
Weapons against Themselves: Identification and Use of Quorum Sensing Volatile Molecules to Control Plant Pathogenic Fungi Growth

 , and
, and
Abstract
:1. Introduction
- -
- To determine the molecules constituting the volatolome of F. culmorum and C. sativus during culture conditions involving low and high spore concentrations.
- -
- To determine whether the volatolome of each fungus in overpopulation conditions has an impact on the mycelial growth of the same species in low spore culture conditions.
- -
- To identify in these volatolomes potential QSM as a first step in the development of a plant fungal pathogen control method using VOCs produced by the targeted species.
2. Materials and Methods
2.1. Biological Material
2.2. Collection and Identification of Fungal Volatile Organic Compounds (VOCs)
2.2.1. Sample Preparation
2.2.2. Extraction of VOCs Emitted by Both Fungi in Normal and Overpopulation Conditions
2.2.3. Analysis of VOCs Emitted by Both Fungi in Normal and Overpopulation Conditions
2.2.4. Identification of VOCs Emitted by Both Fungi in Normal and Overpopulation Conditions
2.3. Characterization of the Effect of Fungal Volatolomes on Fungal Growth
2.4. Characterization of Individual VOCs’ Effects on Fungal Growth
2.5. Statistical Analysis
3. Results
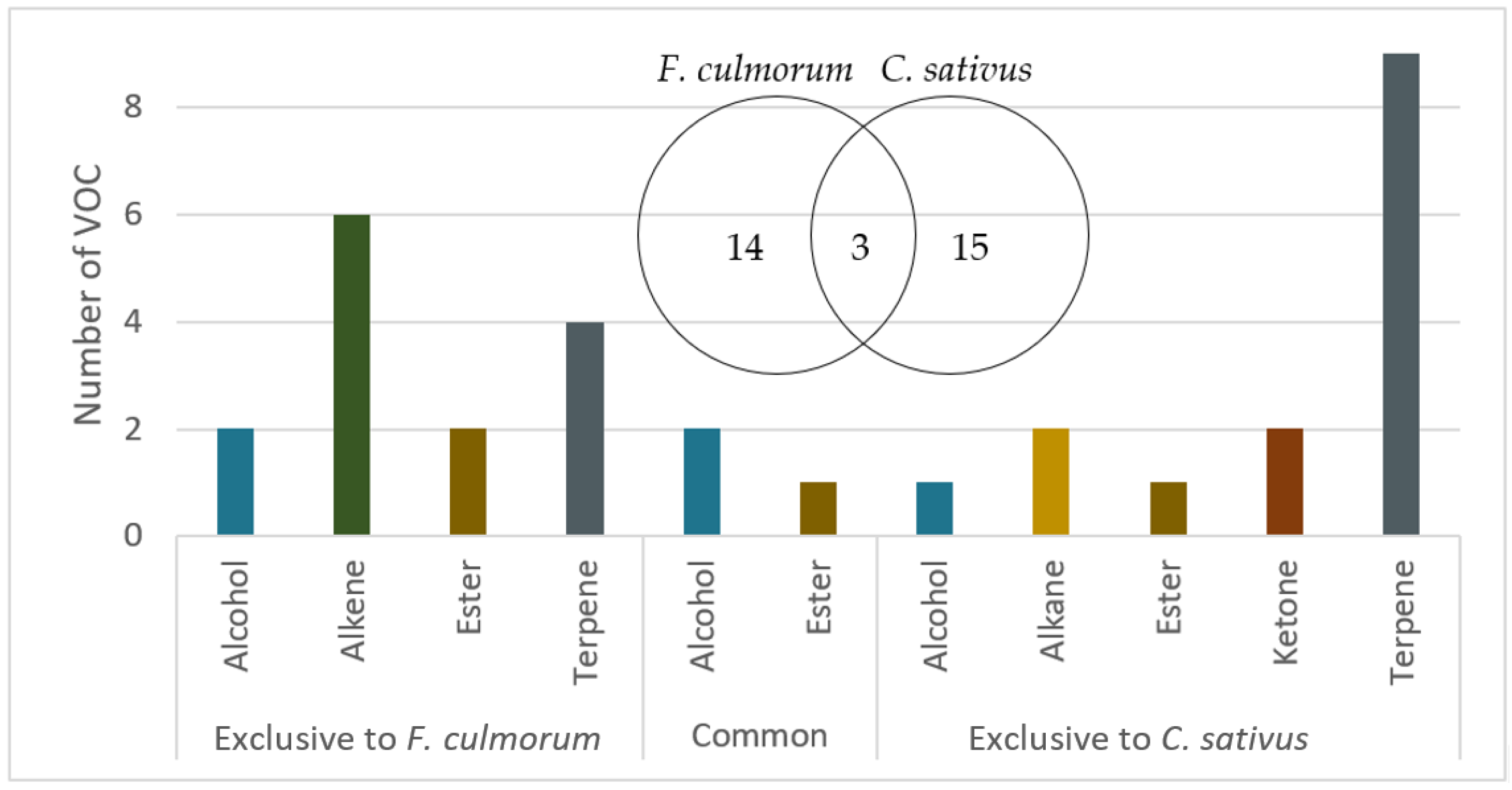
3.1. Collection and Identification of Fungal Volatile Organic Compounds
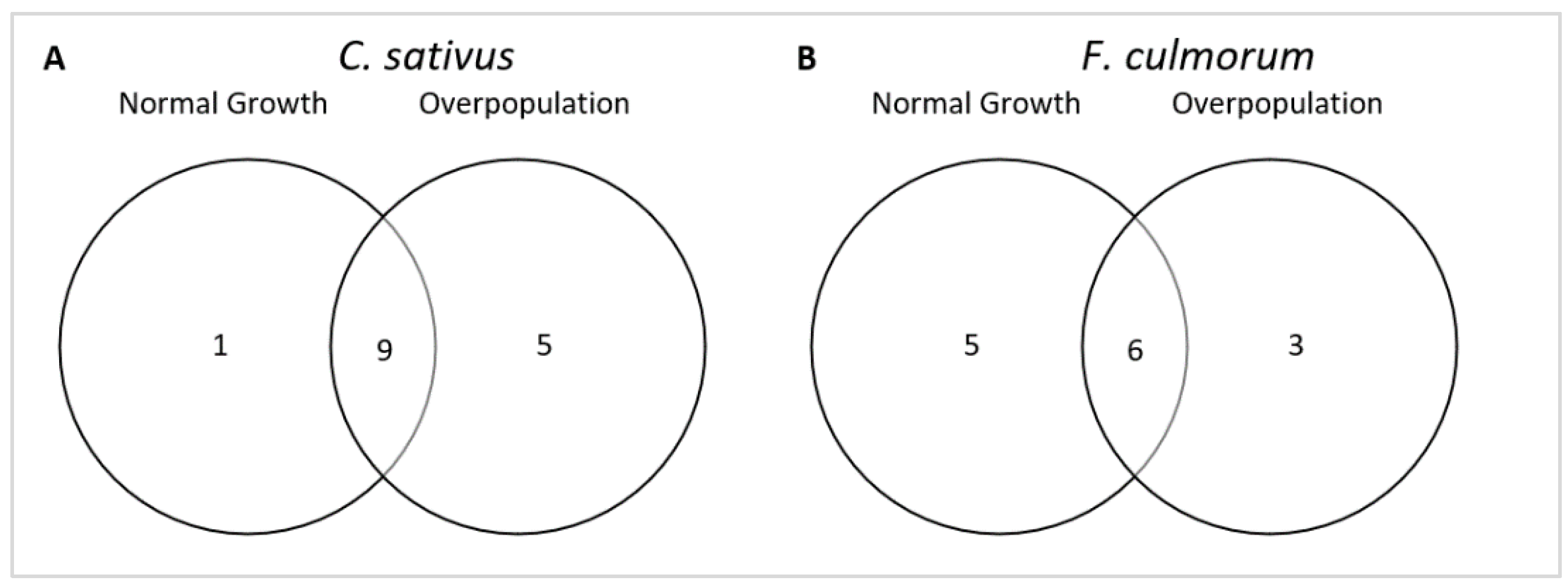
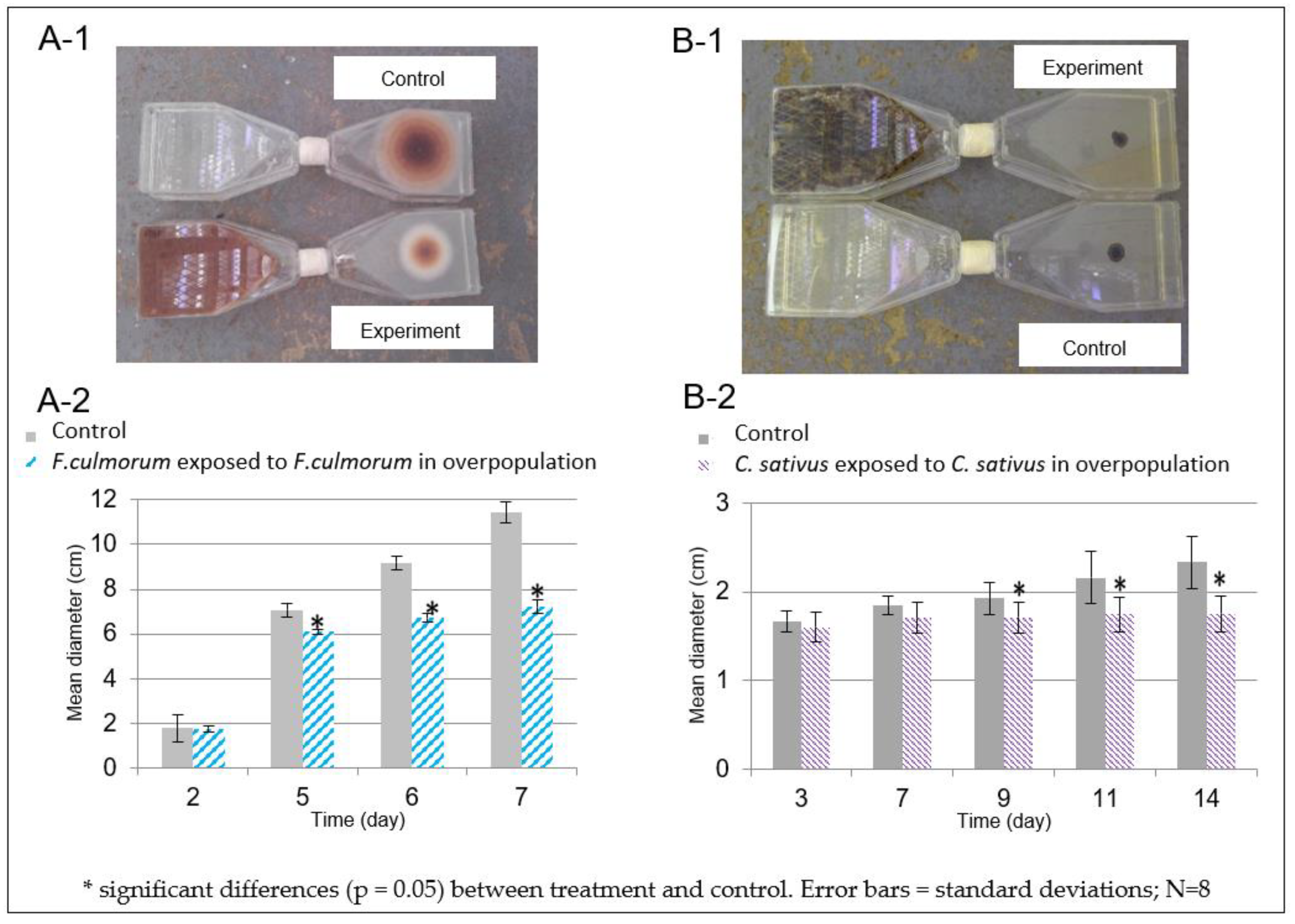
3.2. Characterization of the Fungal Volatolome Effect on Fungal Growth
3.3. Characterization of VOCs’ Individual Effects on Fungal Growth
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Barriuso, J.; Hogan, D.A.; Keshavarz, T.; Martínez, M.J. Role of quorum sensing and chemical communication in fungal biotechnology and pathogenesis. FEMS Microbiol. Rev. 2018, 42, 627–638. [Google Scholar] [CrossRef]
- Miller, M.B.; Bassler, B.L. Quorum sensing in bacteria. Annu. Rev. Microbiol. 2001, 55, 165–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barriuso, J. Quorum sensing mechanisms in fungi. AIMS Microbiol. 2015, 1, 37–47. [Google Scholar] [CrossRef]
- Williams, P. Quorum sensing, communication and cross-kingdom signalling in the bacterial world. Microbiology 2007, 153, 3923–3938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albuquerque, P.; Casadevall, A. Quorum sensing in fungi a review. Med. Mycol. 2012, 50, 337–345. [Google Scholar] [CrossRef] [Green Version]
- Hornby, J.M.; Jensen, E.C.; Lisec, A.D.; Tasto, J.J.; Jahnke, B.; Shoemaker, R.; Dussault, P.; Nickerson, K.W. Quorum Sensing in the Dimorphic Fungus Candida albicans Is Mediated by Farnesol. Appl. Environ. Microbiol. 2001, 67, 2982–2992. [Google Scholar] [CrossRef] [Green Version]
- Lingappa, B.T.; Prasad, M.; Lingappa, Y.; Hunt, D.F.; Biemann, K. Phenethyl alcohol and tryptophol: Autoantibiotics produced by the fungus Candida albicans. Science 1969, 163, 192–194. [Google Scholar] [CrossRef]
- Kügler, S.; Sebghati, T.S.; Eissenberg, L.G.; Goldman, W.E. Phenotypic variation and intracellular parasitism by Histoplasma capsulatum. Proc. Natl. Acad. Sci. USA 2000, 97, 8794–8798. [Google Scholar] [CrossRef] [Green Version]
- Hornby, J.M.; Jacobitz-Kizzier, S.M.; McNeel, D.J.; Jensen, E.C.; Treves, D.S.; Nickerson, K.W. Inoculum Size Effect in Dimorphic Fungi: Extracellular Control of Yeast-Mycelium Dimorphism in Ceratocystis ulmi. Appl. Environ. Microbiol. 2004, 70, 1356–1359. [Google Scholar] [CrossRef] [Green Version]
- Roca, M.G.; Arlt, J.; Jeffree, C.E.; Read, N.D. Cell biology of conidial anastomosis tubes in Neurospora crassa. Eukaryot. Cell 2005, 4, 911–919. [Google Scholar] [CrossRef]
- Severin, F.F.; Meer, M.V.; Smirnova, E.A.; Knorre, D.A.; Skulachev, V.P. Natural causes of programmed death of yeast Saccharomyces cerevisiae. Biochim. Biophys. Acta-Mol. Cell Res. 2008, 1783, 1350–1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.; Chang, Y.C.; Nardone, G.; Kwon-Chung, K.J. TUP1 disruption in Cryptococcus neoformans uncovers a peptide-mediated density-dependent growth phenomenon that mimics quorum sensing. Mol. Microbiol. 2007, 64, 591–601. [Google Scholar] [CrossRef]
- Berrocal, A.; Oviedo, C.; Nickerson, K.W.; Navarrete, J. Quorum sensing activity and control of yeast-mycelium dimorphism in Ophiostoma floccosum. Biotechnol. Lett. 2014, 36, 1503–1513. [Google Scholar] [CrossRef] [PubMed]
- Williams, H.E.; Steele, J.C.P.; Clements, M.O.; Keshavarz, T. γ-Heptalactone is an endogenously produced quorum-sensing molecule regulating growth and secondary metabolite production by Aspergillus nidulans. Appl. Microbiol. Biotechnol. 2012, 96, 773–781. [Google Scholar] [CrossRef] [PubMed]
- Horowitz Brown, S.; Zarnowski, R.; Sharpee, W.C.; Keller, N.P. Morphological transitions governed by density dependence and lipoxygenase activity in Aspergillus flavus. Appl. Environ. Microbiol. 2008, 74, 5674–5685. [Google Scholar] [CrossRef] [Green Version]
- Raina, S.; Odell, M.; Keshavarz, T. Quorum sensing as a method for improving sclerotiorin production in Penicillium sclerotiorum. J. Biotechnol. 2010, 148, 91–98. [Google Scholar] [CrossRef]
- Vitale, S.; Di Pietro, A.; Turrà, D. Autocrine pheromone signalling regulates community behaviour in the fungal pathogen Fusarium oxysporum. Nat. Microbiol. 2019, 4, 1443–1449. [Google Scholar] [CrossRef]
- Bartnicki-Garcia, S.; Nickerson, W.J. Nutrition, Growth, and Morphogenesis of Muscor rouxii. J. Bacteriol. 1962, 84, 841–858. [Google Scholar] [CrossRef] [Green Version]
- Muthukumar, G.; Jensen, E.C.; Nickerson, A.W.; Eckles, M.K.; Nickerson, K.W. Photomorphogenesis in Penicillium isariaeforme: Exogenous calcium substitutes for light. Photochem. Photobiol. 1991, 53, 287–291. [Google Scholar] [CrossRef]
- Gómez-Gil, E.; Franco, A.; Madrid, M.; Vázquez-Marín, B.; Gacto, M.; Fernández-Breis, J.; Vicente-Soler, J.; Soto, T.; Cansado, J. Quorum sensing and stress-activated MAPK signaling repress yeast to hypha transition in the fission yeast schizosaccharomyces Japonicus. PLoS Genet. 2019, 15, e1008192. [Google Scholar] [CrossRef]
- Mehmood, A.; Liu, G.; Wang, X.; Meng, G.; Wang, C.; Liu, Y. Fungal quorum-sensing molecules and inhibitors with potential antifungal activity: A review. Molecules 2019, 24, 1950. [Google Scholar] [CrossRef] [Green Version]
- Trinidad, V.; Bouzas, M. Developmental Biology in Prokaryotes and Lower Eukaryotes; Springer: Cham, Switzerland, 2021; ISBN 9783030775940. [Google Scholar]
- Chitarra, G.S.; Abee, T.; Rombouts, F.M.; Posthumus, M.A.; Dijksterhuis, J. Germination of Penicillium paneum conidia is regulated by 1-octen-3-ol, a volatile self-inhibitor. Appl. Environ. Microbiol. 2004, 70, 2823–2829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chitarra, G.S.; Abee, T.; Rombouts, F.M.; Dijksterhuis, J. 1-Octen-3-ol inhibits conidia germination of Penicillium paneum despite of mild effects on membrane permeability, respiration, intracellular pH, and changes the protein composition. FEMS Microbiol. Ecol. 2005, 54, 67–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morath, S.U.; Hung, R.; Bennett, J.W. Fungal volatile organic compounds: A review with emphasis on their biotechnological potential. Fungal Biol. Rev. 2012, 26, 73–83. [Google Scholar] [CrossRef]
- Tian, X.; Ding, H.; Ke, W.; Wang, L. Quorum Sensing in Fungal Species. Annu. Rev. Microbiol. 2021, 75, 449–469. [Google Scholar] [CrossRef] [PubMed]
- Franco, A.; Gacto, M.; Gómez-Gil, E.; Madrid, M.; Vicente-Soler, J.; Vázquez-Marín, B.; Cansado, J.; Soto, T. Quorum Sensing: A Major Regulator of Fungal Development. In Developmental Biology in Prokaryotes and Lower Eukaryotes; Villa, T.G., de Miguel Bouzas, T., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 331–366. ISBN 978-3-030-77595-7. [Google Scholar]
- Jamil, M.; Ali, A.; Ali, N.; Mujeeb-Kazi, A. Spot Blotch in Bread Wheat: Virulence, Resistance, and Breeding Perspectives; Elsevier Inc.: Amsterdam, The Netherlands, 2020; ISBN 9780128195277. [Google Scholar]
- Scherm, B.; Balmas, V.; Spanu, F.; Pani, G.; Delogu, G.; Pasquali, M.; Migheli, Q. Fusarium culmorum: Causal agent of foot and root rot and head blight on wheat. Mol. Plant Pathol. 2013, 14, 323–341. [Google Scholar] [CrossRef] [PubMed]
- Fiers, M.; Lognay, G.; Fauconnier, M.L.; Jijakli, M.H. Volatile Compound-Mediated Interactions between Barley and Pathogenic Fungi in the Soil. PLoS ONE 2013, 8, e66805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smiley, R.W.; Gourlie, J.A.; Easley, S.A.; Patterson, L.M.; Whittaker, R.G. Crop damage estimates for crown rot of wheat and barley in the Pacific Northwest. Plant Dis. 2005, 89, 595–604. [Google Scholar] [CrossRef] [Green Version]
- Vandendool, H.; Kratz, P.D. A generalization of the retention index system including linear temperature programmed gas-liquid partition chromatography. J. Chromatogr. 1963, 11, 463–471. [Google Scholar] [CrossRef]
- Savel’Eva, E.I.; Gavrilova, O.P.; Gagkaeva, T.Y. Study of the composition of volatile organic compounds emitted by the filamentous fungus Fusarium culmorum by gas chromatography-mass spectrometry combined with solid phase microextraction. Russ. J. Gen. Chem. 2014, 84, 2603–2610. [Google Scholar] [CrossRef]
- Takeuchi, T.; Kimura, T.; Tanaka, H.; Kaneko, S.; Ichii, S.; Kiuchi, M.; Suzuki, T. Analysis of volatile metabolites emitted by soil-derived fungi using head space solid-phase microextraction/gas chromatography/mass spectrometry: I. Aspergillus fumigatus, Aspergillus nidulans, Fusarium solani and Penicillium paneum. Surf. Interface Anal. 2012, 44, 694–698. [Google Scholar] [CrossRef]
- Savelieva, E.I.; Gustyleva, L.K.; Kessenikh, E.D.; Khlebnikova, N.S.; Leffingwell, J.; Gavrilova, O.P.; Gagkaeva, T.Y. Study of the Vapor Phase Over Fusarium Fungi Cultured on Various Substrates. Chem. Biodivers. 2016, 13, 891–903. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Hung, R.; Yap, M.; Bennett, J.W. Age matters: The effects of volatile organic compounds emitted by Trichodermaatroviride on plant growth. Arch. Microbiol. 2015, 197, 723–727. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Cale, J.A.; Hussain, A.; Erbilgin, N. Exposure to Fungal Volatiles Can Influence Volatile Emissions From Other Ophiostomatoid Fungi. Front. Microbiol. 2020, 11, 567462. [Google Scholar] [CrossRef] [PubMed]
- Rinkel, J.; Babczyk, A.; Wang, T.; Stadler, M.; Dickschat, J.S. Volatiles from the hypoxylaceous fungi Hypoxylon griseobrunneum and Hypoxylon macrocarpum. Beilstein J. Org. Chem. 2018, 14, 2974–2990. [Google Scholar] [CrossRef] [Green Version]
- Ponce, M.A.; Lizarraga, S.; Bruce, A.; Kim, T.N.; Morrison, W.R. Grain Inoculated with Different Growth Stages of the Fungus, Aspergillus flavus, Affect the Close-Range Foraging Behavior by a Primary Stored Product Pest, Sitophilus oryzae (Coleoptera: Curculionidae). Environ. Entomol. 2022, 51, 927–939. [Google Scholar] [CrossRef]
- Wu, W.; Tran, W.; Taatjes, C.A.; Alonso-Gutierrez, J.; Lee, T.S.; Gladden, J.M. Rapid discovery and functional characterization of terpene synthases from four endophytic xylariaceae. PLoS ONE 2016, 11, e0146983. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Cheng, Y.; Yang, M.; Liu, Y.; Chen, K.; Long, C.A.; Deng, X. Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of Penicillium molds of citrus fruits. BMC Microbiol. 2014, 14, 242. [Google Scholar] [CrossRef] [Green Version]
- Korsak, Z.; Rydzyński, K. Neurotoxic effects of acute and subchronic inhalation exposure to trimethylbenzene isomers (pseudocumene, mesitylene, hemimellitene) in rats. Int. J. Occup. Med. Environ. Health 1996, 9, 341–349. [Google Scholar]
- Josselin, L.; De Clerck, C.; De Boevre, M.; Moretti, A.; Haïssam Jijakli, M.; Soyeurt, H.; Fauconnier, M.L. Volatile organic compounds emitted by Aspergillus flavus strains producing or not aflatoxin B1. Toxins 2021, 13, 705. [Google Scholar] [CrossRef]
- del Pilar Marquez-Villavicencio, M.; Weber, B.; Witherell, R.A.; Willis, D.K.; Charkowski, A.O. The 3-hydroxy-2-butanone pathway is required for pectobacterium carotovorum pathogenesis. PLoS ONE 2011, 6, e22974. [Google Scholar] [CrossRef]
- Xiao, Z.; Lu, J.R. Generation of acetoin and its derivatives in foods. J. Agric. Food Chem. 2014, 62, 6487–6497. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Gozz, K.; Yan, F.; Chai, Y. Acetic acid acts as a volatile signal to stimulate bacterial biofilm formation. MBio 2015, 6, 406–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fialho, M.B.; de Moraes, M.H.D.; Tremocoldi, A.R.; Pascholati, S.F. Potential of antimicrobial volatile organic compounds to control Sclerotinia sclerotiorum in bean seeds. Pesqui. Agropecu. Bras. 2011, 46, 137–142. [Google Scholar] [CrossRef] [Green Version]
- Saeidi, S.; Hassanpour, K.; Ghamgosha, M.; Heiat, M.; Taheri, R.A.; Mirhosseini, A.; Farnoosh, G. Antibacterial activity of ethyl acetate and aqueous extracts of Mentha longifolia L. and hydroalcoholic extract of Zataria multiflora Boiss. plants against important human pathogens. Asian Pac. J. Trop. Med. 2014, 7, S186–S189. [Google Scholar] [CrossRef] [Green Version]
- Ando, H.; Hatanaka, K.; Ohata, I.; Yamashita-Kitaguchi, Y.; Kurata, A.; Kishimoto, N. Antifungal activities of volatile substances generated by yeast isolated from Iranian commercial cheese. Food Control 2012, 26, 472–478. [Google Scholar] [CrossRef]
- Pimenta, L.; Ferreira, M.A.; Pedroso, M.P.; Campos, V.P. Wood-associated fungi produce volatile organic compounds toxic to root-knot nematode. Sci. Agric. 2017, 74, 303–310. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, N.M.; Shinde, R.B.; Karuppayil, S.M. Effect of alcohols on filamentation, growth, viability and biofilm development in Candida albicans. Brazilian J. Microbiol. 2013, 44, 1315–1320. [Google Scholar] [CrossRef] [Green Version]
- Micalizzi, E.W.; Mack, J.N.; White, G.P.; Avis, T.J.; Smith, M.L. Microbial inhibitors of the fungus Pseudogymnoascus destructans, the causal agent of white-nose syndrome in bats. PLoS ONE 2017, 12, e0179770. [Google Scholar] [CrossRef] [Green Version]
- Zunino, M.P.; Herrera, J.M.; Pizzolitto, R.P.; Rubinstein, H.R.; Zygadlo, J.A.; Dambolena, J.S. Effect of Selected Volatiles on Two Stored Pests: The Fungus Fusarium verticillioides and the Maize Weevil Sithophilus zeamais. J. Agric. Food Chem. 2015, 63, 7743–7749. [Google Scholar] [CrossRef] [Green Version]
- Paul, D.; Park, K.S. Identification of volatiles produced by Cladosporium cladosporioides CL-1, a fungal biocontrol agent that promotes plant growth. Sensors 2013, 13, 13969–13977. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Y.; Tan, X.M.; Yang, J.; Guo, L.P.; Ding, G. Naturally Occurring seco-Sativene Sesquiterpenoid: Chemistry and Biology. J. Agric. Food Chem. 2020, 68, 9827–9838. [Google Scholar] [CrossRef] [PubMed]
- Bonfante, P.; Splivallo, R.; Novero, M.; Bertea, C.M.; Bossi, S. Truffle volatiles inhibit growth and induce an oxidative burst in Arabidopsis thaliana. New Phytol. 2007, 175, 417–424. [Google Scholar]
- Bruce, A.; Stewart, D.; Verrall, S.; Wheatley, R.E. Effect of volatiles from bacteria and yeast on the growth and pigmentation of sapstain fungi. Int. Biodeterior. Biodegrad. 2003, 51, 101–108. [Google Scholar] [CrossRef]
- Yang, Q.; Chen, Y.; Ma, Z. Involvement of BcVeA and BcVelB in regulating conidiation, pigmentation and virulence in Botrytis cinerea. Fungal Genet. Biol. 2013, 50, 63–71. [Google Scholar] [CrossRef]
- Tournu, H.; Carroll, J.; Latimer, B.; Dragoi, A.M.; Dykes, S.; Cardelli, J.; Peters, T.L.; Eberle, K.E.; Palmer, G.E. Identification of small molecules that disrupt vacuolar function in the pathogen Candida albicans. PLoS ONE 2017, 12, e0171145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Josselin, L.; De Clerck, C.; De Boevre, M.; Moretti, A.; Soyeurt, H.; Fauconnier, M.-L. Fungal volatile organic compounds, can be used to develop aflatoxin-specific sensors? In Proceedings of the MycoTWIN-MycoKey-2021 International Conference, Bari, Italy, 9–12 November 2021. [Google Scholar]
- Terra, W.C.; Campos, V.P.; Martins, S.J.; Costa, L.S.A.S.; da Silva, J.C.P.; Barros, A.F.; Lopez, L.E.; Santos, T.C.N.; Smant, G.; Oliveira, D.F. Volatile organic molecules from Fusarium oxysporum strain 21 with nematicidal activity against Meloidogyne incognita. Crop Prot. 2018, 106, 125–131. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IUPAC Name | F. culmorum | C. sativus |
|---|---|---|
| (+)-Sativene | x | |
| Longifolene | x | |
| 3-Methylbutyl ethanoate | x | |
| Pentan-1-ol | x | |
| Toluene | x | |
| 2-Methylpropan-1-ol | x | x |
| 3-Methylbutan-1-ol | x | x |
| Ethyl acetate | x | x |
| Emitted by | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F. culmorum | C. sativus | |||||||||||
| a | Compound name | CAS b | Lit. c | RIcal d | RIstd e | RIref f | Identification g | Ref RI h | NC | O | NC | O |
| Alcohol | 2-Phenylethanol | 60-12-8 | 1, 2 | 1924 | 1924 | MS, RI | NIST | 7 | ||||
| 3-Methylbutan-1-ol | 123-51-3 | 2 | 1201 | 1201 | 1201 | MS, RI, STD | NIST | 7, 14 | 7, 14 | 7, 14 | 7, 14 | |
| 2-Methylpropan-1-ol | 78-83-1 | 1 | 1062 | 1060 | MS, RI | NIST | 14 | 7, 14 | 14 | 7, 14 | ||
| Pentan-1-ol | 71-41-0 | 1, 2 | 1270 | 1249 | 1260 | MS, RI, STD | NIST | 7 | ||||
| Propan-1-ol | 71-23-8 | 1, 3 | 983 | nd | MS, RI, STD | / | 7, 14 | |||||
| Alkene | 2,6-Dimethylanisole | 22469-52-9 | / | 1397 | nd | MS, RI | / | 7 | ||||
| Hemellitol | 526-73-8 | / | 1318 | 1320.6 | MS, RI | NIST | 7 | |||||
| Isoprene | 78-79-5 | 5 | nd | 602 | MS, RI | NIST | 7, 14 | 7, 14 | ||||
| Mesitylene | 108-67-8 | 2 | 1247 | 1246 | MS, RI | NIST | 7, 14 | |||||
| Pseudocumene | 95-63-6 | 2 | 1249 | 1249 | MS, RI | NIST | 14 | |||||
| p-Xylene | 106-42-3 | 3 | 1066 | 1165 | MS, RI | NIST | 7, 14 | |||||
| Toluene | 108-88-3 | 1 | 1021 | 1017 | 1020 | MS, RI, STD | NIST | 7, 14 | 7, 14 | |||
| Alkane | Pentane | 109-66-0 | 3 | nd | 500 | MS, RI | / | 14 | 7, 14 | |||
| Esrer | 3-Methylbutyl ethanoate | 123-92-2 | 1 | 1169 | 1186 | 1167 | MS, RI, STD | NIST | 7 | 7, 14 | ||
| Ethyl 2-methylbutyrate | 7452-79-1 | 1, 2 | 1022 | 1022 | MS, RI | NIST | 7, 14 | |||||
| Ethyl 3-methylbutanoate | 108-64-5 | 1, 2 | 1049 | 1049 | MS, RI | NIST | 14 | 14 | ||||
| Ethyl acetate | 141-78-6 | 1, 3 | 888.5 | 901 | 900 | MS, RI, STD | NIST | 7, 14 | 7, 14 | 7, 14 | ||
| Ketone | Acetoin | 513-86-0 | 1 | 1285.9 | 1286 | MS, RI | NIST | 14 | ||||
| Butan-2-one | 78-93-3 | 1 | nd | nd | MS, RI | / | 7, 14 | |||||
| Terpene | (-)-β-Acoradiene | 090457-37-7 | 2, 3, 5 | 1687.4 | 1693 | MS, RI | NIST | 7 | 7, 14 | |||
| (+)-Aromadendrene | 489-39-4 | 1 | 1652.9 | 1650 | MS, RI | Davies (1990) | 7, 14 | 7 | ||||
| (+)-Sativene | 3650-28-0 | 1, 5 | 1524.5 | 1537 | 1527 | MS, RI, STD | NIST | 7, 14 | 7, 14 | |||
| Cycloisosativene | 22469-52-9 | / | 1410 | 1483 | Internal database | 14 | 14 | |||||
| Epi-bicyclosesquiphellandrene | 54324-03-7 | 1, 3 | 1627.7 | 1633 | MS, RI | NIST | 7, 14 | 14 | ||||
| Germacrene A | 75023-40-4 | 1, 5 | 1744.9 | 1745 | MS, RI | NIST | 7, 14 | 7, 14 | ||||
| Limonene | 3387-41-5 | 1, 3 | 1242 | 1234 | MS, RI | pherobase | 7 | 7 | ||||
| Longifolene | 475-20-7 | 3, 5 | 1589.6 | 1600 | 1590 | MS, RI, STD | NIST | 7, 14 | 7, 14 | |||
| Unkown sesquiterpene 1 | nd | / | nd | nd | MS, RI | / | 7, 14 | |||||
| Unkown sesquiterpene 2 | nd | / | 1674.4 | nd | MS, RI | NIST | 7 | 7 | ||||
| α-Chamigrene | 19912-83-5 | 1, 2 | 1661.7 | nd | MS, RI | / | 7, 14 | 14 | ||||
| α-Gurjunene | 489-40-7 | 1 | 1535 | 1529 | Internal database | 7, 14 | ||||||
| β-Phellandrene | 555-10-2 | 3 | 1202.9 | 1202 | MS, RI | NIST | 7, 14 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Clerck, C.; Josselin, L.; Vangoethem, V.; Lassois, L.; Fauconnier, M.-L.; Jijakli, H. Weapons against Themselves: Identification and Use of Quorum Sensing Volatile Molecules to Control Plant Pathogenic Fungi Growth. Microorganisms 2022, 10, 2459. https://doi.org/10.3390/microorganisms10122459
De Clerck C, Josselin L, Vangoethem V, Lassois L, Fauconnier M-L, Jijakli H. Weapons against Themselves: Identification and Use of Quorum Sensing Volatile Molecules to Control Plant Pathogenic Fungi Growth. Microorganisms. 2022; 10(12):2459. https://doi.org/10.3390/microorganisms10122459
Chicago/Turabian StyleDe Clerck, Caroline, Laurie Josselin, Valentine Vangoethem, Ludivine Lassois, Marie-Laure Fauconnier, and Haïssam Jijakli. 2022. "Weapons against Themselves: Identification and Use of Quorum Sensing Volatile Molecules to Control Plant Pathogenic Fungi Growth" Microorganisms 10, no. 12: 2459. https://doi.org/10.3390/microorganisms10122459