
The Use of Interdisciplinary Approaches to Understand the Biology of Campylobacter jejuni
 , , and
, , and {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Microorganism Level
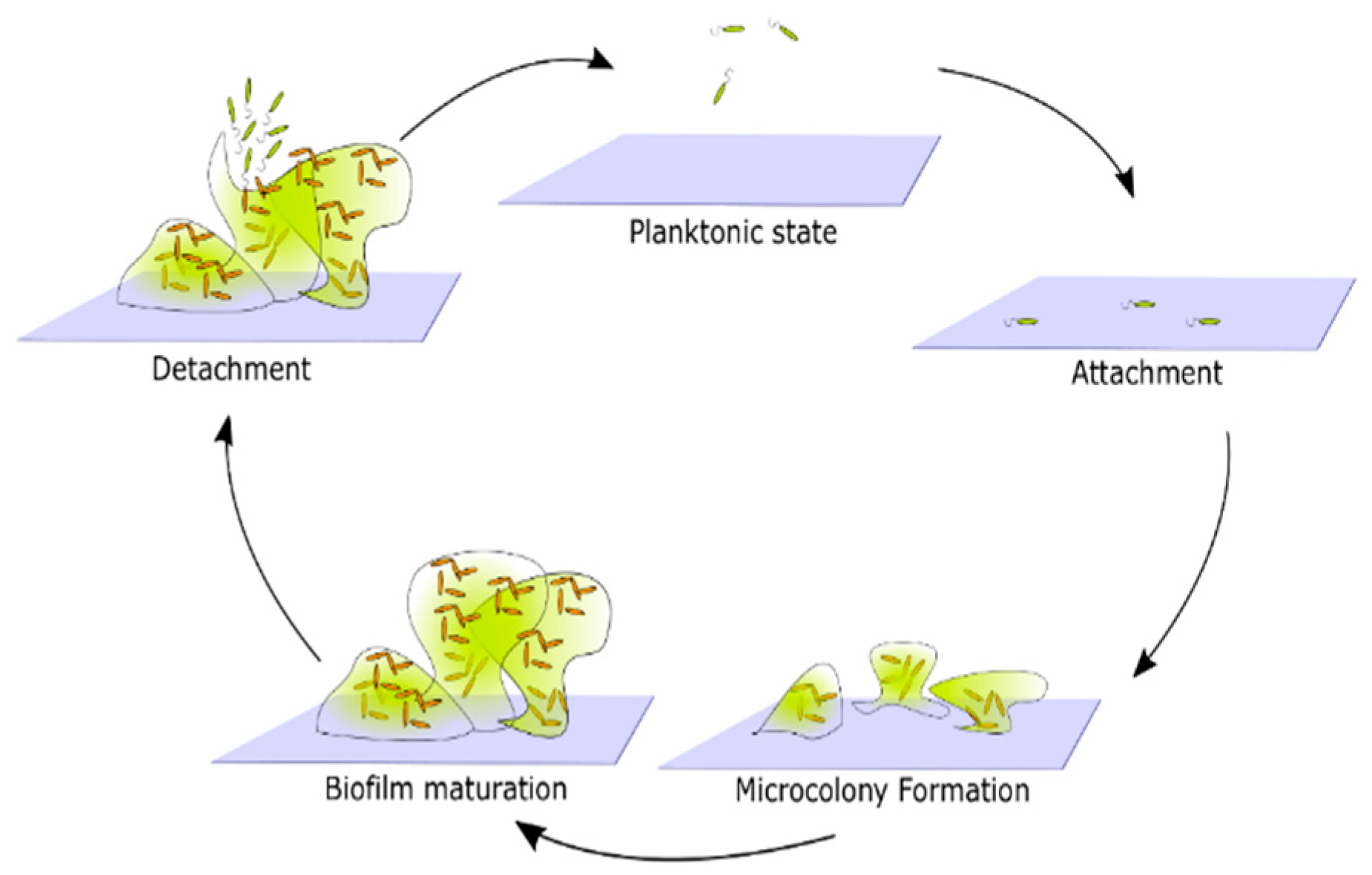
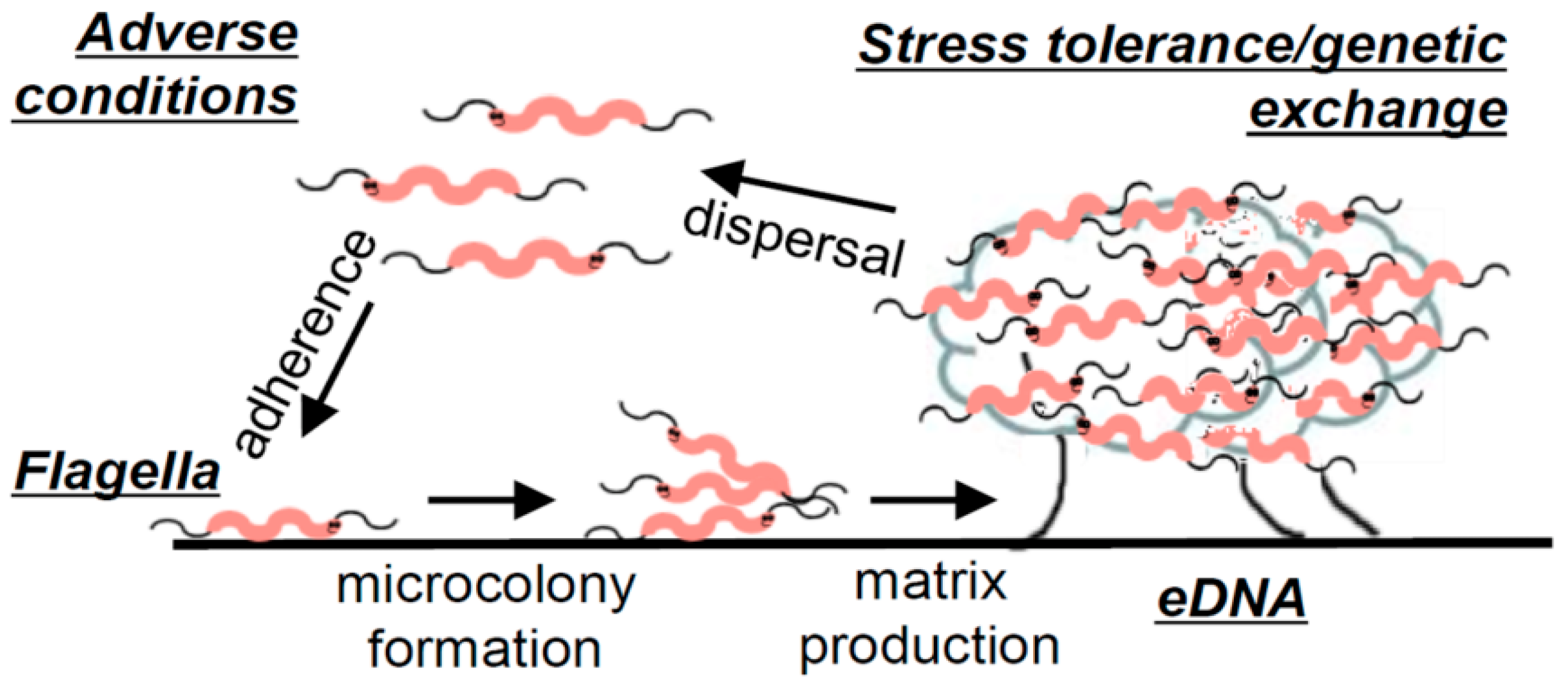
2.1. Biofilm Formation Studies
2.2. Predictive Studies for C. jejuni Survival
2.3. Metabolic Modelling and Growth Requirements
3. Animal Infection Model Level
4. Epidemiological Studies at the Host Population Level
5. Summary
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Campylobacter (Key Facts). Available online: https://www.who.int/news-room/fact-sheets/detail/campylobacter (accessed on 29 August 2021).
- Sheppard, S.K.; Maiden, M.C.J. The evolution of Campylobacter jejuni and Campylobacter coli. Cold Spring Harb. Perspect. Biol. 2015, 7, a018119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cody, A.J.; Maiden, M.C.; Strachan, N.J.; McCarthy, N.D. A systematic review of source attribution of human campylobacteriosis using multilocus sequence typing. Eurosurveillance 2019, 24, 1800696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tack, D.M.; Ray, L.; Griffin, P.M.; Cieslak, P.R.; Dunn, J.; Rissman, T.; Jervis, R.; Lathrop, S.; Muse, A.; Duwell, M.; et al. Preliminary Incidence and Trends of Infections with Pathogens Transmitted Commonly Through Food—Foodborne Diseases Active Surveillance Network, 10 U.S. Sites, 2016–2019. MMWR Morb. Mortal Wkly. Rep. 2020, 69, 509–514. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union One Health 2019 Zoonoses Report. EFSA J. 2021, 19, e06406. [Google Scholar] [CrossRef]
- Fonseca, B.B.; Fernandez, H.; Rossi, D.A. Campylobacter spp. and Related Organisms in Poultry; Springer International Publishing: Minas Gerais, Brazil; Valdivia, Chile, 2016; Available online: https://link.springer.com/content/pdf/10.1007/978-3-319-29907-5.pdf (accessed on 29 August 2017).
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2014. EFSA J. 2015, 13, 4329. [Google Scholar] [CrossRef]
- Adak, G.K.; Meakins, S.M.; Yip, H.; Lopman, B.A.; O’Brien, S.J. Disease Risks from Foods, England and Wales, 1996–2000. Emerg. Infect. Dis. 2005, 11, 365–372. [Google Scholar] [CrossRef]
- Walker, L.J.; Wallace, R.L.; Smith, J.J.; Graham, T.; Saputra, T.; Symes, S.; Stylianopoulos, A.; Polkinghorne, B.G.; Kirk, M.D.; Glass, K. Prevalence of Campylobacter coli and Campylobacter jejuni in Retail Chicken, Beef, Lamb, and Pork Products in Three Australian States. J. Food Prot. 2019, 82, 2126–2134. [Google Scholar] [CrossRef]
- Shagieva, E.; Demnerova, K.; Michova, H. Waterborne Isolates of Campylobacter jejuni Are Able to Develop Aerotolerance, Survive Exposure to Low Temperature, and Interact with Acanthamoeba polyphaga. Front. Microbiol. 2021, 12, 3162. [Google Scholar] [CrossRef]
- Magajna, B.A.; Schraft, H. Campylobacter jejuni biofilm cells become viable but non-culturable (VBNC) in low nutrient conditions at 4 °C more quickly than their planktonic counterparts. Food Control 2015, 50, 45–50. [Google Scholar] [CrossRef]
- Nasher, F.; Lehri, B.; Horney, M.F.; Stabler, R.A.; Wren, B.W. Survival of Campylobacter jejuni 11168H in Acanthamoebae castellanii Provides Mechanistic Insight into Host Pathogen Interactions. Microorganisms 2022, 10, 1894. [Google Scholar] [CrossRef]
- Hofreuter, D. Defining the metabolic requirements for the growth and colonization capacity of Campylobacter jejuni. Front. Cell. Infect. Microbiol. 2014, 4, 137. [Google Scholar] [CrossRef] [Green Version]
- Svensson, S.; Pryjma, M.; Gaynor, E.C. Flagella-Mediated Adhesion and Extracellular DNA Release Contribute to Biofilm Formation and Stress Tolerance of Campylobacter jejuni. PLoS ONE 2014, 9, e106063. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, Z.; Smith, B.; Fazil, A.; Wu, J.; Ryan, S.D.; Munther, D. pH dependent C. jejuni thermal inactivation models and application to poultry scalding. J. Food Eng. 2018, 223, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Metris, A.; Reuter, M.; Gaskin, D.J.H.; Baranyi, J.; Van Vliet, A.H.M. In vivo and in silico determination of essential genes of Campylobacter jejuni. BMC Genom. 2011, 12, 535. [Google Scholar] [CrossRef] [Green Version]
- Richelle, A.; David, B.; Demaegd, D.; Dewerchin, M.; Kinet, R.; Morreale, A.; Portela, R.; Zune, Q.; Von Stosch, M. Towards a widespread adoption of metabolic modeling tools in biopharmaceutical industry: A process systems biology engineering perspective. npj Syst. Biol. Appl. 2020, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Damte, D.; Suh, J.-W.; Lee, S.-J.; Yohannes, S.B.; Hossain, A.; Park, S.-C. Putative drug and vaccine target protein identification using comparative genomic analysis of KEGG annotated metabolic pathways of Mycoplasma hyopneumoniae. Genomics 2013, 102, 47–56. [Google Scholar] [CrossRef] [Green Version]
- Ferrarini, M.G.; Siqueira, F.M.; Mucha, S.G.; Palama, T.L.; Jobard, É.; Elena-Herrmann, B.; Vasconcelos, A.T.R.; Tardy, F.; Schrank, I.S.; Zaha, A.; et al. Insights on the virulence of swine respiratory tract mycoplasmas through genome-scale metabolic modeling. BMC Genom. 2016, 17, 353. [Google Scholar] [CrossRef] [Green Version]
- Giallourou, N.; Medlock, G.L.; Bolick, D.T.; Medeiros, P.H.; Ledwaba, S.E.; Kolling, G.L.; Tung, K.; Guerry, P.; Swann, J.R.; Guerrant, R.L. A novel mouse model of Campylobacter jejuni enteropathy and diarrhea. PLoS Pathog. 2018, 14, e1007083. [Google Scholar] [CrossRef] [Green Version]
- Cha, W.; Henderson, T.; Collins, J.; Manning, S.D. Factors associated with increasing campylobacteriosis incidence in Michigan, 2004–2013. Epidemiol. Infect. 2016, 144, 3316–3325. [Google Scholar] [CrossRef] [Green Version]
- Gradel, K.O.; Nielsen, H.L.; Schønheyder, H.C.; Ejlertsen, T.; Kristensen, B.; Nielsen, H. Increased Short- and Long-Term Risk of Inflammatory Bowel Disease After Salmonella or Campylobacter Gastroenteritis. Gastroenterology 2009, 137, 495–501. [Google Scholar] [CrossRef]
- Joshua, G.W.P.; Guthrie-Irons, C.; Karlyshev, A.V.; Wren, B.W. Biofilm formation in Campylobacter jejuni. Microbiology 2006, 152, 387–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, J.; Oh, E.; Jeon, B. Enhanced Transmission of Antibiotic Resistance in Campylobacter jejuni Biofilms by Natural Transformation. Antimicrob. Agents Chemother. 2014, 58, 7573–7575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Toole, G.; Kaplan, H.B.; Kolter, R. Biofilm Formation as Microbial Development. Annu. Rev. Microbiol. 2000, 54, 49–79. [Google Scholar] [CrossRef] [PubMed]
- Reeser, R.J.; Medler, R.T.; Billington, S.J.; Jost, B.H.; Joens, L.A. Characterization of Campylobacter jejuni Biofilms under Defined Growth Conditions. Appl. Environ. Microbiol. 2007, 73, 1908–1913. [Google Scholar] [CrossRef] [Green Version]
- Kalmokoff, M.; Lanthier, P.; Tremblay, T.-L.; Foss, M.; Lau, P.C.; Sanders, G.; Austin, J.; Kelly, J.; Szymanski, C.M. Proteomic Analysis of Campylobacter jejuni 11168 Biofilms Reveals a Role for the Motility Complex in Biofilm Formation. J. Bacteriol. 2006, 188, 4312–4320. [Google Scholar] [CrossRef] [Green Version]
- Brown, H.L.; Reuter, M.; Salt, L.J.; Cross, K.L.; Betts, R.P.; van Vliet, A.H.M. Chicken Juice Enhances Surface Attachment and Biofilm Formation of Campylobacter jejuni. Appl. Environ. Microbiol. 2014, 80, 7053–7060. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Ma, L.; Nie, J.; Konkel, M.E.; Lu, X. Environmental Stress-Induced Bacterial Lysis and Extracellular DNA Release Contribute to Campylobacter jejuni Biofilm Formation. Appl. Environ. Microbiol. 2018, 84, e02068-17. [Google Scholar] [CrossRef] [Green Version]
- Brown, H.L.; Hanman, K.; Reuter, M.; Betts, R.P.; Van Vliet, A.H.M. Campylobacter jejuni biofilms contain extracellular DNA and are sensitive to DNase I treatment. Front. Microbiol. 2015, 6, 699. [Google Scholar] [CrossRef] [Green Version]
- Gaasbeek, E.J.; Wagenaar, J.A.; Guilhabert, M.R.; van Putten, J.P.M.; Parker, C.T.; van der Wal, F.J. Nucleases Encoded by the Integrated Elements CJIE2 and CJIE4 Inhibit Natural Transformation of Campylobacter jejuni. J. Bacteriol. 2010, 192, 936–941. [Google Scholar] [CrossRef] [Green Version]
- Vorkapic, D.; Pressler, K.; Schild, S. Multifaceted roles of extracellular DNA in bacterial physiology. Curr. Genet. 2016, 62, 71–79. [Google Scholar] [CrossRef]
- De Vries, S.P.; Gupta, S.; Baig, A.; Wright, E.; Wedley, A.; Jensen, A.N.; Lora, L.L.; Humphrey, S.; Skovgård, H.; Macleod, K.; et al. Genome-wide fitness analyses of the foodborne pathogen Campylobacter jejuni in in vitro and in vivo models. Sci. Rep. 2017, 7, 1251. [Google Scholar] [CrossRef] [Green Version]
- Reuter, M.; Mallett, A.; Pearson, B.M.; van Vliet, A.H.M. Biofilm Formation by Campylobacter jejuni Is Increased under Aerobic Conditions. Appl. Environ. Microbiol. 2010, 76, 2122–2128. [Google Scholar] [CrossRef] [Green Version]
- Teh, A.H.T.; Lee, S.M.; Dykes, G.A. The influence of dissolved oxygen level and medium on biofilm formation by Campylobacter jejuni. Food Microbiol. 2017, 61, 120–125. [Google Scholar] [CrossRef]
- Teh, A.H.T.; Lee, S.M.; Dykes, G.A. The Influence of Prior Modes of Growth, Temperature, Medium, and Substrate Surface on Biofilm Formation by Antibiotic-Resistant Campylobacter jejuni. Curr. Microbiol. 2016, 73, 859–866. [Google Scholar] [CrossRef]
- Moore, J.E. Comparison of basal broth media for the optimal laboratory recovery of Campylobacter jejuni and Campylobacter coli. Ir. J. Med. Sci. 2000, 169, 187–189. [Google Scholar] [CrossRef]
- Kim, S.-H.; Park, C.; Lee, E.-J.; Bang, W.-S.; Kim, Y.-J.; Kim, J.-S. Biofilm formation of Campylobacter strains isolated from raw chickens and its reduction with DNase I treatment. Food Control 2017, 71, 94–100. [Google Scholar] [CrossRef]
- Dzianach, P.A.; Dykes, G.A.; Strachan, N.J.C.; Forbes, K.J.; Pérez-Reche, F.J. Challenges of biofilm control and utilization: Lessons from mathematical modelling. J. R. Soc. Interface 2019, 16, 20190042. [Google Scholar] [CrossRef] [Green Version]
- Zweifel, C.; Stephan, R. Microbial Decontamination of Poultry Carcasses. In Microbial Decontamination in the Food Industry: Novel Methods and Applications; Demirci, A., Ngadi, M.O., Eds.; Woodhead Publishing: Zurich, Switzerland, 2012; p. 804. [Google Scholar]
- Zhang, L.; Garner, L.J.; McKee, S.R.; Bilgili, S.F. Effectiveness of Several Antimicrobials Used in a Postchill Decontamination Tank against Salmonella and Campylobacter on Broiler Carcass Parts. J. Food Prot. 2018, 81, 1134–1141. [Google Scholar] [CrossRef]
- Kure, C.F.; Axelsson, L.; Carlehög, M.; Måge, I.; Jensen, M.R.; Holck, A. The effects of a pilot-scale steam decontamination system on the hygiene and sensory quality of chicken carcasses. Food Control 2020, 109, 106948. [Google Scholar] [CrossRef]
- EFSA Panel on Biological Hazards (BIOHAZ). Scientific Opinion on Campylobacter in broiler meat production: Control options and performance objectives and/or targets at different stages of the food chain. EFSA J. 2011, 9, 2105. [Google Scholar] [CrossRef]
- Yoon, K.S.; Burnette, C.N.; Oscar, T.P. Development of Predictive Models for the Survival of Campylobacter jejuni (ATCC 43051) on Cooked Chicken Breast Patties and in Broth as a Function of Temperature. J. Food Prot. 2004, 67, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Geeraerd, A.; Valdramidis, V.; Van Impe, J. GInaFiT, a freeware tool to assess non-log-linear microbial survivor curves. Int. J. Food Microbiol. 2005, 102, 95–105. [Google Scholar] [CrossRef] [PubMed]
- González, M.; Skandamis, P.N.; Hänninen, M.-L. A modified Weibull model for describing the survival of Campylobacter jejuni in minced chicken meat. Int. J. Food Microbiol. 2009, 136, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.H.; Kim, H.S.; Yoon, K.S. Survival and Risk Comparison of Campylobacter jejuni on Various Processed Meat Products. Int. J. Environ. Res. Public Health 2016, 13, 580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blankenship, L.C.; Craven, S.E. Campylobacter jejuni survival in chicken meat as a function of temperature. Appl. Environ. Microbiol. 1982, 44, 88–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, K.F.; Le Tran, H.; Kanenaka, R.Y.; Kathariou, S. Survival of clinical and poultry-derived isolates of Campylobacter jejuni at a low temperature (4 degrees C). Appl. Environ. Microbiol. 2001, 67, 4186–4191. [Google Scholar] [CrossRef] [Green Version]
- Garénaux, A.; Jugiau, F.; Rama, F.; de Jonge, R.; Denis, M.; Federighi, M.; Ritz, M. Survival of Campylobacter jejuni Strains from Different Origins under Oxidative Stress Conditions: Effect of Temperature. Curr. Microbiol. 2008, 56, 293–297. [Google Scholar] [CrossRef]
- Umaraw, P.; Prajapati, A.; Verma, A.K.; Pathak, V.; Singh, V.P. Control of Campylobacter in poultry industry from farm to poultry processing unit: A review. Crit. Rev. Food Sci. Nutr. 2017, 57, 659–665. [Google Scholar] [CrossRef]
- Valtierra-Rodríguez, D.; Heredia, N.L.; Garcia, S.; Sánchez, E. Reduction of Campylobacter jejuni and Campylobacter coli in Poultry Skin by Fruit Extracts. J. Food Prot. 2010, 73, 477–482. [Google Scholar] [CrossRef]
- Oberhardt, M.A.; Palsson, B.O.; Papin, J.A. Applications of genome-scale metabolic reconstructions. Mol. Syst. Biol. 2009, 5, 320. [Google Scholar] [CrossRef]
- Cole, J.A.; Kohler, L.; Hedhli, J.; Luthey-Schulten, Z. Spatially-resolved metabolic cooperativity within dense bacterial colonies. BMC Syst. Biol. 2015, 9, 15. [Google Scholar] [CrossRef] [Green Version]
- Phalak, P.; Chen, J.; Carlson, R.P.; Henson, M.A. Metabolic modeling of a chronic wound biofilm consortium predicts spatial partitioning of bacterial species. BMC Syst. Biol. 2016, 10, 90. [Google Scholar] [CrossRef] [Green Version]
- Tejera, N.; Crossman, L.; Pearson, B.; Stoakes, E.; Nasher, F.; Djeghout, B.; Poolman, M.; Wain, J.; Singh, D. Genome-Scale Metabolic Model Driven Design of a Defined Medium for Campylobacter jejuni M1cam. Front. Microbiol. 2020, 11, 1072. [Google Scholar] [CrossRef]
- Guccione, E.; del Rocio Leon-Kempis, M.; Pearson, B.M.; Hitchin, E.; Mulholland, F.; van Diemen, P.M.; Stevens, M.P.; Kelly, D.J. Amino acid-dependent growth of Campylobacter jejuni: Key roles for aspartase (AspA) under microaerobic and oxygen-limited conditions and identification of AspB (Cj0762), essential for growth on glutamate. Mol. Microbiol. 2008, 69, 77–93. [Google Scholar] [CrossRef]
- Muraoka, W.T.; Zhang, Q. Phenotypic and Genotypic Evidence for L-Fucose Utilization by Campylobacter jejuni. J. Bacteriol. 2011, 193, 1065–1075. [Google Scholar] [CrossRef] [Green Version]
- Snelling, W.J.; Matsuda, M.; Moore, J.E.; Dooley, J.S.G. Campylobacter jejuni. Lett. Appl. Microbiol. 2005, 41, 297–302. [Google Scholar] [CrossRef]
- Newell, D. Animal models of Campylobacter jejuni colonization and disease and the lessons to be learned from similar Helicobacter pylori models. J. Appl. Microbiol. 2001, 90, 57S–67S. [Google Scholar] [CrossRef]
- Black, R.E.; Perlman, D.; Clements, M.L.; Levine, M.M.; Blaser, M.J. Human Volunteer Studies with Campylobacter jejuni. In Campylobacter jejuni: Current Status and Future Trends; Nachamkin, I., Blaser, M.J., Tompkins, L., Eds.; ASM Press: Washington, DC, USA, 1993; pp. 207–215. [Google Scholar]
- Bacon, D.J.; Alm, R.A.; Burr, D.H.; Hu, L.; Kopecko, D.J.; Ewing, C.P.; Trust, T.J.; Guerry, P. Involvement of a Plasmid in Virulence of Campylobacter jejuni 81-176. Infect. Immun. 2000, 68, 4384–4390. [Google Scholar] [CrossRef] [Green Version]
- Bereswill, S.; Fischer, A.; Plickert, R.; Haag, L.M.; Otto, B.; Kühl, A.A.; Dasti, J.I.; Zautner, A.E.; Muñoz, M.; Loddenkemper, C.; et al. Novel Murine Infection Models Provide Deep Insights into the “Ménage à Trois” of Campylobacter jejuni, Microbiota and Host Innate Immunity. PLoS ONE 2011, 6, e20953. [Google Scholar] [CrossRef]
- Champion, O.L.; Wagley, S.; Titball, R.W. Galleria mellonella as a model host for microbiological and toxin research. Virulence 2016, 7, 840–845. [Google Scholar] [CrossRef]
- Tsai, C.J.-Y.; Loh, J.M.S.; Proft, T. Galleria mellonella infection models for the study of bacterial diseases and for antimicrobial drug testing. Virulence 2016, 7, 214–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senior, N.J.; Bagnall, M.C.; Champion, O.L.; Reynolds, S.; La Ragione, R.; Woodward, M.J.; Salguero, F.J.; Titball, R.W. Galleria mellonella as an infection model for Campylobacter jejuni virulence. J. Med. Microbiol. 2011, 60, 661–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humphrey, S.; Lacharme-Lora, L.; Chaloner, G.; Gibbs, K.; Humphrey, T.; Williams, N.; Wigley, P. Heterogeneity in the Infection Biology of Campylobacter jejuni Isolates in Three Infection Models Reveals an Invasive and Virulent Phenotype in a ST21 Isolate from Poultry. PLoS ONE 2015, 10, e0141182. [Google Scholar] [CrossRef] [PubMed]
- Teunis, P.F.; Nagelkerke, N.J.; Haas, C.N. Dose response models for infectious gastroenteritis. Risk Anal. 1999, 19, 1251–1260. [Google Scholar] [CrossRef] [PubMed]
- Teunis, P.F.; Kasuga, F.; Fazil, A.; Ogden, I.D.; Rotariu, O.; Strachan, N.J. Dose–response modeling of Salmonella using outbreak data. Int. J. Food Microbiol. 2010, 144, 243–249. [Google Scholar] [CrossRef]
- Strachan, N.; Doyle, M.; Kasuga, F.; Rotariu, O.; Ogden, I. Dose response modelling of Escherichia coli O157 incorporating data from foodborne and environmental outbreaks. Int. J. Food Microbiol. 2005, 103, 35–47. [Google Scholar] [CrossRef]
- Teunis, P.F.M.; Ogden, I.D.; Strachan, N.J.C. Hierarchical dose response of E. coli O157:H7 from human outbreaks incorporating heterogeneity in exposure. Epidemiol. Infect. 2008, 136, 761–770. [Google Scholar] [CrossRef]
- Xie, G.; Roiko, A.; Stratton, H.; Lemckert, C.; Dunn, P.K.; Mengersen, K. A Generalized QMRA Beta-Poisson Dose-Response Model. Risk Anal. 2016, 36, 1948–1958. [Google Scholar] [CrossRef]
- Langley, G.R.; Adcock, I.M.; Busquet, F.; Crofton, K.M.; Csernok, E.; Giese, C.; Heinonen, T.; Herrmann, K.; Hofmann-Apitius, M.; Landesmann, B.; et al. Towards a 21st-century roadmap for biomedical research and drug discovery: Consensus report and recommendations. Drug Discov. Today 2017, 22, 327–339. [Google Scholar] [CrossRef] [Green Version]
- Wagenaar, J.A.; French, N.P.; Havelaar, A.H. Preventing Campylobacter at the Source: Why Is It So Difficult? Clin. Infect. Dis. 2013, 57, 1600–1606. [Google Scholar] [CrossRef]
- Brauer, F. Mathematical epidemiology: Past, present, and future. Infect. Dis. Model. 2017, 2, 113–127. [Google Scholar] [CrossRef]
- Yébenes, J.C.; Ruiz-Rodriguez, J.C.; Ferrer, R.; Clèries, M.; Bosch, A.; Lorencio, C.; Rodriguez, A.; Nuvials, X.; Martin-Loeches, I.; Artigas, A.; et al. Epidemiology of sepsis in Catalonia: Analysis of incidence and outcomes in a European setting. Ann. Intensive Care 2017, 7, 19. [Google Scholar] [CrossRef] [Green Version]
- Kleinbaum, D.G.; Kupper, L.L.; Chambless, L.E. Logistic regression analysis of epidemiologic data: Theory and practice. Commun. Stat.-Theory Methods 1982, 11, 485–547. [Google Scholar] [CrossRef]
- Bavishi, C.; DuPont, H.L. Systematic review: The use of proton pump inhibitors and increased susceptibility to enteric infection. Aliment. Pharmacol. Ther. 2011, 34, 1269–1281. [Google Scholar] [CrossRef]
- Futagami, S.; Itoh, T.; Sakamoto, C. Systematic review with meta-analysis: Post-infectious functional dyspepsia. Aliment. Pharmacol. Ther. 2015, 41, 177–188. [Google Scholar] [CrossRef] [Green Version]
- Riddle, M.S.; Murray, J.A.; Cash, B.D.; Pimentel, M.; Porter, C.K. Pathogen-Specific Risk of Celiac Disease Following Bacterial Causes of Foodborne Illness: A Retrospective Cohort Study. Dig. Dis. Sci. 2013, 58, 3242–3245. [Google Scholar] [CrossRef]
- Kaakoush, N.O.; Castaño-Rodríguez, N.; Mitchell, H.M.; Man, S.M. Global Epidemiology of Campylobacter Infection. Clin. Microbiol. Rev. 2015, 28, 687–720. [Google Scholar] [CrossRef] [Green Version]
- Sears, A.; Baker, M.G.; Wilson, N.; Marshall, J.; Muellner, P.; Campbell, D.M.; Lake, R.J.; French, N. Marked Campylobacteriosis Decline after Interventions Aimed at Poultry, New Zealand. Emerg. Infect. Dis. 2011, 17, 1007–1015. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Reche, F.J.; Rotariu, O.; Lopes, B.S.; Forbes, K.J.; Strachan, N.J.C. Mining whole genome sequence data to efficiently attribute individuals to source populations. Sci. Rep. 2020, 10, 12124. [Google Scholar] [CrossRef]
- Falush, D. Bacterial genomics: Microbial GWAS coming of age. Nat. Microbiol. 2016, 1, 16059. [Google Scholar] [CrossRef]
- Yahara, K.; Méric, G.; Taylor, A.J.; de Vries, S.P.; Murray, S.; Pascoe, B.; Mageiros, L.; Torralbo, A.; Vidal, A.; Ridley, A.; et al. Genome-wide association of functional traits linked with Campylobacter jejuni survival from farm to fork. Environ. Microbiol. 2017, 19, 361–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ocejo, M.; Oporto, B.; Lavín, J.L.; Hurtado, A. Whole genome-based characterisation of antimicrobial resistance and genetic diversity in Campylobacter jejuni and Campylobacter coli from ruminants. Sci. Rep. 2021, 11, 8998. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Tyson, G.H.; Chen, Y.; Li, C.; Mukherjee, S.; Young, S.; Lam, C.; Folster, J.P.; Whichard, J.M.; McDermott, P.F. Whole-Genome Sequencing Analysis Accurately Predicts Antimicrobial Resistance Phenotypes in Campylobacter spp. Appl. Environ. Microbiol. 2016, 82, 459–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marotta, F.; Garofolo, G.; Di Marcantonio, L.; Di Serafino, G.; Neri, D.; Romantini, R.; Sacchini, L.; Alessiani, A.; Di Donato, G.; Nuvoloni, R.; et al. Antimicrobial resistance genotypes and phenotypes of Campylobacter jejuni isolated in Italy from humans, birds from wild and urban habitats, and poultry. PLoS ONE 2019, 14, e0223804. [Google Scholar] [CrossRef] [Green Version]
- Earle, S.G.; Wu, C.-H.; Charlesworth, J.; Stoesser, N.; Gordon, N.C.; Walker, T.M.; Spencer, C.C.A.; Iqbal, Z.; Clifton, D.A.; Hopkins, K.L.; et al. Identifying lineage effects when controlling for population structure improves power in bacterial association studies. Nat. Microbiol. 2016, 1, 16041. [Google Scholar] [CrossRef] [Green Version]
- Bender, R. Quantitative Risk Assessment in Epidemiological Studies Investigating Threshold Effects. Biom. J. 1999, 41, 305–319. [Google Scholar] [CrossRef]
- Duffy, L.L.; Blackall, P.J.; Cobbold, R.N.; Fegan, N. Quantitative effects of in-line operations on Campylobacter and Escherichia coli through two Australian broiler processing plants. Int. J. Food Microbiol. 2014, 188, 128–134. [Google Scholar] [CrossRef] [Green Version]
- Rawson, T.; Paton, R.S.; Colles, F.M.; Maiden, M.C.J.; Dawkins, M.S.; Bonsall, M.B. A Mathematical Modeling Approach to Uncover Factors Influencing the Spread of Campylobacter in a Flock of Broiler-Breeder Chickens. Front. Microbiol. 2020, 11, 576646. [Google Scholar] [CrossRef]
- Muller, C.J.; MacLehose, R.F. Estimating predicted probabilities from logistic regression: Different methods correspond to different target populations. Int. J. Epidemiol. 2014, 43, 962–970. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dzianach, P.A.; Pérez-Reche, F.J.; Strachan, N.J.C.; Forbes, K.J.; Dykes, G.A. The Use of Interdisciplinary Approaches to Understand the Biology of Campylobacter jejuni. Microorganisms 2022, 10, 2498. https://doi.org/10.3390/microorganisms10122498
Dzianach PA, Pérez-Reche FJ, Strachan NJC, Forbes KJ, Dykes GA. The Use of Interdisciplinary Approaches to Understand the Biology of Campylobacter jejuni. Microorganisms. 2022; 10(12):2498. https://doi.org/10.3390/microorganisms10122498
Chicago/Turabian StyleDzianach, Paulina A., Francisco J. Pérez-Reche, Norval J. C. Strachan, Ken J. Forbes, and Gary A. Dykes. 2022. "The Use of Interdisciplinary Approaches to Understand the Biology of Campylobacter jejuni" Microorganisms 10, no. 12: 2498. https://doi.org/10.3390/microorganisms10122498