Grinding Beads Influence Microbial DNA Extraction from Organic-Rich Sub-Seafloor Sediment


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
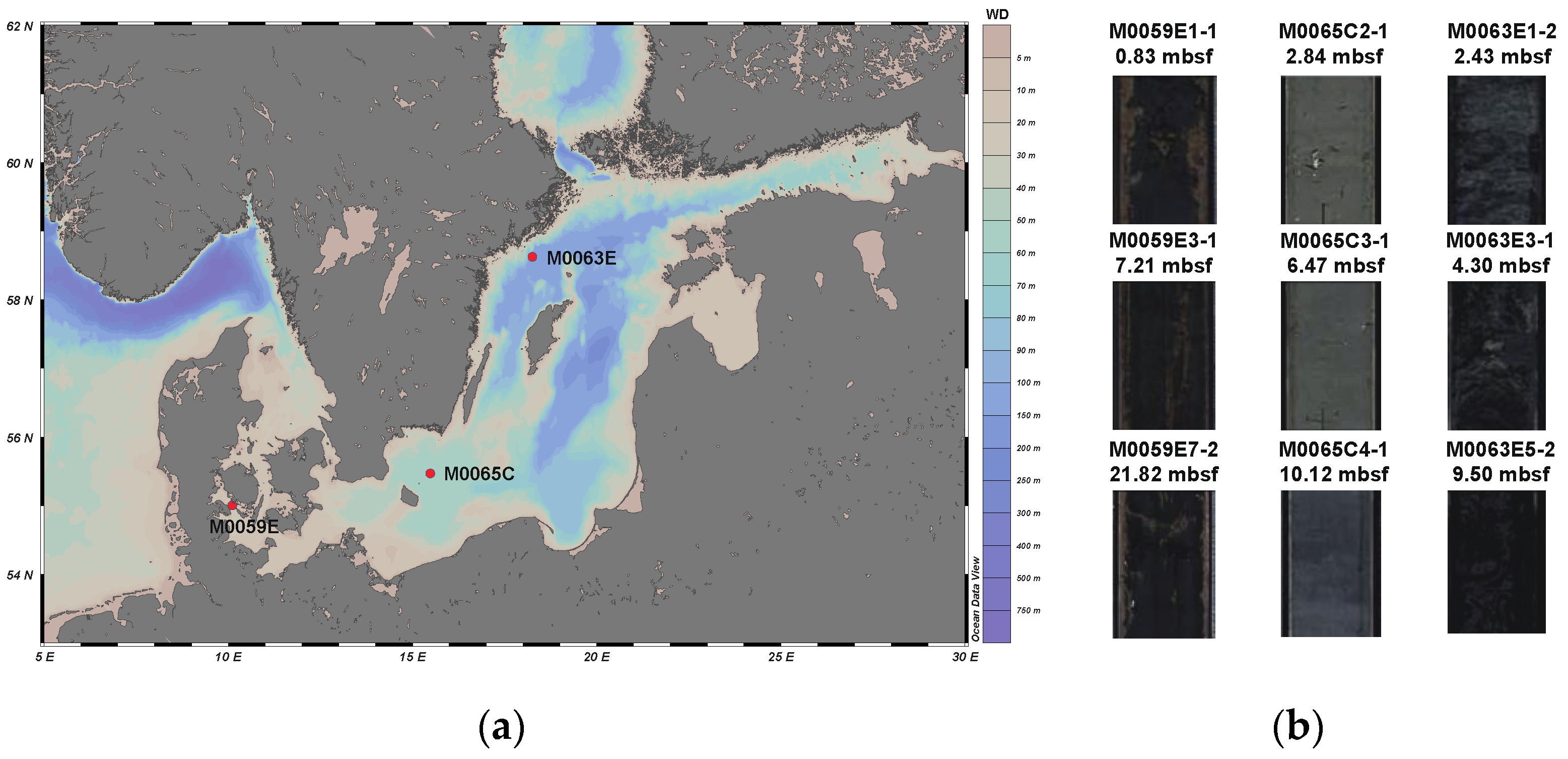
2.1. Sample Collection
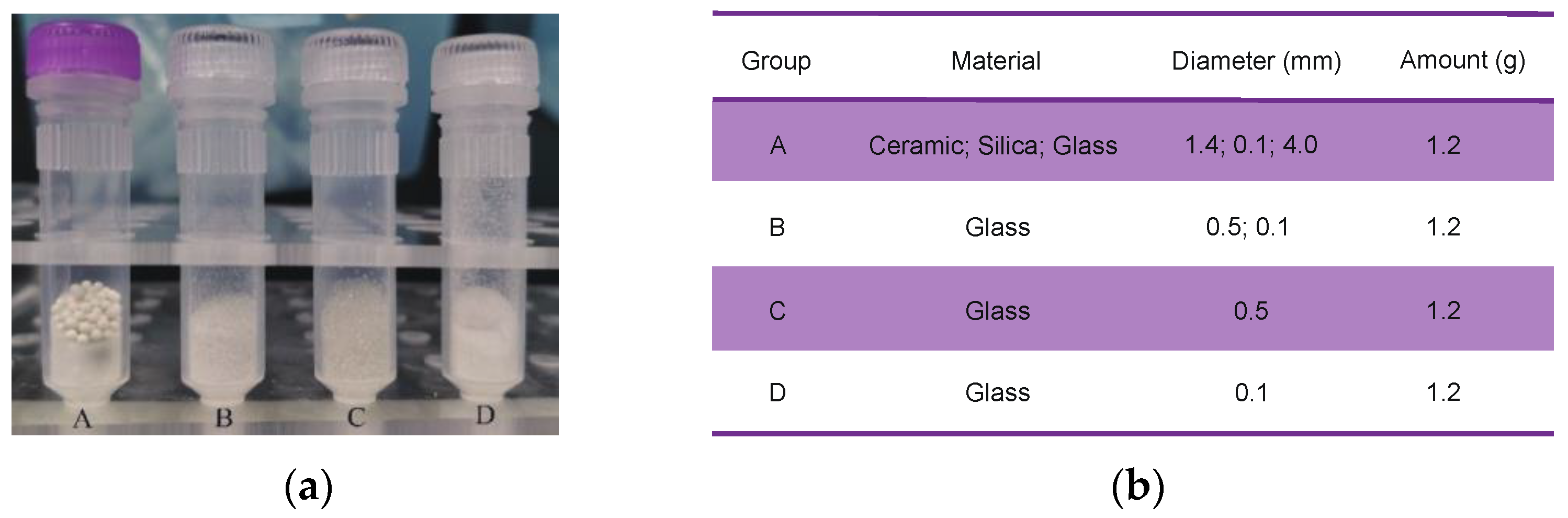
2.2. Combinations of Grinding Beads
2.3. DNA Extraction
- (I)
- Each sediment sample was thawed at 4 °C until it was soft enough to scrape off the surface with a sterile knife. Then, 0.5 g of the wet sediment sample was added to a 2-mL lysis tube with one of the groups of beads.
- (II)
- Lysis buffer (1 mL) containing 978 μL of sodium phosphate buffer and 122 μL of MT buffer was added to the lysis tube. The tube was then secured horizontally on a Vortex-Genie Adapter (Thermo Fisher Scientific, Waltham, MA, USA) and vortex mixed gently to dissolve the sediment and mix it with the beads and buffer.
- (III)
- The lysis tube was homogenized in a FastPrep instrument twice for 45 s at 6.0 m s−1 with a 5 min interval in an ice bath between the two cycles. This step was used to disrupt the cell wall and release nucleic acids.
- (IV)
- The sediment/bead mixture was centrifuged at 14,000× g for 10 min at 4 °C to pelletize any debris such as insoluble cellular debris and the lysis matrix.
- (V)
- The supernatant was transferred to a clean 2-mL microcentrifuge tube. PPS (250 μL) was added to separate the solubilized nucleic acids from the cellular debris and lysis matrix. After inverting the tube 10 times, it was centrifuged at 14,000× g for 5 min (4 °C).
- (VI)
- The supernatant was transferred to two clean 2-mL microcentrifuge tubes (approximately 0.5 mL each) and then resuspended with 0.5 mL of binding matrix. After vortex mixing, the tubes were placed in an ice bath for 15 min.
- (VII)
- For each of two tubes, 300 μL of the supernatant was carefully removed and discarded to avoid disturbing the settled binding matrix. The binding matrix in the remaining amount of supernatant was gently resuspended. The remaining 700 μL of the mixtures was transferred to a spin filter and centrifuged at 14,000× g for 2 min (4 °C). The spin filter was then emptied and reused for another tube of the mixture.
- (VIII)
- Prepared SEWS-M (500 μL with an appropriate volume of ethanol added) was added to solubilize any impurities. The pellet was gently resuspended using liquid from a pipet tip, which was followed by centrifugation at 14,000× g for 1 min (4 °C). The tube was then emptied. After washing the spin filter three times, it was centrifuged at 14,000× g for 2 min without any added liquid to completely remove SEWS-M.
- (IX)
- The catch tube was replaced with a new, clean catch tube, and the spin filter was air dried for 10 min at room temperature to remove the residual alcohol.
- (X)
- The binding matrix (above the spin filter) was gently resuspended in 100 μL of DES and incubated for 5 min at 55 °C in a heat block or water bath.
- (XI)
- The tube was centrifuged at 14,000× g for 1 min to bring the eluted DNA into the clean catch tube and the spin filter was discarded. The DNA samples were stored at −20 °C until required for amplification.
2.4. Analyses of Sequences and Statistics
3. Results
3.1. Sample Description
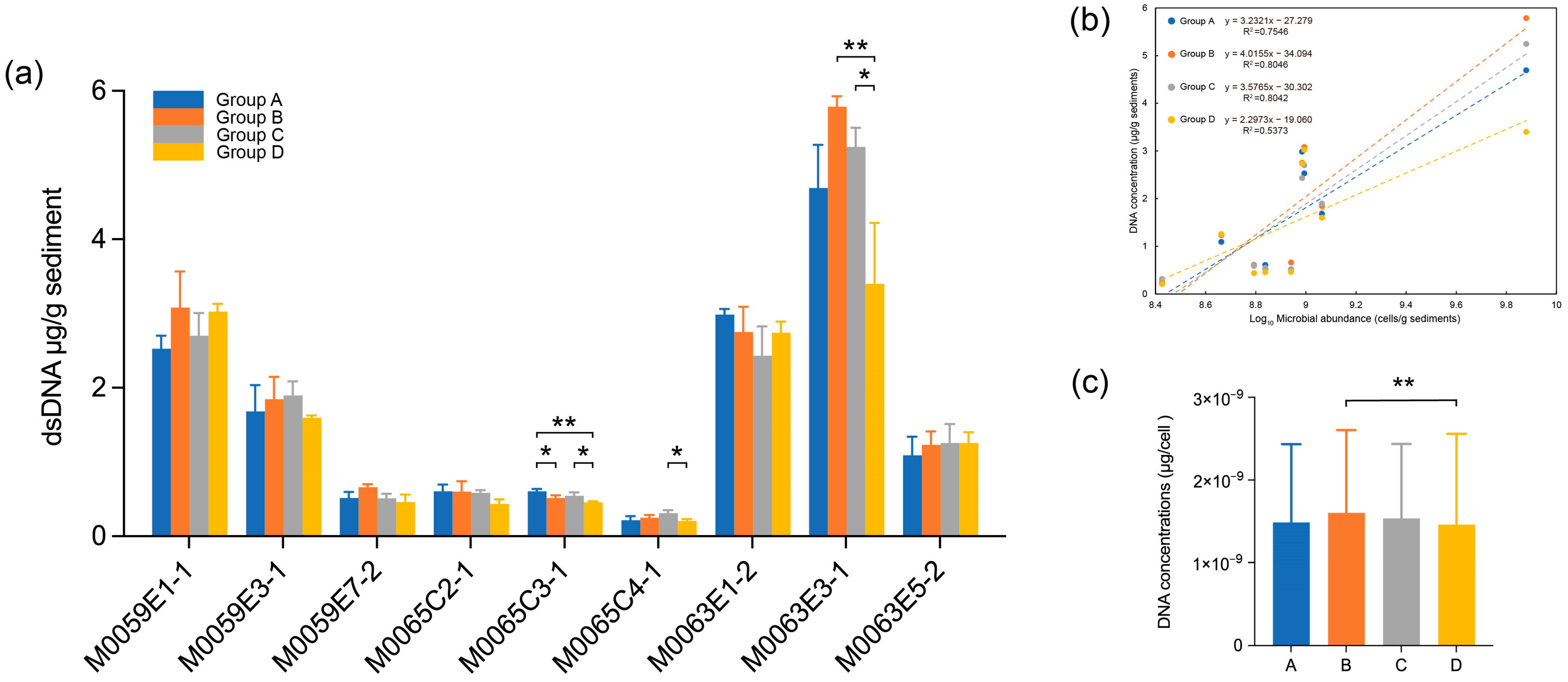
3.2. Effect of Grinding Beads on DNA Quality
3.3. Effect of Grinding Beads on the Microbial Community Alpha Diversity
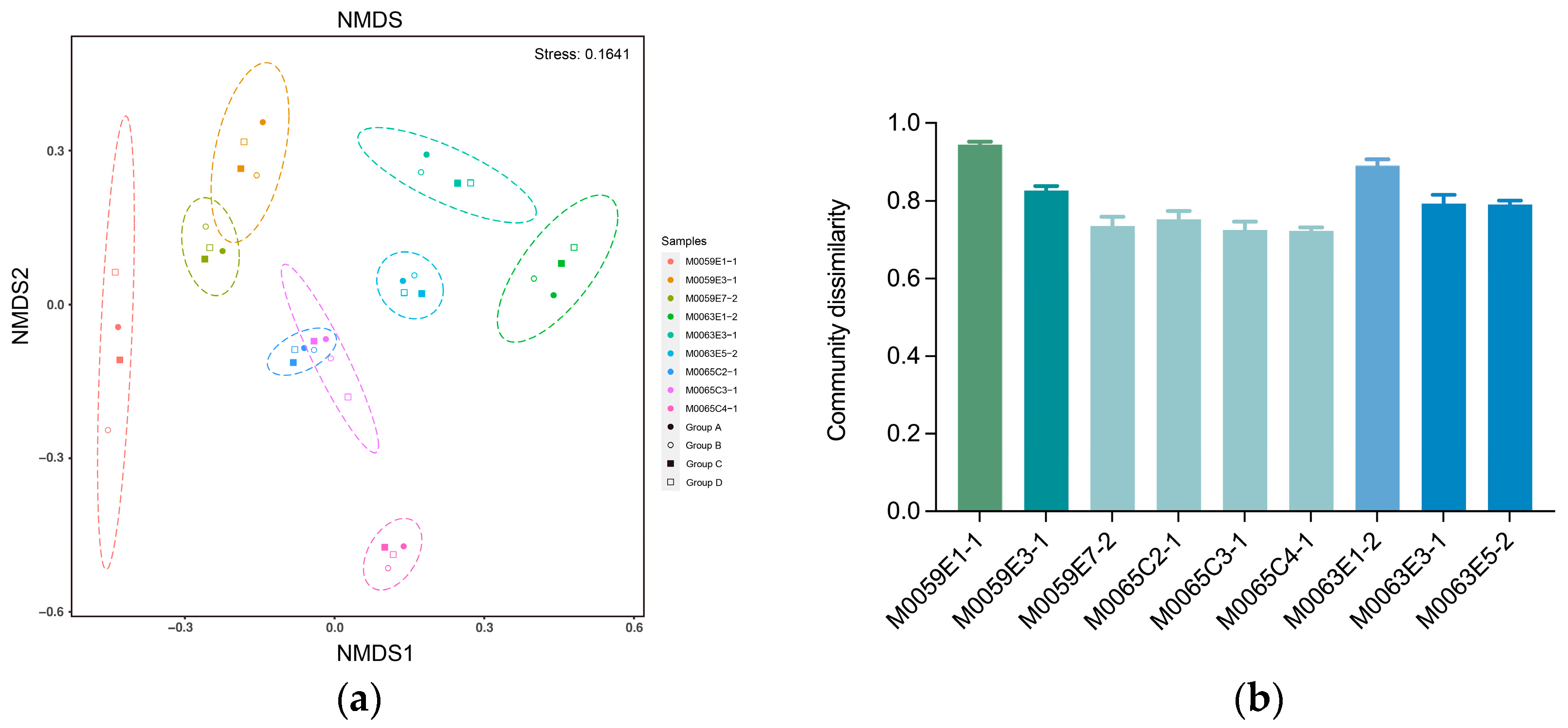
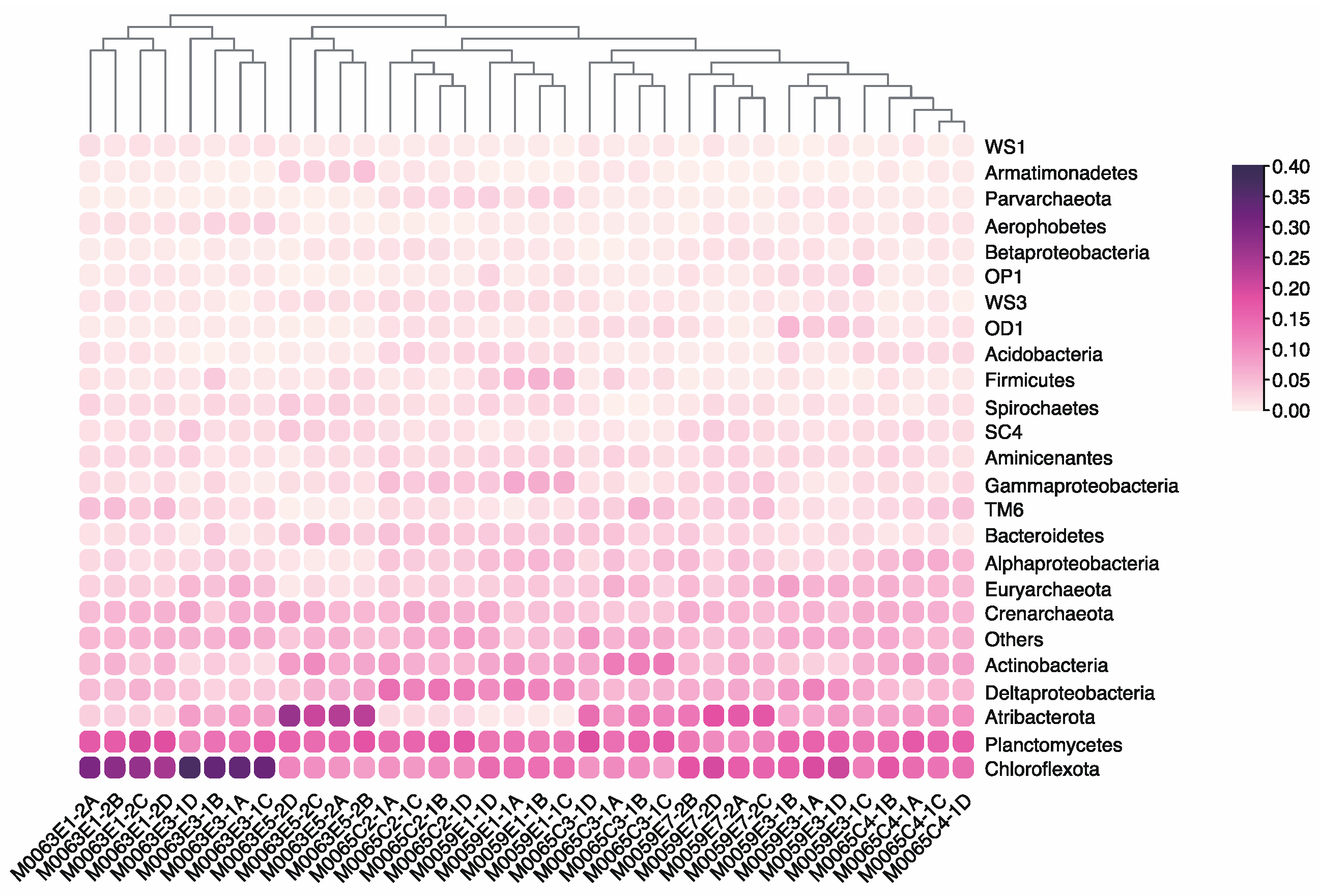
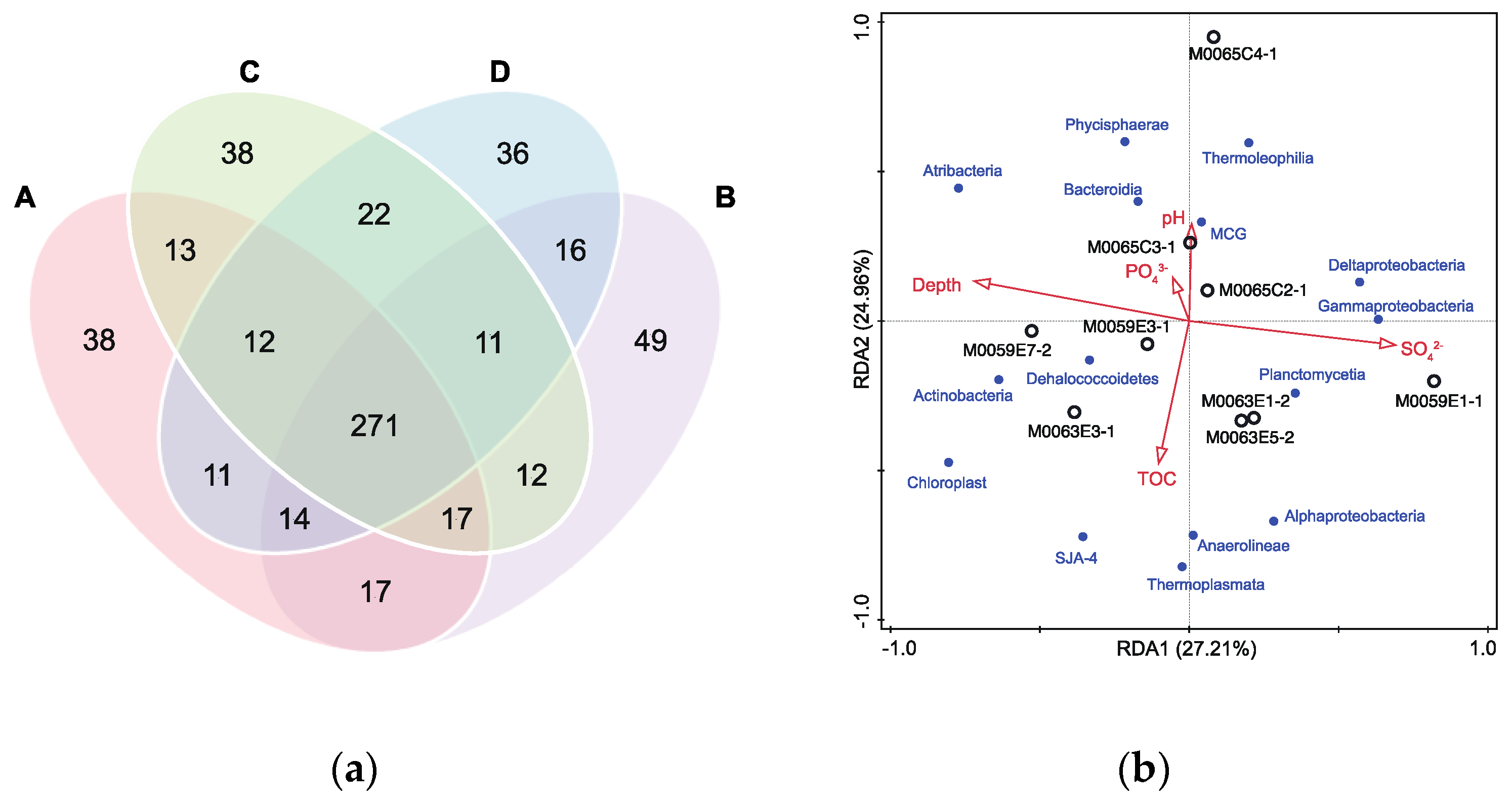
3.4. Effect of Grinding Beads on the Microbial Community Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martin-Laurent, F.; Philippot, L.; Hallet, S.; Chaussod, R.; Germon, J.C.; Soulas, G.; Catroux, G. DNA Extraction from Soils: Old Bias for New Microbial Diversity Analysis Methods. Appl. Environ. Microb. 2001, 67, 4397. [Google Scholar] [CrossRef] [Green Version]
- Lipp, J.S.; Morono, Y.; Inagaki, F.; Hinrichs, K.-U. Significant Contribution of Archaea to Extant Biomass in Marine Subsurface Sediments. Nature 2008, 454, 991–994. [Google Scholar] [CrossRef] [PubMed]
- Morono, Y.; Terada, T.; Hoshino, T.; Inagaki, F. Hot-Alkaline DNA Extraction Method for Deep-Subseafloor Archaeal Communities. Appl. Environ. Microb. 2014, 80, 1985–1994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrigg, C.; Rice, O.; Kavanagh, S.; Collins, G.; O’Flaherty, V. DNA Extraction Method Affects Microbial Community Profiles from Soils and Sediment. Appl. Microbiol. Biotechnol. 2007, 77, 955–964. [Google Scholar] [CrossRef] [PubMed]
- Morgan, J.L.; Darling, A.E.; Eisen, J.A. Metagenomic Sequencing of an In Vitro-Simulated Microbial Community. PLoS ONE 2010, 5, e10209. [Google Scholar] [CrossRef] [PubMed]
- Salonen, A.; Nikkilä, J.; Jalanka-Tuovinen, J.; Immonen, O.; Rajilić-Stojanović, M.; Kekkonen, R.A.; Palva, A.; de Vos, W.M. Comparative Analysis of Fecal DNA Extraction Methods with Phylogenetic Microarray: Effective Recovery of Bacterial and Archaeal DNA Using Mechanical Cell Lysis. J. Microbiol. Methods 2010, 81, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Ariefdjohan, M.W.; Savaiano, D.A.; Nakatsu, C.H. Comparison of DNA Extraction Kits for PCR-DGGE Analysis of Human Intestinal Microbial Communities from Fecal Specimens. Nutr. J. 2010, 9, 23. [Google Scholar] [CrossRef] [Green Version]
- Scupham, A.J.; Jones, J.A.; Wesley, I.V. Comparison of DNA Extraction Methods for Analysis of Turkey Cecal Microbiota. J. Appl. Microbiol. 2007, 102, 401–409. [Google Scholar] [CrossRef]
- Inceoglu, O.; Hoogwout, E.F.; Hill, P.; Elsas, J.D. van Effect of DNA Extraction Method on the Apparent Microbial Diversity of Soil. Appl. Environ. Microb. 2010, 76, 3378–3382. [Google Scholar] [CrossRef] [Green Version]
- Yuan, S.; Cohen, D.B.; Ravel, J.; Abdo, Z.; Forney, L.J. Evaluation of Methods for the Extraction and Purification of DNA from the Human Microbiome. PLoS ONE 2012, 7, e33865. [Google Scholar] [CrossRef]
- Guo, F.; Zhang, T. Biases during DNA Extraction of Activated Sludge Samples Revealed by High Throughput Sequencing. Appl. Microbiol. Biotechnol. 2013, 97, 4607–4616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alain, K.; Callac, N.; Ciobanu, M.-C.; Reynaud, Y.; Duthoit, F.; Jebbar, M. DNA Extractions from Deep Subseafloor Sediments: Novel Cryogenic-Mill-Based Procedure and Comparison to Existing Protocols. J. Microbiol. Methods 2011, 87, 355–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reed, D.W.; Fujita, Y.; Delwiche, M.E.; Blackwelder, D.B.; Sheridan, P.P.; Uchida, T.; Colwell, F.S. Microbial Communities from Methane Hydrate-Bearing Deep Marine Sediments in a Forearc Basin. Appl. Environ. Microbiol. 2002, 68, 3759–3770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roose-Amsaleg, C.L.; Garnier-Sillam, E.; Harry, M. Extraction and Purification of Microbial DNA from Soil and Sediment Samples. Appl. Soil Ecol. 2001, 18, 47–60. [Google Scholar] [CrossRef]
- Leff, L.G.; Dana, J.R.; McArthur, J.V.; Shimkets, L.J. Comparison of Methods of DNA Extraction from Stream Sediments. Appl. Environ. Microb. 1995, 61, 1141–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Courtois, S.; Frostegård, Å.; Göransson, P.; Depret, G.; Jeannin, P.; Simonet, P. Quantification of Bacterial Subgroups in Soil: Comparison of DNA Extracted Directly from Soil or from Cells Previously Released by Density Gradient Centrifugation. Environ. Microbiol. 2001, 3, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Luna, G.M.; Dell’Anno, A.; Danovaro, R. DNA Extraction Procedure: A Critical Issue for Bacterial Diversity Assessment in Marine Sediments. Environ. Microbiol. 2006, 8, 308–320. [Google Scholar] [CrossRef]
- Steffan, R.J.; Goksøyr, J.; Bej, A.K.; Atlas, R.M. Recovery of DNA from Soils and Sediments. Appl. Environ. Microbiol. 1988, 54, 2908–2915. [Google Scholar] [CrossRef] [Green Version]
- Lindahl, V. Improved Soil Dispersion Procedures for Total Bacterial Counts, Extraction of Indigenous Bacteria and Cell Survival. J. Microbiol. Methods 1996, 25, 279–286. [Google Scholar] [CrossRef]
- Gabor, E.M.; Vries, E.J.; Janssen, D.B. Efficient Recovery of Environmental DNA for Expression Cloning by Indirect Extraction Methods. FEMS Microbiol. Ecol. 2003, 44, 153–163. [Google Scholar] [CrossRef]
- Saidi-Mehrabad, A.; Neuberger, P.; Cavaco, M.; Froese, D.; Lanoil, B. Optimization of Subsampling, Decontamination, and DNA Extraction of Difficult Peat and Silt Permafrost Samples. Sci. Rep. 2020, 10, 14295. [Google Scholar] [CrossRef] [PubMed]
- Kuske, C.R.; Banton, K.L.; Adorada, D.L.; Stark, P.C.; Hill, K.K.; Jackson, P.J. Small-Scale DNA Sample Preparation Method for Field PCR Detection of Microbial Cells and Spores in Soil. Appl. Environ. Microb. 1998, 64, 2463–2472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robe, P.; Nalin, R.; Capellano, C.; Vogel, T.M.; Simonet, P. Extraction of DNA from Soil. Eur. J. Soil Biol. 2003, 39, 183–190. [Google Scholar] [CrossRef]
- Moré, M.I.; Herrick, J.B.; Silva, M.C.; Ghiorse, W.C.; Madsen, E.L. Quantitative Cell Lysis of Indigenous Microorganisms and Rapid Extraction of Microbial DNA from Sediment. Appl. Environ. Microbiol. 1994, 60, 1572–1580. [Google Scholar] [CrossRef] [Green Version]
- Whitehouse, C.A.; Hottel, H.E. Comparison of Five Commercial DNA Extraction Kits for the Recovery of Francisella Tularensis DNA from Spiked Soil Samples. Mol. Cell Probe 2007, 21, 92–96. [Google Scholar] [CrossRef]
- Vishnivetskaya, T.A.; Layton, A.C.; Lau, M.C.Y.; Chauhan, A.; Cheng, K.R.; Meyers, A.J.; Murphy, J.R.; Rogers, A.W.; Saarunya, G.S.; Williams, D.E.; et al. Commercial DNA Extraction Kits Impact Observed Microbial Community Composition in Permafrost Samples. FEMS Microbiol. Ecol. 2014, 87, 217–230. [Google Scholar] [CrossRef]
- Miller, D.N.; Bryant, J.E.; Madsen, E.L.; Ghiorse, W.C. Evaluation and Optimization of DNA Extraction and Purification Procedures for Soil and Sediment Samples. Appl. Environ. Microbiol. 1999, 65, 4715–4724. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Brock, M.; Arana, C.; Dende, C.; van Oers, N.S.; Hooper, L.V.; Raj, P. Impact of Bead-Beating Intensity on the Genus- and Species-Level Characterization of the Gut Microbiome Using Amplicon and Complete 16S RRNA Gene Sequencing. Front. Cell Infect. Microbiol. 2021, 11, 678522. [Google Scholar] [CrossRef]
- Bürgmann, H.; Pesaro, M.; Widmer, F.; Zeyer, J. A Strategy for Optimizing Quality and Quantity of DNA Extracted from Soil. J. Microbiol. Methods 2001, 45, 7–20. [Google Scholar] [CrossRef]
- Vanysacker, L.; Declerck, S.A.J.; Hellemans, B.; Meester, L.D.; Vankelecom, I.; Declerck, P. Bacterial Community Analysis of Activated Sludge: An Evaluation of Four Commonly Used DNA Extraction Methods. Appl. Microbiol. Biotechnol. 2010, 88, 299–307. [Google Scholar] [CrossRef]
- Lever, M.A.; Torti, A.; Eickenbusch, P.; Michaud, A.B.; Šantl-Temkiv, T.; Jørgensen, B.B. A Modular Method for the Extraction of DNA and RNA, and the Separation of DNA Pools from Diverse Environmental Sample Types. Front. Microbiol. 2015, 6, 476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrén, T.; Björck, S.; Andrén, E.; Conley, D.; Zillén, L.; Anjar, J. The Baltic Sea Basin; Central and East European Development Studies (CEEDES); Springer: Berlin/Heidelberg, Germany, 2011; pp. 75–97. [Google Scholar]
- Andrén, T.; Jørgensen, B.B.; Cotterill, C.; Green, S.; the IODP Expedition 347 Scientific Party. IODP Expedition 347: Baltic Sea Basin Paleoenvironment and Biosphere. Sci. Drill. 2015, 20, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Andrén, T.; Jørgensen, B.B.; Cotterill, C.; the IODP347 Scientists. Baltic Sea paleoenvironment. In Proceedings of the Integrated Ocean Drilling Program, Kiel, Germany, 12 September–1 November 2014; Volume 347. [Google Scholar]
- Hardisty, D.S.; Riedinger, N.; Planavsky, N.J.; Asael, D.; Andren, T.; Jorgensen, B.B.; Lyons, T.W. A Holocene History of Dynamic Water Column Redox Conditions in the Landsort Deep, Baltic Sea. Am. J. Sci. 2016, 316, 713–745. [Google Scholar] [CrossRef] [Green Version]
- Parada, A.E.; Needham, D.M.; Fuhrman, J.A. Every Base Matters: Assessing Small Subunit RRNA Primers for Marine Microbiomes with Mock Communities, Time Series and Global Field Samples. Environ. Microbiol. 2015, 18, 1403–1414. [Google Scholar] [CrossRef] [PubMed]
- Apprill, A.; McNally, S.; Parsons, R.; Weber, L. Minor Revision to V4 Region SSU RRNA 806R Gene Primer Greatly Increases Detection of SAR11 Bacterioplankton. Aquat. Microbiol. Ecol. 2015, 75, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME Allows Analysis of High-Throughput Community Sequencing Data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-Filtering Vastly Improves Diversity Estimates from Illumina Amplicon Sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2015, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 4–9. [Google Scholar]
- Robeson, M.S.; O’Rourke, D.R.; Kaehler, B.D.; Ziemski, M.; Dillon, M.R.; Foster, J.T.; Bokulich, N.A. RESCRIPt: Reproducible Sequence Taxonomy Reference Database Management for the Masses. bioRxiv 2020, 17, e1009581. [Google Scholar]
- Cai, L.; Jørgensen, B.B.; Suttle, C.A.; He, M.; Cragg, B.A.; Jiao, N.; Zhang, R. Active and Diverse Viruses Persist in the Deep Sub-Seafloor Sediments over Thousands of Years. ISME J. 2019, 13, 1857–1864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maukonen, J.; Simões, C.; Saarela, M. The Currently Used Commercial DNA−extraction Methods Give Different Results of Clostridial and Actinobacterial Populations Derived from Human Fecal Samples. FEMS Microbiol. Ecol. 2012, 79, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Ferrand, J.; Patron, K.; Legrand-Frossi, C.; Frippiat, J.-P.; Merlin, C.; Alauzet, C.; Lozniewski, A. Comparison of Seven Methods for Extraction of Bacterial DNA from Fecal and Cecal Samples of Mice. J. Microbiol. Methods 2014, 105, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Knauth, S.; Schmidt, H.; Tippkötter, R. Comparison of Commercial Kits for the Extraction of DNA from Paddy Soils. Lett. Appl. Microbiol. 2013, 56, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Jarosz, W.; Durant, J.-F.; Irenge, L.M.W.B.; Fogt-Wyrwas, R.; Mizgajska-Wiktor, H.; Gala, J.-L. Optimized DNA-Based Identification of Toxocara spp. Eggs in Soil and Sand Samples. Parasite Vector 2021, 14, 426. [Google Scholar] [CrossRef]
- Jørgensen, B.B.; Andrén, T.; Marshall, I.P.G. Sub−seafloor Biogeochemical Processes and Microbial Life in the Baltic Sea. Environ. Microbiol. 2020, 22, 1688–1706. [Google Scholar] [CrossRef] [Green Version]
- Frostegård, Å.; Courtois, S.; Ramisse, V.; Clerc, S.; Bernillon, D.; Gall, F.L.; Jeannin, P.; Nesme, X.; Simonet, P. Quantification of Bias Related to the Extraction of DNA Directly from Soils. Appl. Environ. Microb. 1999, 65, 5409–5420. [Google Scholar] [CrossRef] [Green Version]
- Gobbi, A.; Santini, R.G.; Filippi, E.; Ellegaard-Jensen, L.; Jacobsen, C.S.; Hansen, L.H. Quantitative and Qualitative Evaluation of the Impact of the G2 Enhancer, Bead Sizes and Lysing Tubes on the Bacterial Community Composition during DNA Extraction from Recalcitrant Soil Core Samples Based on Community Sequencing and QPCR. PLoS ONE 2019, 14, e0200979. [Google Scholar] [CrossRef] [Green Version]
- Rantakokko-Jalava, K.; Jalava, J. Optimal DNA Isolation Method for Detection of Bacteria in Clinical Specimens by Broad-Range PCR. J. Clin. Microbiol. 2002, 40, 4211–4217. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Bruns, M.A.; Tiedje, J.M. DNA Recovery from Soils of Diverse Composition. Appl. Environ. Microbiol. 1996, 62, 316–322. [Google Scholar] [CrossRef] [Green Version]
- Glombitza, C.; Egger, M.; Røy, H.; Jørgensen, B.B. Controls on Volatile Fatty Acid Concentrations in Marine Sediments (Baltic Sea). Geochim. Cosmochim. Acta 2019, 258, 226–241. [Google Scholar] [CrossRef]
- Ryan, R.P.; Monchy, S.; Cardinale, M.; Taghavi, S.; Crossman, L.; Avison, M.B.; Berg, G.; van der Lelie, D.; Dow, J.M. The Versatility and Adaptation of Bacteria from the Genus Stenotrophomonas. Nat. Rev. Microbiol. 2009, 7, 514–525. [Google Scholar] [CrossRef] [PubMed]
- Volossiouk, T.; Robb, E.J.; Nazar, R.N. Direct DNA Extraction for PCR-Mediated Assays of Soil Organisms. Appl. Environ. Microbiol. 1995, 61, 3972–3976. [Google Scholar] [CrossRef] [Green Version]
- Hasty, D.L.; Ofek, I.; Courtney, H.S.; Doyle, R.J. Multiple Adhesins of Streptococci. Infect. Immun. 1992, 60, 2147–2152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birkeland, N.K.; Hofstad, T. Oligosaccharides Obtained by Partial Hydrolysis of Lipopolysaccharides from Leptotrichia Buccalis. Eur. J. Oral Sci. 1985, 93, 432–435. [Google Scholar] [CrossRef]
- Knudsen, B.E.; Bergmark, L.; Munk, P.; Lukjancenko, O.; Priemé, A.; Aarestrup, F.M.; Pamp, S.J. Impact of Sample Type and DNA Isolation Procedure on Genomic Inference of Microbiome Composition. Msystems 2016, 1, e00095-16. [Google Scholar] [CrossRef] [Green Version]
- Nobu, M.K.; Dodsworth, J.A.; Murugapiran, S.K.; Rinke, C.; Gies, E.A.; Webster, G.; Schwientek, P.; Kille, P.; Parkes, R.J.; Sass, H.; et al. Phylogeny and Physiology of Candidate Phylum ‘Atribacteria’ (OP9/JS1) Inferred from Cultivation-Independent Genomics. ISME J. 2016, 10, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Parkes, R.J.; Cragg, B.; Roussel, E.; Webster, G.; Weightman, A.; Sass, H. A Review of Prokaryotic Populations and Processes in Sub-Seafloor Sediments, Including Biosphere: Geosphere Interactions. Mar. Geol. 2014, 352, 409–425. [Google Scholar] [CrossRef]
- Fullerton, H.; Moyer, C.L. Comparative Single-Cell Genomics of Chloroflexi from the Okinawa Trough Deep-Subsurface Biosphere. Appl. Environ. Microbiol. 2016, 82, 3000–3008. [Google Scholar] [CrossRef] [Green Version]
- Petro, C.; Zäncker, B.; Starnawski, P.; Jochum, L.M.; Ferdelman, T.G.; Jørgensen, B.B.; Røy, H.; Kjeldsen, K.U.; Schramm, A. Marine Deep Biosphere Microbial Communities Assemble in Near-Surface Sediments in Aarhus Bay. Front. Microbiol. 2019, 10, 758. [Google Scholar] [CrossRef] [Green Version]
- Puente-Sánchez, F.; Arce-Rodríguez, A.; Oggerin, M.; García-Villadangos, M.; Moreno-Paz, M.; Blanco, Y.; Rodríguez, N.; Bird, L.; Lincoln, S.A.; Tornos, F.; et al. Viable Cyanobacteria in the Deep Continental Subsurface. Proc. Natl. Acad. Sci. USA 2018, 115, 10702–10707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchi, T.S.; Engelhaupt, E.; Westman, P.; Andrén, T.; Rolff, C.; Elmgren, R. Cyanobacterial Blooms in the Baltic Sea: Natural or Human−induced? Limnol. Oceanogr. 2000, 45, 716–726. [Google Scholar] [CrossRef]
- Poutanen, E.-L.; Nikkil, K. Carotenoid Pigments as Tracers of Cyanobacterial Blooms in Recent and Post-Glacial Sediments of the Baltic Sea. Ambio J. Hum. Environ. 2001, 30, 179–183. [Google Scholar] [CrossRef]
- Funkey, C.P.; Conley, D.J.; Reuss, N.S.; Humborg, C.; Jilbert, T.; Slomp, C.P. Hypoxia Sustains Cyanobacteria Blooms in the Baltic Sea. Environ. Sci. Technol. 2014, 48, 2598–2602. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, J.; Chen, H.; Cai, L.; He, M.; Zhang, R.; Wang, L. Grinding Beads Influence Microbial DNA Extraction from Organic-Rich Sub-Seafloor Sediment. Microorganisms 2022, 10, 2505. https://doi.org/10.3390/microorganisms10122505
Niu J, Chen H, Cai L, He M, Zhang R, Wang L. Grinding Beads Influence Microbial DNA Extraction from Organic-Rich Sub-Seafloor Sediment. Microorganisms. 2022; 10(12):2505. https://doi.org/10.3390/microorganisms10122505
Chicago/Turabian StyleNiu, Jingjing, Hong Chen, Lanlan Cai, Maoqiu He, Rui Zhang, and Long Wang. 2022. "Grinding Beads Influence Microbial DNA Extraction from Organic-Rich Sub-Seafloor Sediment" Microorganisms 10, no. 12: 2505. https://doi.org/10.3390/microorganisms10122505