

Metagenome-Based Exploration of Bacterial Communities Associated with Cyanobacteria Strains Isolated from Thermal Muds

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
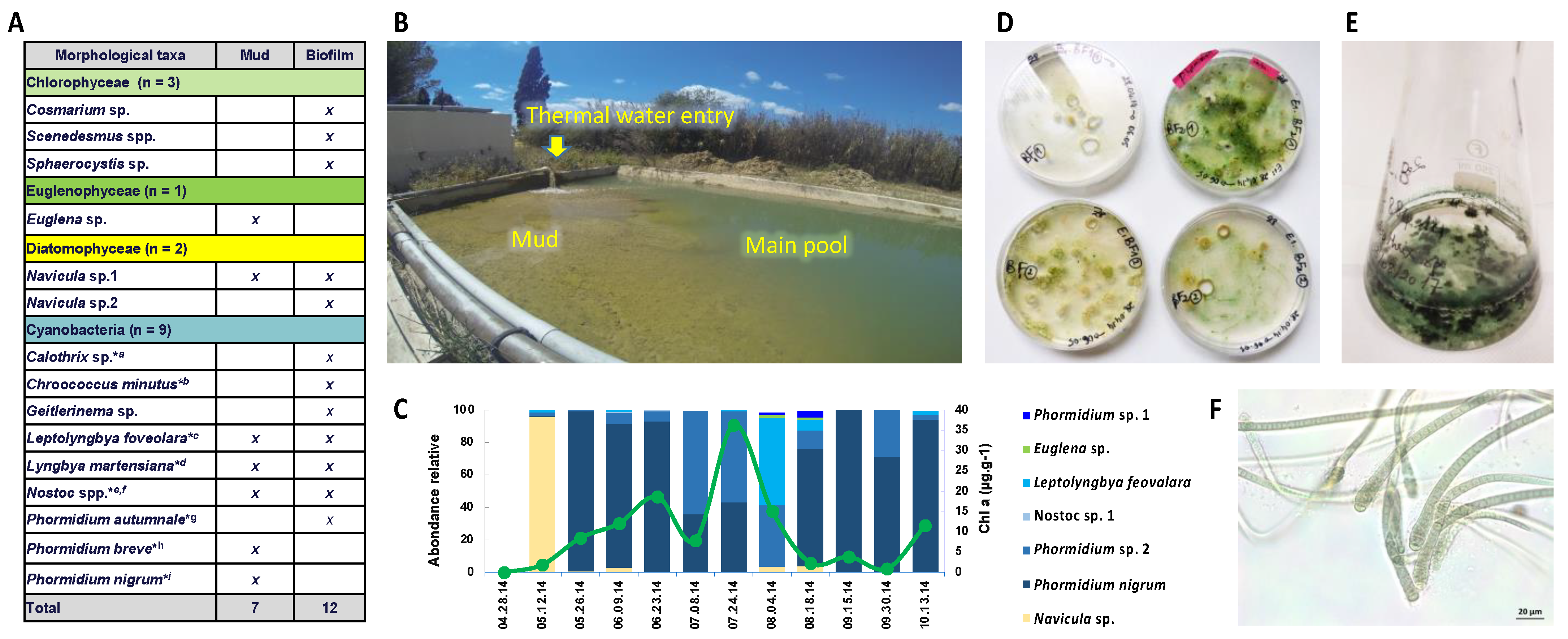
2.1. Field Sampling and Cyanobacteria Isolation
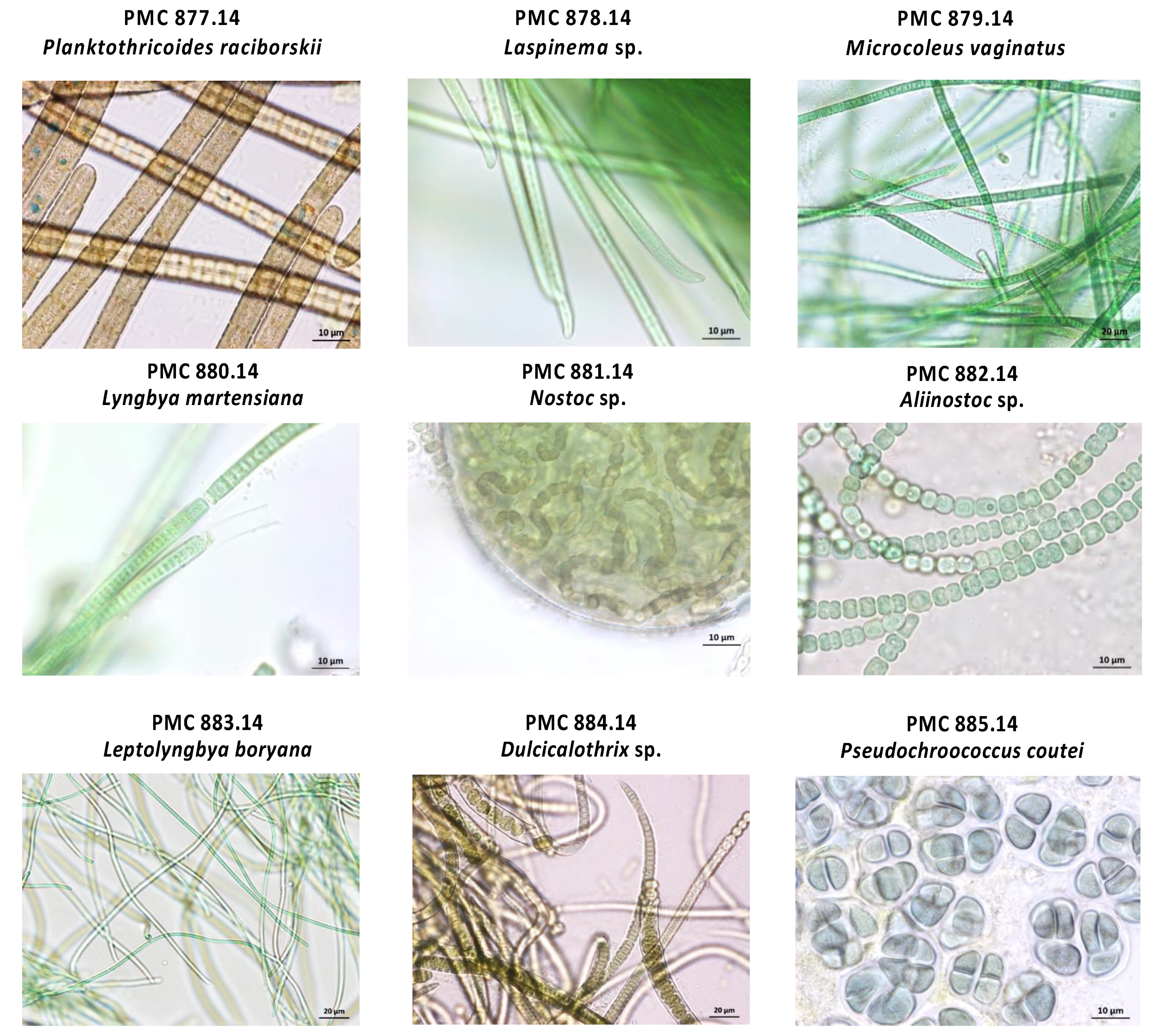
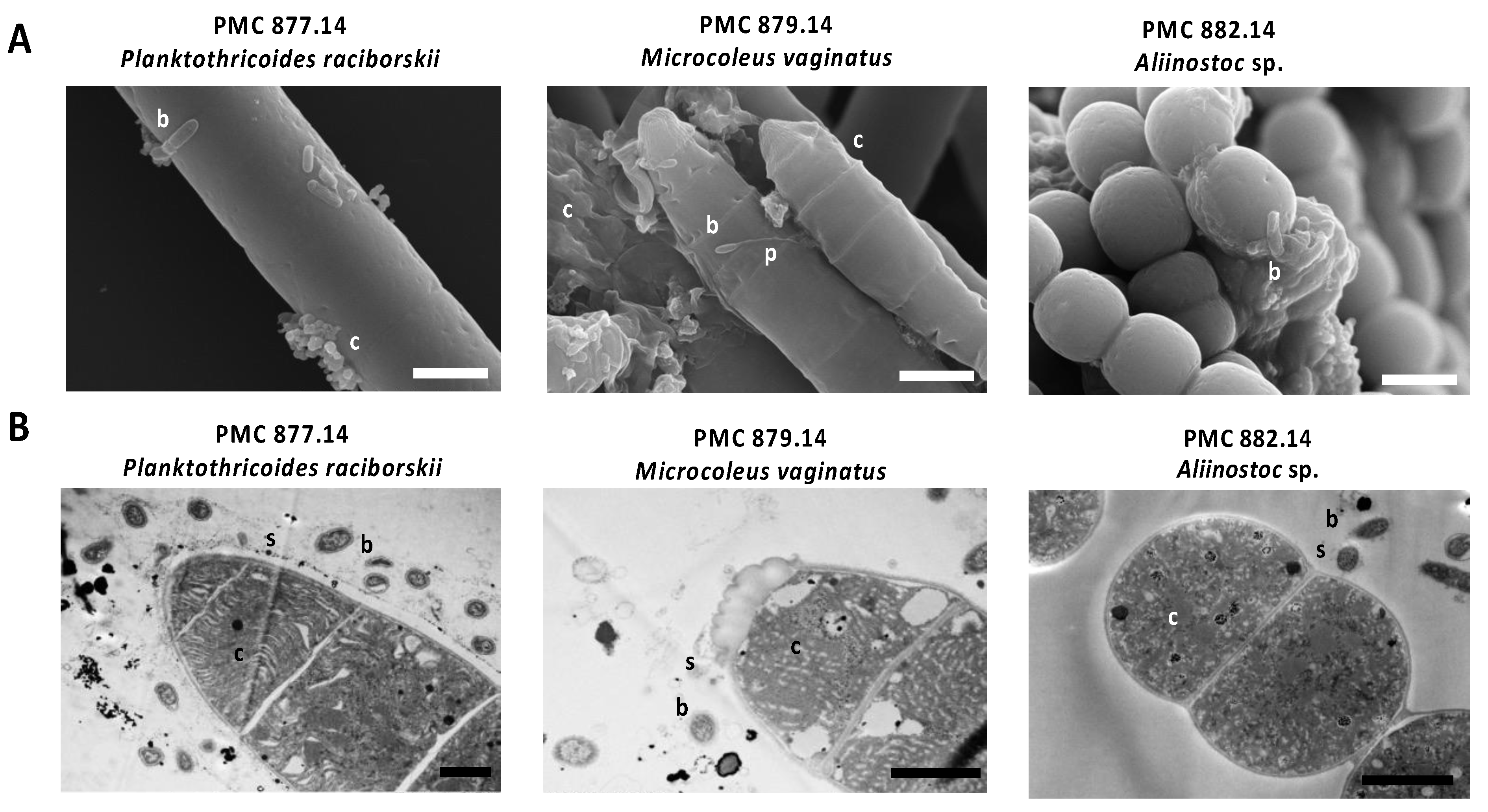
2.2. Morphological and Ultrastructural Analyses of Cyanospheres
2.3. Genomic DNA Extraction
2.4. Taxonomic Composition of Mud Samples and Cyanosphere
2.5. Metagenome-Assembled Genomes from Cyanobacteria and Heterotrophs
3. Results and Discussion
3.1. Cyanobacteria Isolates
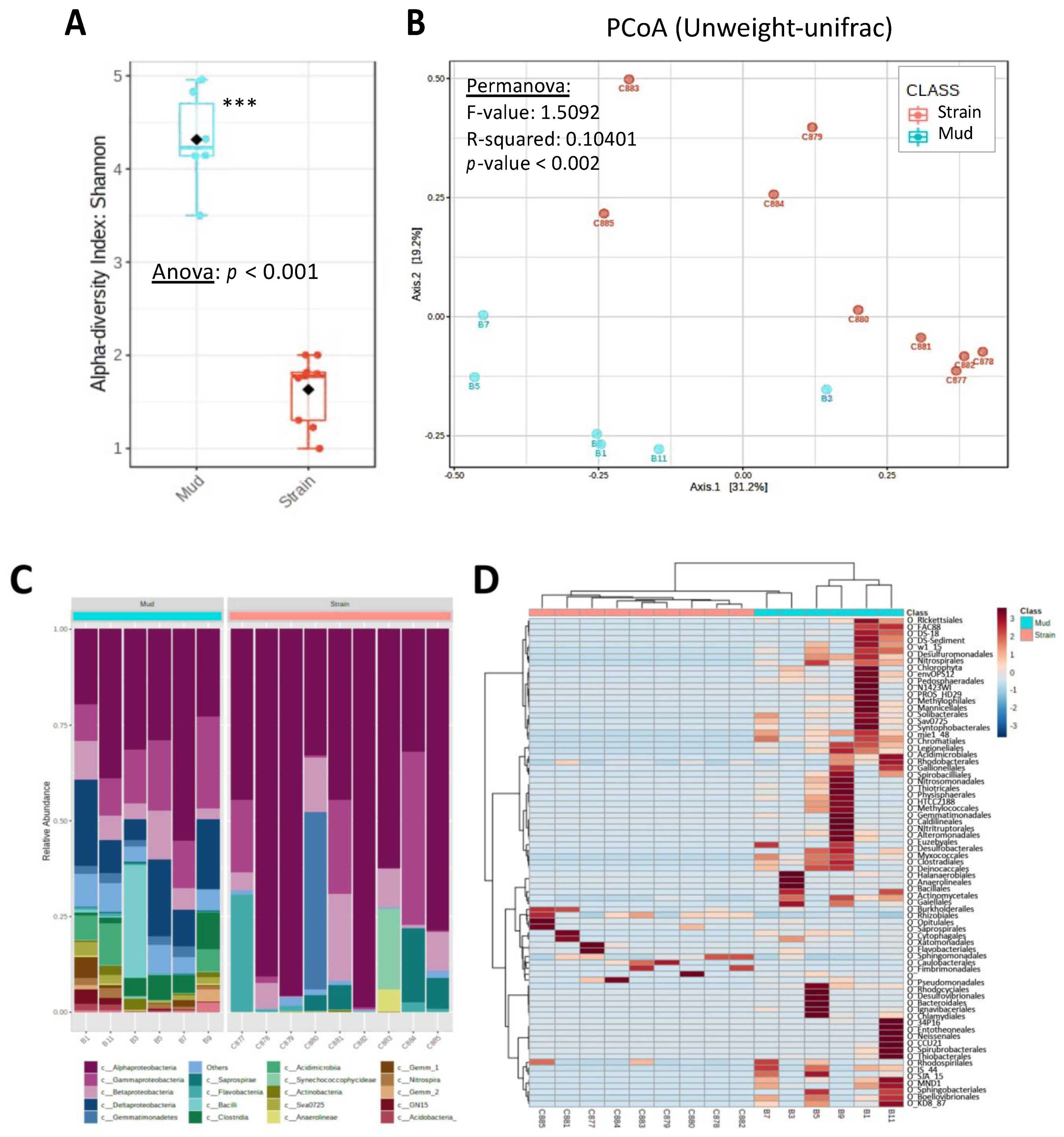
3.2. Cyanosphere Composition
3.3. Cyanosphere’s Main Actors Revealed by Metagenomics
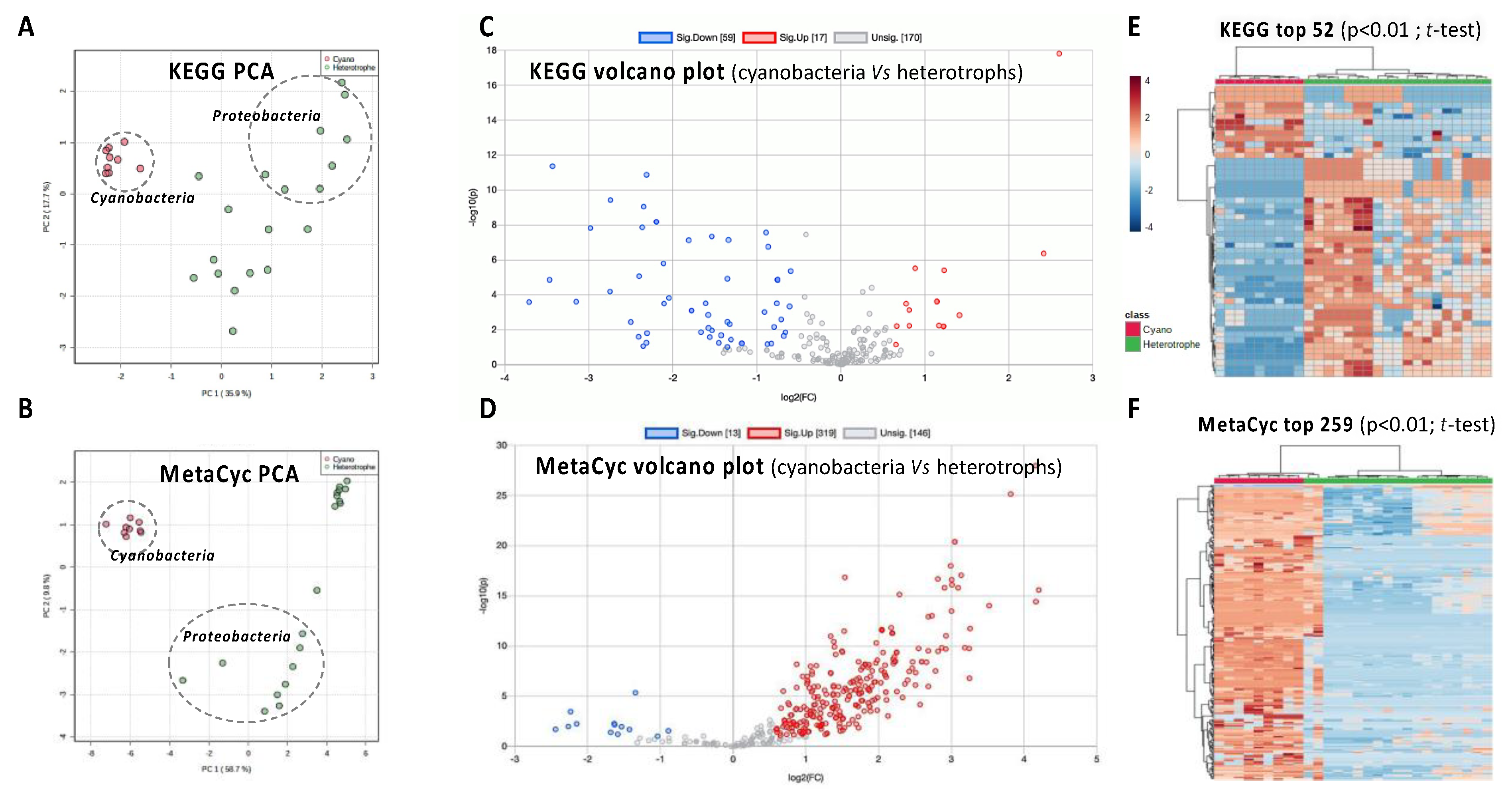
3.4. Functional Genomics of the Cyanosphere
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Whitton, B.; Potts, M. Introduction to the Cyanobacteria. In Ecology of Cyanobacteria: Their Diversity in Time and Space; Springer Science & Business Media: Dordrecht, The Netherlands, 2000. [Google Scholar]
- Whitton, B.A. (Ed.) Ecology of Cyanobacteria II: Their Diversity in Space and Time; Whitton, B.A. (Ed.) Springer Science & Business Media: Dordrecht, The Netherlands, 2012. [Google Scholar]
- Mazard, S.; Penesyan, A.; Ostrowski, M.; Paulsen, I.; Egan, S. Tiny Microbes with a Big Impact: The Role of Cyanobacteria and Their Metabolites in Shaping Our Future. Mar. Drugs 2016, 14, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seymour, J.R.; Amin, S.A.; Raina, J.B.; Stocker, R. Zooming in on the phycosphere: The ecological interface for phytoplankton–bacteria relationships. Nat. Microbiol. 2017, 2, 17065. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.K.; Hutchison, J.R.; Renslow, R.S.; Kim, Y.M.; Chrisler, W.B.; Engelmann, H.E.; Dohnalkova, A.C.; Hu, D.H.; Metz, T.O.; Fredrickson, J.K.; et al. Phototrophic biofilm assembly in microbial-mat-derived unicyanobacterial consortia: Model systems for the study of autotroph-heterotroph interactions. Front. Microbiol. 2014, 5, 109. [Google Scholar] [CrossRef] [PubMed]
- Morris, J.J.; Kirkegaard, R.; Szul, M.J.; Johnson, Z.I.; Zinser, E.R. Facilitation of robust growth of Prochlorococcus colonies and dilute liquid cultures by “helper” heterotrophic bacteria. Appl. Environ. Microbiol. 2008, 74, 4530–4534. [Google Scholar] [CrossRef] [Green Version]
- Jackrel, S.L.; Yang, J.W.; Schmidt, K.C.; Denef, V.J. Host specificity of microbiome assembly and its fitness effects in phytoplankton. ISME J. 2021, 15, 774–788. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Wang, Y.; Lu, J.; Lin, W.; Chen, F.; Jiao, N. Metagenomic and metaproteomic insights into photoautotrophic and heterotrophic interactions in a Synechococcus culture. mBio 2020, 11, e03261-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demay, J.; Halary, S.; Knittel-Obrecht, A.; Villa, P.; Duval, C.; Hamlaoui, S.; Roussel, T.; Yéprémian, C.; Reinhardt, A.; Bernard, C.; et al. Anti-inflammatory, antioxidant, and wound-healing properties of cyanobacteria from thermal mud of Balaruc-Les-Bains, France: A multi-approach study. Biomolecules 2020, 11, 28. [Google Scholar] [CrossRef] [PubMed]
- Demay, J.; Bernard, C.; Reinhardt, A.; Marie, B. Natural Products from Cyanobacteria: Focus on Beneficial Activities. Mar. Drugs 2019, 17, 320. [Google Scholar] [CrossRef] [Green Version]
- Cai, H.Y.; Yan, Z.S.; Wang, A.J.; Krumholz, L.R.; Jiang, H.L. Analysis of the attached microbial community on mucilaginous cyanobacterial aggregates in the eutrophic Lake Taihu reveals the importance of Planctomycetes. Microb. Ecol. 2013, 66, 73–83. [Google Scholar] [CrossRef]
- Decho, A.W. Microbial bio fi lms in intertidal systems: An overview. Cont. Shelf Res. 2000, 20, 1257–1273. [Google Scholar] [CrossRef]
- Gris, B.; Treu, L.; Zampieri, R.M.; Caldara, F.; Romualdi, C.; Campanaro, S.; La Rocca, N. Microbiota of the therapeutic euganean thermal muds with a focus on the main cyanobacteria species. Microorganisms 2020, 8, 1590. [Google Scholar] [CrossRef] [PubMed]
- Baudinat, C. Contribution à L’étude de la Maturation de Péloïdes: Application Aux Stations Thermales de Balaruc-Les-Bains (34) et Cransac (12). Ph.D. Thesis, University of Montpellier 1, Montpellier, France, 1986. [Google Scholar]
- Dupuis, E. Pré-étude relative à l’évaluation de la production algale des eaux thermales de Balaruc-les-Bains. Master Thesis, University of Montpellier 1, Montpellier, France, 1987. [Google Scholar]
- Duval, C.; Hamlaoui, S.; Piquet, B.; Toutirais, G.; Yéprémian, C.; Reinhardt, A.; Duperron, S.; Marie, B.; Demay, J.; Bernard, C. Diversity of cyanobacteria from thermal muds (Balaruc-Les-Bains, France) with the description of Pseudochroococcus coutei gen. nov., sp. nov. FEMS Microbes 2021, 2, xtab006. [Google Scholar] [CrossRef]
- Ward, D.M.; Castenholz, R.W.; Miller, S.R. Cyanobacteria in geothermal habitats. In Ecology of Cyanobacteria II; Springer: Dordrecht, The Netherlands, 2012; pp. 39–63. [Google Scholar]
- Nelson, C.; Giraldo-Silva, A.; Garcia-Pichel, F. A symbiotic nutrient exchange within the cyanosphere microbiome of the biocrust cyanobacterium, Microcoleus vaginatus. ISME J. 2021, 15, 282–292. [Google Scholar] [CrossRef] [PubMed]
- Cook, K.V.; Li, C.; Cai, H.; Krumholz, L.R.; Hambright, K.D.; Paerl, H.W.; Steffen, M.M.; Wilson, A.E.; Burford, M.A.; Grossart, H.P.; et al. The global Microcystis interactome. Limnol. Oceanogr. 2020, 65, S194–S207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Carrascal, O.M.; Tromas, N.; Terrat, Y.; Moreno, E.; Giani, A.; Corrêa Braga Marques, L.; Fortin, N.; Shapiro, B.J. Single-colony sequencing reveals microbe-by-microbiome phylosymbiosis between the cyanobacterium Microcystis and its associated bacteria. Microbiome 2021, 9, 194. [Google Scholar] [CrossRef] [PubMed]
- Rippka, R. Isolation and purification of cyanobacteria. Methods Enzymol. 1988, 167, 3–27. [Google Scholar]
- Hamlaoui, S.; Yéprémian, C.; Duval, C.; Marie, B.; Djédiat, C.; Piquet, B.; Bernard, C.; Duperron, S. The culture collection of cyanobacteria and microalgae at the French National Museum of Natural History: A century old but still alive and kicking! Including in memoriam: Professor Alain Couté. Cryptogam. Algol. 2022, 43, 41–83. [Google Scholar] [CrossRef]
- Parveen, B.; Ravet, V.; Djediat, C.; Mary, I.; Quiblier, C.; Debroas, D.; Humbert, J.F. Bacterial communities associated with M icrocystis colonies differ from free-living communities living in the same ecosystem. Environ. Microbiol. Rep. 2013, 5, 716–724. [Google Scholar]
- Stanier, R.Y.; Deruelles, J.; Rippka, R.; Herdman, M.; Waterbury, J.B. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. Microbiology 1979, 111, 1–61. [Google Scholar]
- Parada, A.E.; Needham, D.M.; Fuhrman, J.A. Every base matters: Assessing small subunit rRNA primers for marine microbiomes with mock communities, time series and global field samples. Environ. Microbiol. 2016, 18, 1403–1414. [Google Scholar] [CrossRef] [PubMed]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Holmes, S.P. Exact sequence variants should replace operational taxonomic units in marker-gene data analysis. ISME J. 2017, 11, 2639–2643. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Liu, P.; Zhou, G.; Xia, J. Using Microbiome Analyst for comprehensive statistical, functional, and meta-analysis of microbiome data. Nat. Protoc. 2020, 15, 799–821. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.; Nikolenko, S.; Pham, S.; Prjibelski, A.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [Green Version]
- Cambuy, D.; Coutinho, F.; Dutilh, B. Contig annotation tool CAT robustly classifies assembled metagenomic contigs and long sequences. bioRxiv 2016, 072868. [Google Scholar] [CrossRef] [Green Version]
- Pruesse, E.; Peplies, J.; Glöckner, F.O. SINA: Accurate high-throughput multiple sequence alignment of ribosomal RNA genes. Bioinformatics 2012, 28, 1823–1829. [Google Scholar] [CrossRef] [Green Version]
- Vallenet, D.; Calteau, A.; Dubois, M.; Amours, P.; Bazin, A.; Beuvin, M.; Burlot, L.; Bussell, X.; Fouteau, S.; Gautreau, G.; et al. MicroScope: An Integrated Platform for the Annotation and Exploration of Microbial Gene Functions through Genomic, Pangenomic and Metabolic Comparative Analysis. Nucleic Acids Res. 2019, 48, D579–D589. [Google Scholar] [CrossRef] [Green Version]
- Parks, D.H.; Imelfort, M.; Skennerton, C.; Hugenholtz, P.; Tyson, G. CheckM: Assessing the Quality of Microbial Genomes Recovered from Isolates, Single Cells, and Metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altman, T.; Travers, M.; Kothari, A.; Caspi, R.; Karp, P.D. A systematic comparison of the MetaCyc and KEGG pathway databases. BMC Bioinform. 2013, 14, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chong, J.; Wishart, D.S.; Xia, J. Using metaboanalyst 4.0 for comprehensive and integrative metabolomics data analysis. Curr. Protoc. Bioinform. 2019, 68, e86. [Google Scholar] [CrossRef] [PubMed]
- Waterbury, J.B. The cyanobacteria—Isolation, purification and identification. In The Prokaryotes; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; Volume 4, pp. 1053–1073. [Google Scholar]
- Bagatini, I.L.; Eiler, A.; Bertilsson, S.; Klaveness, D.; Tessarolli, L.P.; Vieira, A.A.H. Host-specificity and dynamics in bacterial communities associated with bloom-forming freshwater phytoplankton. PLoS ONE 2014, 9, e85950. [Google Scholar] [CrossRef] [Green Version]
- Kaplan Can, H.; Gurbuz, F.; Odabaşı, M. Partial characterization of cyanobacterial extracellular polymeric substances for aquatic ecosystems. Aquat. Ecol. 2019, 53, 431–440. [Google Scholar] [CrossRef]
- Escalas, A.; Troussellier, M.; Melayah, D.; Bruto, M.; Nicolas, S.; Bernard, C.; Hugoni, M. Strong reorganization of multi-domain microbial networks associated with primary producers sedimentation from oxic to anoxic conditions in an hypersaline lake. FEMS Microbiol. Ecol. 2021, 97, fiab163. [Google Scholar] [CrossRef]
- Paerl, H.W. Specific associations of the bluegreen algae Anabaena and Aphanizomenon with bacteria in Freshwater blooms. J. Phycol. 1976, 12, 431–435. [Google Scholar] [CrossRef]
- Louati, I.; Pascault, N.; Debroas, D.; Bernard, C.; Humbert, J.F.; Leloup, J. Structural diversity of bacterial communities associated with bloom-forming freshwater cyanobacteria differs according to the cyanobacterial genus. PLoS ONE 2015, 10, e0140614. [Google Scholar] [CrossRef] [Green Version]
- Rastogi, R.P.; Sinha, R.P.; Singh, S.P.; Häder, D.P. Photoprotective compounds from marine organisms. J. Ind. Microbiol. Biotechnol. 2010, 37, 537–558. [Google Scholar] [CrossRef]
- Wang, Z.; Sun, Q.; Zhang, H.; Wang, J.; Fu, Q.; Qiao, H.; Wang, Q. Insight into antibacterial mechanism of polysaccharides: A review. LWT 2021, 150, 111929. [Google Scholar] [CrossRef]
- Taylor, J.D.; Cottingham, S.D.; Billinge, J.; Cunliffe, M. Seasonal microbial community dynamics correlate with phytoplankton-derived polysaccharides in surface coastal waters. ISME J. 2014, 8, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Buchan, A.; LeCleir, G.R.; Gulvik, C.A.; González, J.M. Master recyclers: Features and functions of bacteria associated with phytoplankton blooms. Nat. Rev. Microbiol. 2014, 12, 686–698. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Zancarini, A.; Louati, I.; De Cesare, S.; Duval, C.; Tambosco, K.; Bernard, C.; Debroas, D.; Song, L.; Leloup, J.; et al. Bacterial communities associated with four cyanobacterial genera display structural and functional differences: Evidence from an experimental approach. Front. Microbiol. 2016, 7, 1662. [Google Scholar] [CrossRef] [PubMed]
- Semenov, M.V. Metabarcoding and metagenomics in soil ecology research: Achievements, challenges, and prospects. Biol. Bull. Rev. 2021, 11, 40–53. [Google Scholar] [CrossRef]
- Halary, S.; Duperron, S.; Tiam, S.K.; Duval, C.; Dhénain, E.; Bernard, C.; Marie, B. Unexpected Micro-Spatial Scale Genomic Diversity of the Bloom-Forming Cyanobacterium Aphanizomenon Gracile and its Phycosphere. Preprint 2021. [Google Scholar] [CrossRef]
- Pascault, N.; Rué, O.; Loux, V.; Pédron, J.; Martin, V.; Tambosco, J.; Bernard, C.; Humbert, J.F.; Leloup, J. Insights into the cyanosphere: Capturing the respective metabolisms of cyanobacteria and chemotrophic bacteria in natural conditions? Environ. Microbiol. Rep. 2021, 13, 364–374. [Google Scholar] [CrossRef]
- Bouma-Gregson, K.; Olm, M.R.; Probst, A.J.; Anantharaman, K.; Power, M.E.; Banfield, J.F. Microbial diversity and metabolic potential in cyanotoxin producing cyanobacterial mats throughout a river network. bioRxiv 2018, 294421. [Google Scholar] [CrossRef]
- Hirschberg, J.; Chamovitz, D. Carotenoids in Cyanobacteria BT. In The Molecular Biology of Cyanobacteria; Springer: Dordrecht, The Netherlands, 1994; pp. 559–579. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain__MAG (16S Phylum Annotation) | nb_contigs | Size (pb) | GC% | nb CDSs | Microscope Affiliation |
|---|---|---|---|---|---|
| PMC 877__Bacteroidetes | 8 | 2,940,501 | 32.92 | 2707 | Bacteroidetes |
| PMC 877__Cyanobacteria | 15 | 7,391,557 | 43.50 | 7130 | Planktothricoides |
| PMC 877__Proteobacteria | 7 | 3,625,685 | 64.63 | 3616 | Sphingomonadales |
| PMC 878__Cyanobacteria | 69 | 7,336,742 | 47.37 | 5829 | Laspinema |
| PMC 878__Proteobacteria | 45 | 12,013,276 | 65.81 | 11,940 | Proteobacteria |
| PMC 879__Cyanobacteria | 34 | 6,900,942 | 45.68 | 6400 | Microcoleus |
| PMC 879__Proteobacteria | 11 | 10,840,780 | 67.92 | 10,627 | Proteobacteria |
| PMC 880__Bacteroidetes | 1803 | 14,271,057 | 48.22 | 14,012 | Bacteroidetes |
| PMC 880__Cyanobacteria | 132 | 6,444,104 | 39.60 | 5904 | Lyngbya |
| PMC 880__Gemmatimonadetes | 25 | 4,542,236 | 65.84 | 4112 | Gemmatimonadetes |
| PMC 880__Planctomycetes | 216 | 6,159,044 | 53.18 | 5068 | Planctomycetes |
| PMC 880__Proteobacteria | 2151 | 21,578,483 | 64.87 | 22,250 | Proteobacteria |
| PMC 881__Cyanobacteria | 648 | 7,938,353 | 41.66 | 7711 | Nostoc |
| PMC 881__Proteobacteria | 1146 | 27,227,936 | 67.65 | 25,649 | Proteobacteria |
| PMC 882__Armatimonadetes | 5 | 7,360,037 | 54.92 | 6711 | Armatimonadetes |
| PMC 882__Cyanobacteria | 46 | 8,132,153 | 41.34 | 7582 | Trichormus |
| PMC 882__Proteobacteria | 41 | 9,857,661 | 66.03 | 9926 | Proteobacteria |
| PMC 883__Armatimonadetes | 4 | 3,575,512 | 55.83 | 3399 | Fimbriimonadales |
| PMC 883__Chloroflexi | 6 | 5,641,484 | 58.03 | 5032 | Chloroflexi |
| PMC 883__Cyanobacteria | 5 | 6,694,777 | 46.94 | 6313 | Leptolyngbya |
| PMC 883__Proteobacteria | 72 | 6,645,423 | 67.63 | 6660 | Proteobacteria |
| PMC 884__Bacteroidetes | 4 | 2,942,276 | 39.52 | 3054 | Bacteroidetes |
| PMC 884__Cyanobacteria | 46 | 13,243,152 | 38.60 | 11,176 | Calothrix |
| PMC 884__Proteobacteria | 132 | 11,961,935 | 68.16 | 12,136 | Proteobacteria |
| PMC 885__Bacteroidetes | 30 | 7,094,005 | 39.52 | 6347 | Flavobacterium |
| PMC 885__Cyanobacteria | 137 | 5,858,006 | 35.29 | 5328 | Chroococcus |
| PMC 885__Proteobacteria | 52 | 18,208,582 | 65.84 | 17,483 | Proteobacteria |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Halary, S.; Duperron, S.; Demay, J.; Duval, C.; Hamlaoui, S.; Piquet, B.; Reinhardt, A.; Bernard, C.; Marie, B. Metagenome-Based Exploration of Bacterial Communities Associated with Cyanobacteria Strains Isolated from Thermal Muds. Microorganisms 2022, 10, 2337. https://doi.org/10.3390/microorganisms10122337
Halary S, Duperron S, Demay J, Duval C, Hamlaoui S, Piquet B, Reinhardt A, Bernard C, Marie B. Metagenome-Based Exploration of Bacterial Communities Associated with Cyanobacteria Strains Isolated from Thermal Muds. Microorganisms. 2022; 10(12):2337. https://doi.org/10.3390/microorganisms10122337
Chicago/Turabian StyleHalary, Sébastien, Sébastien Duperron, Justine Demay, Charlotte Duval, Sahima Hamlaoui, Bérénice Piquet, Anita Reinhardt, Cécile Bernard, and Benjamin Marie. 2022. "Metagenome-Based Exploration of Bacterial Communities Associated with Cyanobacteria Strains Isolated from Thermal Muds" Microorganisms 10, no. 12: 2337. https://doi.org/10.3390/microorganisms10122337