – | – | 2+ | 2+ | – | – | 2+ | – | – | 3+ | – |
| SS03 | – | 2+ | – | 2+ | – | – | 2+ | 2+ | – | – | 2+ | 2+ | – | – | – |
| SS04 | + | 2+ | – | – | – | – | + | 2+ | – | 2+ | 2+ | – | + | – | – |
| SS05 | + | + | – | – | – | – | – | 2+ | – | – | 2+ | – | + | 2+ | – |
| SS06 | + | – | – | – | 2+ | – | 2+ | + | 2+ | 2+ | + | + | + | – | 2+ |
| SS07 | + | + | – | – | – | – | – | – | – | – | – | – | + | – | – |
| SS08 | 2+ | + | – | – | – | – | – | – | – | – | 2+ | – | – | – | – |
| SS09 | + | 2+ | – | – | – | – | + | + | – | – | 2+ | – | – | – | – |
| SS10 | – | + | – | – | – | – | + | + | 2+ | – | – | – | – | – | – |
| SS11 | + | – | 2+ | – | – | – | – | – | – | – | – | – | + | – | – |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fijan, S.; Kocbek, P.; Steyer, A.; Vodičar, P.M.; Strauss, M. The Antimicrobial Effect of Various Single-Strain and Multi-Strain Probiotics, Dietary Supplements or Other Beneficial Microbes against Common Clinical Wound Pathogens. Microorganisms 2022, 10, 2518. https://doi.org/10.3390/microorganisms10122518
Fijan S, Kocbek P, Steyer A, Vodičar PM, Strauss M. The Antimicrobial Effect of Various Single-Strain and Multi-Strain Probiotics, Dietary Supplements or Other Beneficial Microbes against Common Clinical Wound Pathogens. Microorganisms. 2022; 10(12):2518. https://doi.org/10.3390/microorganisms10122518
Chicago/Turabian StyleFijan, Sabina, Primož Kocbek, Andrej Steyer, Polona Maver Vodičar, and Maja Strauss. 2022. "The Antimicrobial Effect of Various Single-Strain and Multi-Strain Probiotics, Dietary Supplements or Other Beneficial Microbes against Common Clinical Wound Pathogens" Microorganisms 10, no. 12: 2518. https://doi.org/10.3390/microorganisms10122518