
Changes in Phylogenetic and Functional Diversity of Ciliates along the Course of a Mediterranean Karstic River
 , , , , , , , and
, , , , , , , and 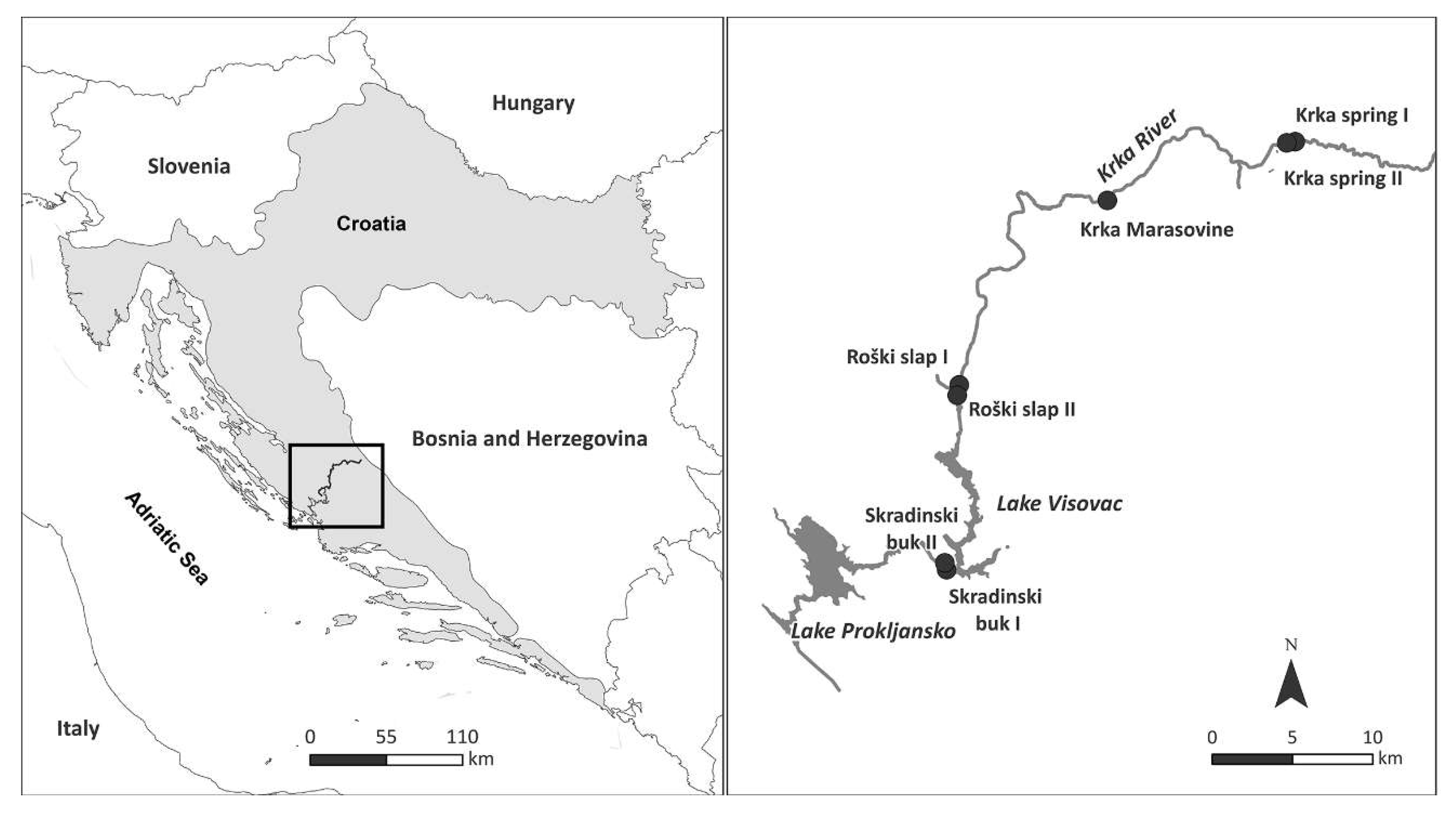{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling Procedure
2.3. DNA Extraction and PCR Reaction
2.4. Bioinformatics and Statistics
3. Results
3.1. Sequencing
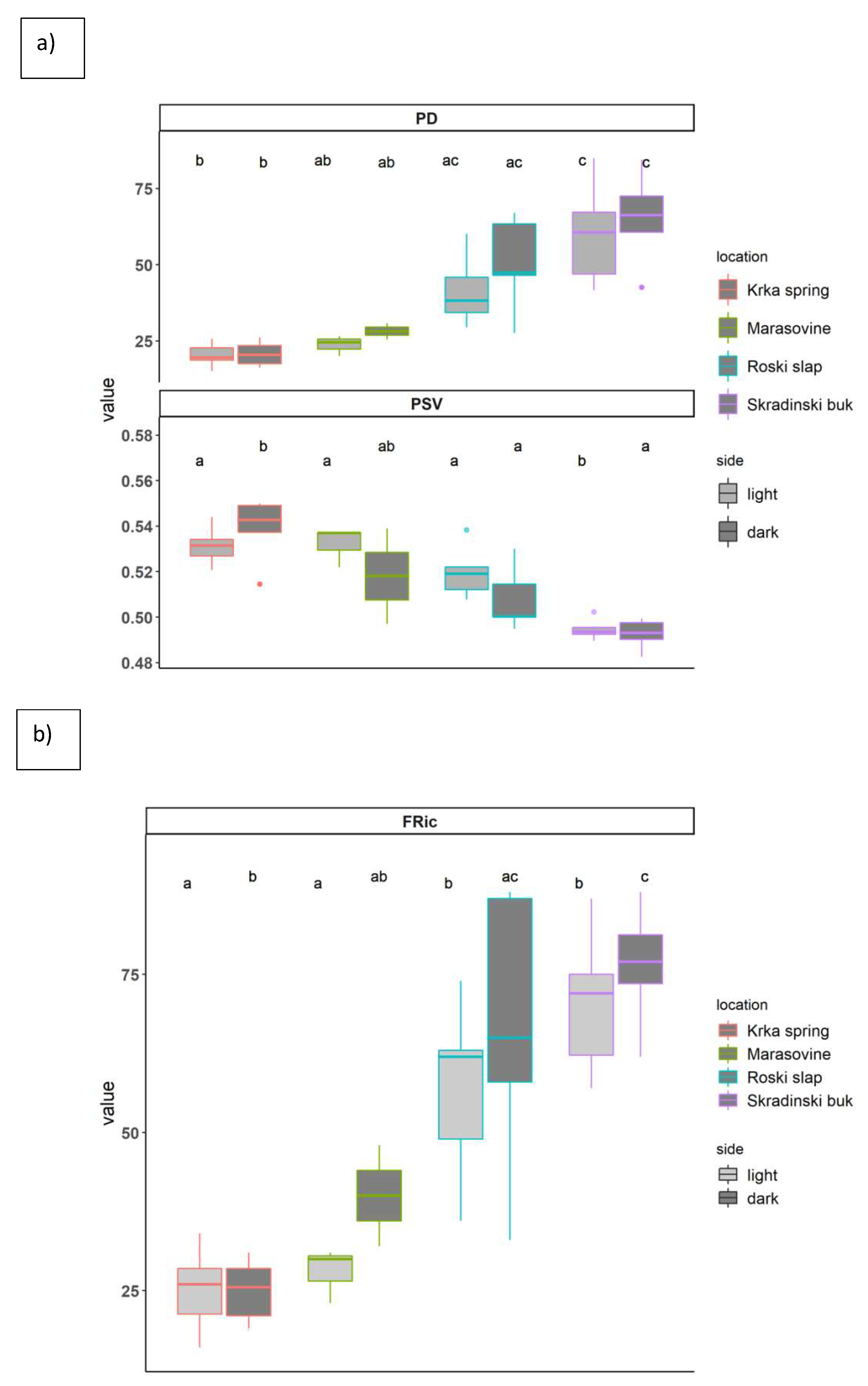
3.2. Ciliate Phylogenetic and Functional Diversity Metrics
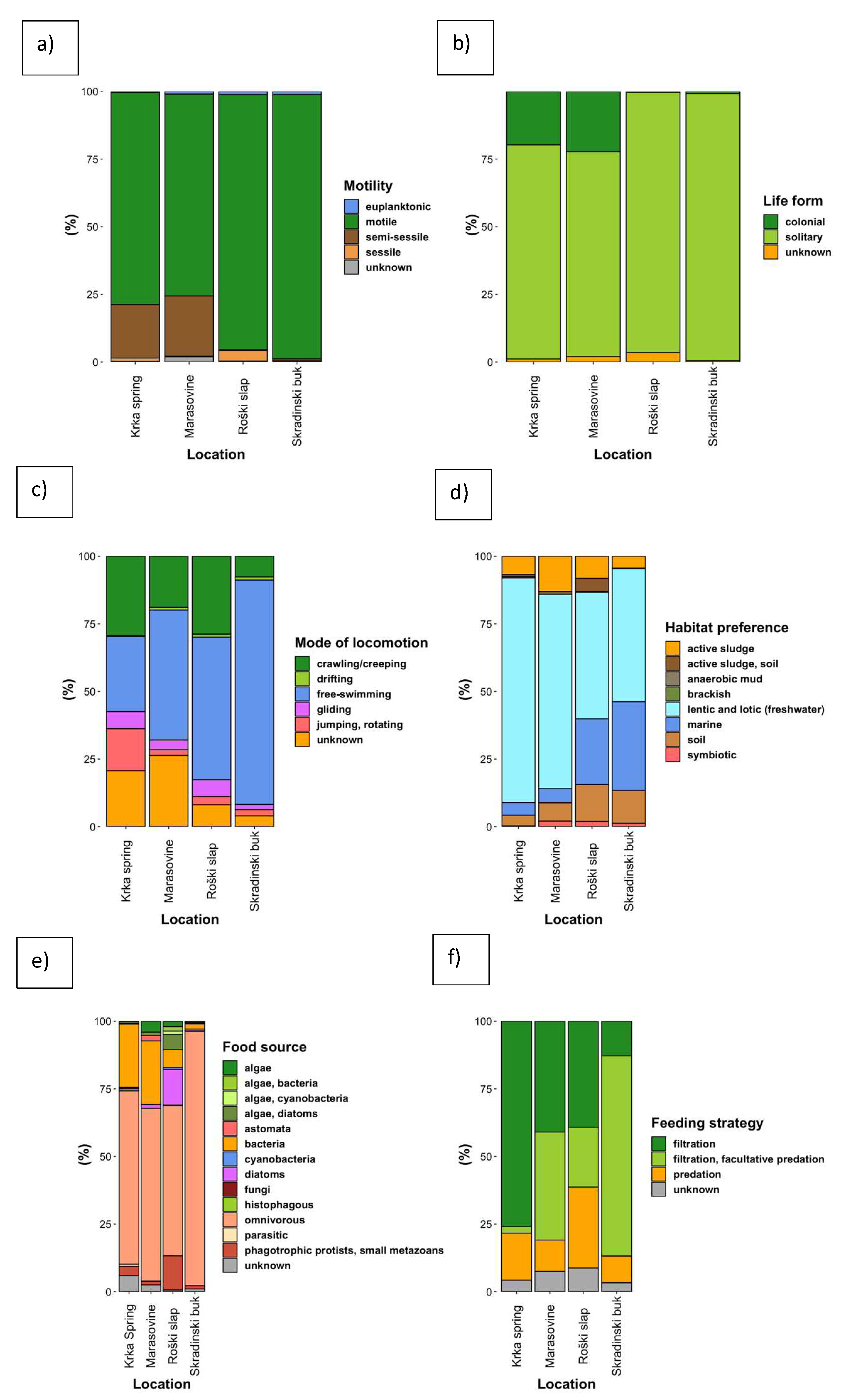
3.3. Community Composition with Regard to Ciliate Functional Groups
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singer, D.; Seppey, C.V.; Lentendu, G.; Dunthorn, M.; Bass, D.; Belbahri, L.; Blandenier, Q.; Debroas, D.; de Groot, G.A.; de Vargas, C.; et al. Protist taxonomic and functional diversity in soil, freshwater and marine ecosystems. Environ. Int. 2020, 146, 106262. [Google Scholar] [CrossRef]
- Weitere, M.; Erken, M.; Majdi, N.; Arndt, H.; Norf, H.; Reinshagen, M.; Traunspurger, W.; Walterscheid, A.; Wey, J.K. The food web perspective on aquatic biofilms. Ecol. Monogr. 2018, 88, 543–559. [Google Scholar] [CrossRef]
- Foissner, W.; Berger, H. A user-friendly guide to the ciliates (Protozoa, Ciliophora) commonly used by hydrobiologists as bioindicators in rivers, lakes, and waste waters, with notes on their ecology. Freshw. Biol. 1996, 35, 375–482. [Google Scholar] [CrossRef]
- Hughes, J. Freshwater Ecology And Conservation: Approaches And Techniques; Freshwater Ecology and Conservation; Oxford University Press: Oxford, UK, 2018. [Google Scholar]
- Kulaš, A.; Gulin, V.; Kepčija, R.M.; Žutinić, P.; Perić, M.S.; Orlić, S.; Kajan, K.; Stoeck, T.; Lentendu, G.; Čanjevac, I.; et al. Ciliates (Alveolata, Ciliophora) as bioindicators of environmental pressure: A karstic river case. Ecol. Indic. 2021, 124, 107430. [Google Scholar] [CrossRef]
- Gulin, V.; Kepčija, R.M.; Perić, M.S.; Felja, I.; Fajković, H.; Križnjak, K. Environmental and periphyton response to stream revitalization–A pilot study from a tufa barrier. Ecol. Indic. 2021, 126, 107629. [Google Scholar] [CrossRef]
- Gulin, V.; Vlaičević, B.; Perić, M.S.; Rebrina, F.; Kepčija, R.M. Taxonomic and Functional Metrics of Ciliates and Amoeboid Protists in Response to Stream Revitalization. Front. Microbiol. 2022, 13, 842395. [Google Scholar] [CrossRef] [PubMed]
- Lentendu, G.; Buosi, P.R.B.; Cabral, A.F.; Segóvia, B.T.; Meira, B.R.; Lansac-Tôha, F.M.; Velho, L.F.M.; Ritter, C.D.; Dunthorn, M. Protist Biodiversity and Biogeography in Lakes From Four Brazilian River–Floodplain Systems. J. Eukaryot. Microbiol. 2018, 66, 592–599. [Google Scholar] [CrossRef]
- Singer, D.; Kosakyan, A.; Seppey, C.V.W.; Pillonel, A.; Fernández, L.D.; Fontaneto, D.; Mitchell, E.; Lara, E. Environmental filtering and phylogenetic clustering correlate with the distribution patterns of cryptic protist species. Ecology 2018, 99, 904–914. [Google Scholar] [CrossRef] [Green Version]
- Lentendu, G.; Dunthorn, M. Phylogenetic relatedness drives protist assembly in marine and terrestrial environments. Glob. Ecol. Biogeogr. 2021, 30, 1532–1544. [Google Scholar] [CrossRef]
- Cornwell, W.K.; Schwilk, L.D.W.; Ackerly, D. A Trait-Based Test For Habitat Filtering: Convex Hull Volume. Ecology 2006, 87, 1465–1471. [Google Scholar] [CrossRef]
- Lebrija-Trejos, E.; Pérez-García, E.A.; Meave, J.A.; Bongers, F.; Poorter, L. Functional traits and environmental filtering drive community assembly in a species-rich tropical system. Ecology 2010, 91, 386–398. [Google Scholar] [CrossRef] [Green Version]
- Levine, J. A trail map for trait-based studies. Nature 2015, 529, 163–164. [Google Scholar] [CrossRef] [PubMed]
- Asefa, M.; Cao, M.; Zhang, G.; Ci, X.; Li, J.; Yang, J. Environmental filtering structures tree functional traits combination and lineages across space in tropical tree assemblages. Sci. Rep. 2017, 7, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boet, O.; Arnan, X.; Retana, J. The role of environmental vs. biotic filtering in the structure of European ant communities: A matter of trait type and spatial scale. PLoS ONE 2020, 15, e0228625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerhold, P.; Cahill, J.F., Jr.; Winter, M.; Bartish, I.V.; Prinzing, A. Phylogenetic patterns are not proxies of community assembly mechanisms (they are far better). Funct. Ecol. 2015, 29, 600–614. [Google Scholar] [CrossRef]
- Cadotte, M.W.; Cavender-Bares, J.; Tilman, D.; Oakley, T. Using Phylogenetic, Functional and Trait Diversity to Understand Patterns of Plant Community Productivity. PLoS ONE 2009, 4, e5695. [Google Scholar] [CrossRef] [PubMed]
- Flynn, D.F.B.; Mirotchnick, N.; Jain, M.; Palmer, M.I.; Naeem, S. Functional and phylogenetic diversity as predictors of biodiversity–ecosystem-function relationships. Ecology 2011, 92, 1573–1581. [Google Scholar] [CrossRef] [Green Version]
- Faith, D.P. Conservation evaluation and phylogenetic diversity. Biol. Conserv. 1992, 61, 1–10. [Google Scholar] [CrossRef]
- Srivastava, D.S.; Cadotte, M.W.; MacDonald, A.A.M.; Marushia, R.G.; Mirotchnick, N. Phylogenetic diversity and the functioning of ecosystems. Ecol. Lett. 2012, 15, 637–648. [Google Scholar] [CrossRef]
- Ritchie, A.M.; Hua, X.; Cardillo, M.; Yaxley, K.J.; Dinnage, R.; Bromham, L. Phylogenetic diversity metrics from molecular phylogenies: Modelling expected degree of error under realistic rate variation. Divers. Distrib. 2020, 27, 164–178. [Google Scholar] [CrossRef]
- Helmus, M.R.; Bland, T.J.; Williams, C.K.; Ives, A.R. Phylogenetic Measures of Biodiversity. Am. Nat. 2007, 169, E68–E83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozupone, C.; Knight, R. UniFrac: A New Phylogenetic Method for Comparing Microbial Communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozupone, C.; Hamady, M.; Knight, R. UniFrac–An online tool for comparing microbial community diversity in a phylogenetic context. BMC Bioinform. 2006, 7, 371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, C.H.; Fine, P.V.A. Phylogenetic beta diversity: Linking ecological and evolutionary processes across space in time. Ecol. Lett. 2008, 11, 1265–1277. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.; Lladser, M.E.; Knights, D.; Stombaugh, J.; Knight, R. UniFrac: An effective distance metric for microbial community comparison. ISME J. 2011, 5, 169–172. [Google Scholar] [CrossRef] [Green Version]
- Chang, Q.; Luan, Y.; Sun, F. Variance adjusted weighted UniFrac: A powerful beta diversity measure for comparing communities based on phylogeny. BMC Bioinform. 2011, 12, 118. [Google Scholar] [CrossRef] [Green Version]
- De Vrieze, J.; Christiaens, M.E.; Walraedt, D.; Devooght, A.; Ijaz, U.Z.; Boon, N. Microbial community redundancy in anaerobic digestion drives process recovery after salinity exposure. Water Res. 2017, 111, 109–117. [Google Scholar] [CrossRef]
- Nock, C.A.; Vogt, R.J.; Beisner, B.E. Functional traits. In eLS; John Wiley & Sons Ltd.: Chichester, UK, 2016. [Google Scholar] [CrossRef]
- Walker, B. Conserving Biological Diversity through Ecosystem Resilience. Conserv. Biol. 1995, 9, 747–752. [Google Scholar] [CrossRef]
- Mason, N.W.H.; Mouillot, D.; Lee, W.G.; Wilson, J.B. Functional richness, functional evenness and functional divergence: The primary components of functional diversity. Oikos 2005, 111, 112–118. [Google Scholar] [CrossRef]
- Mouchet, M.A.; Villéger, S.; Mason, N.W.H.; Mouillot, D. Functional diversity measures: An overview of their redundancy and their ability to discriminate community assembly rules. Funct. Ecol. 2010, 24, 867–876. [Google Scholar] [CrossRef]
- Weisse, T. Functional diversity of aquatic ciliates. Eur. J. Protistol. 2017, 61, 331–358. [Google Scholar] [CrossRef]
- Van Wichelen, J.; Johansson, L.S.; Vanormelingen, P.; Declerck, S.A.; Lauridsen, T.L.; De Meester, L.; Jeppesen, E.; Vyverman, W. Planktonic ciliate community structure in shallow lakes of lowland Western Europe. Eur. J. Protistol. 2013, 49, 538–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foissner, W.; Agatha, S.; Berger, H. Soil Ciliates (Protozoa, Ciliophora) from Namibia (Southwest Africa), with Emphasis on Two Contrasting Environments, the Etosha Region and the Namib Desert; Part I: Text and Line Drawings; Biologiezentrum des Oberösterreichisches Landesmuseum: Linz, Austria, 2002. [Google Scholar]
- Adl, S.M.; Bass, D.; Lane, C.E.; Lukeš, J.; Schoch, C.L.; Smirnov, A.; Agatha, S.; Berney, C.; Brown, M.W.; Burki, F.; et al. Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes. J. Eukaryot. Microbiol. 2018, 66, 4–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlaičević, B.; Kepčija, R.M.; Gulin, V.; Čakalić, I.T.; Kepec, M.; Čerba, D. Key drivers influencing the colonization of periphytic ciliates and their functional role in hydrologically dynamic floodplain lake ecosystem. Knowl. Manag. Aquat. Ecosyst. 2021, 422, 33. [Google Scholar] [CrossRef]
- Vlaičević, B.; Gulin, V.; Kepčija, R.M.; Čakalić, I.T. Periphytic Ciliate Communities in Lake Ecosystem of Temperate Riverine Floodplain: Variability in Taxonomic and Functional Composition and Diversity with Seasons and Hydrological Changes. Water 2022, 14, 551. [Google Scholar] [CrossRef]
- Ford, T.; Pedley, H. A review of tufa and travertine deposits of the world. Earth-Sci. Rev. 1996, 41, 117–175. [Google Scholar] [CrossRef]
- Primc-Habdija, B.; Matoničkin, R. A new freshwater folliculinid (Ciliophora) from the karstic region of Croatia. Eur. J. Protistol. 2005, 41, 37–43. [Google Scholar] [CrossRef]
- Bonacci, O. Hydrological investigations of Dinaric karst at the Krčić catchment and the river Krka springs (Yugoslavia). J. Hydrol. 1985, 82, 317–326. [Google Scholar] [CrossRef]
- Bonacci, O.; Ljubenkov, I.; Roje-Bonacci, T. Karst flash floods: An example from the Dinaric karst (Croatia). Nat. Hazards Earth Syst. Sci. 2006, 6, 195–203. [Google Scholar] [CrossRef]
- Perica, D.; Orešić, D.; Trajbar, S. Geomorfološka obilježja doline i poriječja rijeke Krke s osvrtom na dio od Knina do Bilušića buka. Geoadria 2005, 10, 131–156. [Google Scholar] [CrossRef] [Green Version]
- Amaral-Zettler, L.A.; McCliment, E.A.; Ducklow, H.W.; Huse, S.M. A Method for Studying Protistan Diversity Using Massively Parallel Sequencing of V9 Hypervariable Regions of Small-Subunit Ribosomal RNA Genes. PLoS ONE 2009, 4, e6372. [Google Scholar] [CrossRef]
- Stock, A.; Jürgens, K.; Bunge, J.; Stoeck, T. Protistan diversity in suboxic and anoxic waters of the Gotland Deep (Baltic Sea) as revealed by 18S rRNA clone libraries. Aquat. Microb. Ecol. 2009, 55, 267–284. [Google Scholar] [CrossRef]
- Stoeck, T.; Bass, D.; Nebel, M.; Christen, R.; Jones, M.D.M.; Breiner, H.-W.; Richards, T.A. Multiple marker parallel tag environmental DNA sequencing reveals a highly complex eukaryotic community in marine anoxic water. Mol. Ecol. 2010, 19, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Mahé, F.; Rognes, T.; Quince, C.; de Vargas, C.; Dunthorn, M. Swarm v2: Highly-scalable and high-resolution amplicon clustering. PeerJ 2015, 3, e1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillou, L.; Bachar, D.; Audic, S.; Bass, D.; Berney, C.; Bittner, L.; Boutte, C.; Burgaud, G.; de Vargas, C.; Decelle, J.; et al. The Protist Ribosomal Reference database (PR2): A catalog of unicellular eukaryote Small Sub-Unit rRNA sequences with curated taxonomy. Nucleic Acids Res. Oxf. Acad. 2013, 41, D597–D604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger, S.A.; Krompass, D.; Stamatakis, A. Performance, Accuracy, and Web Server for Evolutionary Placement of Short Sequence Reads under Maximum Likelihood. Syst. Biol. 2011, 60, 291–302. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chernomor, O.; Von Haeseler, A.; Minh, B.Q. Terrace Aware Data Structure for Phylogenomic Inference from Supermatrices. Syst. Biol. 2016, 65, 997–1008. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. Available online: https://www.R-project.org/ (accessed on 29 June 2021).
- Gloor, G.B.; Macklaim, J.M.; Pawlowsky-Glahn, V.; Egozcue, J.J. Microbiome Datasets Are Compositional: And This Is Not Optional. Front. Microbiol. 2017, 8, 2224. [Google Scholar] [CrossRef] [Green Version]
- Faith, D.P.; Baker, A.M. Phylogenetic Diversity (PD) and Biodiversity Conservation: Some Bioinformatics Challenges. Evol. Bioinform. 2006, 2, 121–128. [Google Scholar] [CrossRef]
- Kembel, S.W.; Cowan, P.D.; Helmus, M.R.; Cornwell, W.K.; Morlon, H.; Ackerly, D.D.; Blomberg, S.P.; Webb, C.O. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.; Wagner, H. Vegan: Community Ecology Package. R Package Version 2.4-6. 2018. Available online: https://CRAN.R-project.org/package=vegan (accessed on 27 April 2018).
- Tucker, C.M.; Davies, T.J.; Cadotte, M.W.; Pearse, W.D. On the relationship between phylogenetic diversity and trait diversity. Ecology 2018, 99, 1473–1479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tucker, C.M.; Cadotte, M.W.; Carvalho, S.B.; Jonathan Davies, T.; Ferrier, S.; Fritz, S.A.; Grenyer, R.; Helmus, M.R.; Jin, L.S.; Mooers, A.O.; et al. A guide to phylogenetic metrics for conservation, community ecology and macroecology. Biol. Rev. 2017, 92, 698–715. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P.; Shipley, B. FD: Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology R Package Version 1.0-12. 2014. Available online: https://cran.r-project.org/web/packages/FD (accessed on 18 October 2022).
- Slowikowski, K.; Schep, A.; Hughes, S.; Dang, K.T.; Lukauskas, S.; Irisson, J.O.; Kamvar, Z.N.; Ryan, T.; Christophe, D.; Hiroaki, Y. Automatically Position Non-overlapping Text Labels with “ggplot2”. 2021. Available online: https://github.com/slowkow/ggrepel (accessed on 18 October 2022).
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.A.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Limberger, R.; Wickham, S.A. Competition–colonization trade-offs in a ciliate model community. Oecologia 2011, 167, 723–732. [Google Scholar] [CrossRef] [Green Version]
- Jackson, K.M.; Berger, J. Survival of ciliate protozoa under starvation conditions and at low bacterial levels. Microb. Ecol. 1984, 10, 47–59. [Google Scholar] [CrossRef]
- Kusuoka, Y.; Watanabe, Y. Growth and survival of peritrich ciliates in an urban stream. Oecologia 1987, 73, 16–20. [Google Scholar] [CrossRef]
- Esteban, G.F.; Fenchel, T.M. Ecology of Protozoa: The Biology of Free-living Phagotrophic Protists, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2021. [Google Scholar]
- Biggs, B.J.F. Eutrophication of streams and rivers: Dissolvednutrient–chlorophyll relationships for benthic algae. J. N. Amer. Benthol. Soc. 2000, 19, 17–31. [Google Scholar] [CrossRef] [Green Version]
- Biggs, B.J.F.; Close, M.E. Periphyton biomass dynamics in gravel bedrivers: The relative effects of flows and nutrients. Freshw. Biol. 1989, 22, 209–231. [Google Scholar] [CrossRef]
- Besemer, K. Biodiversity, community structure and function of biofilms in stream ecosystems. Res. Microbiol. 2015, 166, 774–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biondić, B.; Biondić, R. Hidrogeologija Dinarskog Krša u Hrvatskoj; Sveučilište u Zagrebu: Geotehnički Fakultet, Varaždin, 2014; pp. 197–203. [Google Scholar]
- Prelovšek, M.; Hajna, N.Z. (Eds.) Pressures and Protection of the Underground Karst–Cases from Slovenia and Croatia; Karst Research Institute ZRC SAZU: Postojna, Slovenia, 2011. [Google Scholar]
- Smith, S.V.; Swaney, D.P.; Buddemeier, R.W.; Scarsbrook, M.R.; Weatherhead, M.A.; Humborg, C.; Eriksson, H.; Hannerz, F. River Nutrient Loads and Catchment Size. Biogeochemistry 2005, 75, 83–107. [Google Scholar] [CrossRef]
- WHO. Manganese in Drinking-water. Background Document for Development of WHO Guidelines for Drinking-water Quality; World Health Organization: Geneva, Switzerland, 2011.
- Baković, N.; Kepčija, R.M.; Siemensma, F.J. Transitional and small aquatic cave habitats diversification based on protist assemblages in the Veternica cave (Medvednica Mt., Croatia). Subterr. Biol. 2022, 42, 43–60. [Google Scholar] [CrossRef]
- Kulaš, A.; Udovič, M.G.; Tapolczai, K.; Žutinić, P.; Orlić, S.; Levkov, Z. Diatom eDNA metabarcoding and morphological methods for bioassessment of karstic river. Sci. Total Environ. 2022, 829, 154536. [Google Scholar] [CrossRef] [PubMed]
- Snyder, R.E.; Chesson, P. How the Spatial Scales of Dispersal, Competition, and Environmental Heterogeneity Interact to Affect Coexistence. Am. Nat. 2004, 164, 633–650. [Google Scholar] [CrossRef]
- Bonacci, O.; Andrić, I.; Roje-Bonacci, T. Hydrological analysis of Skradinski Buk tufa waterfall (Krka River, Dinaric karst, Croatia). Environ. Earth Sci. 2017, 76, 669. [Google Scholar] [CrossRef]
- Gibert, J.; Danielopol, D.; Stanford, J.A. Groundwater Ecology; Academic Press: San Diego, CA, USA, 1994. [Google Scholar]
- Medinger, R.; Nolte, V.; Pandey, R.V.; Jost, S.; Ottenwälder, B.; Schlötterer, C.; Boenigk, J. Diversity in a hidden world: Potential and limitation of next-generation sequencing for surveys of molecular diversity of eukaryotic microorganisms. Mol. Ecol. 2010, 19, 32–40. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gulin Beljak, V.; Kulaš, A.; Lentendu, G.; Vlaičević, B.; Gligora Udovič, M.; Sertić Perić, M.; Rebrina, F.; Žutinić, P.; Orlić, S.; Matoničkin Kepčija, R. Changes in Phylogenetic and Functional Diversity of Ciliates along the Course of a Mediterranean Karstic River. Microorganisms 2022, 10, 2493. https://doi.org/10.3390/microorganisms10122493
Gulin Beljak V, Kulaš A, Lentendu G, Vlaičević B, Gligora Udovič M, Sertić Perić M, Rebrina F, Žutinić P, Orlić S, Matoničkin Kepčija R. Changes in Phylogenetic and Functional Diversity of Ciliates along the Course of a Mediterranean Karstic River. Microorganisms. 2022; 10(12):2493. https://doi.org/10.3390/microorganisms10122493
Chicago/Turabian StyleGulin Beljak, Vesna, Antonija Kulaš, Guillaume Lentendu, Barbara Vlaičević, Marija Gligora Udovič, Mirela Sertić Perić, Fran Rebrina, Petar Žutinić, Sandi Orlić, and Renata Matoničkin Kepčija. 2022. "Changes in Phylogenetic and Functional Diversity of Ciliates along the Course of a Mediterranean Karstic River" Microorganisms 10, no. 12: 2493. https://doi.org/10.3390/microorganisms10122493