
Theileria terrestris nov. sp.: A Novel Theileria in Lowland Tapirs (Tapirus terrestris) from Two Different Biomes in Brazil
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. DNA Extraction and PCR Protocols for Mammals’ Endogenous Genes
2.3. Conventional and Nested PCR Protocols for Piroplasmida Detection and Molecular Characterization
2.4. Sequencing, Sequence Analysis and Phylogeny
3. Results
3.1. PCR for Endogenous Mammal’s Genes
3.2. Screening Nested PCR for Piroplasmida Partial 18S rRNA Gene (~800 bp)
3.3. Amplification of Molecular Markers for Additional Molecular Characterization of Piroplasmida
3.4. Amplification of Extended Sequences from the Piroplasmida 18S rRNA Gene (~1500 bp)
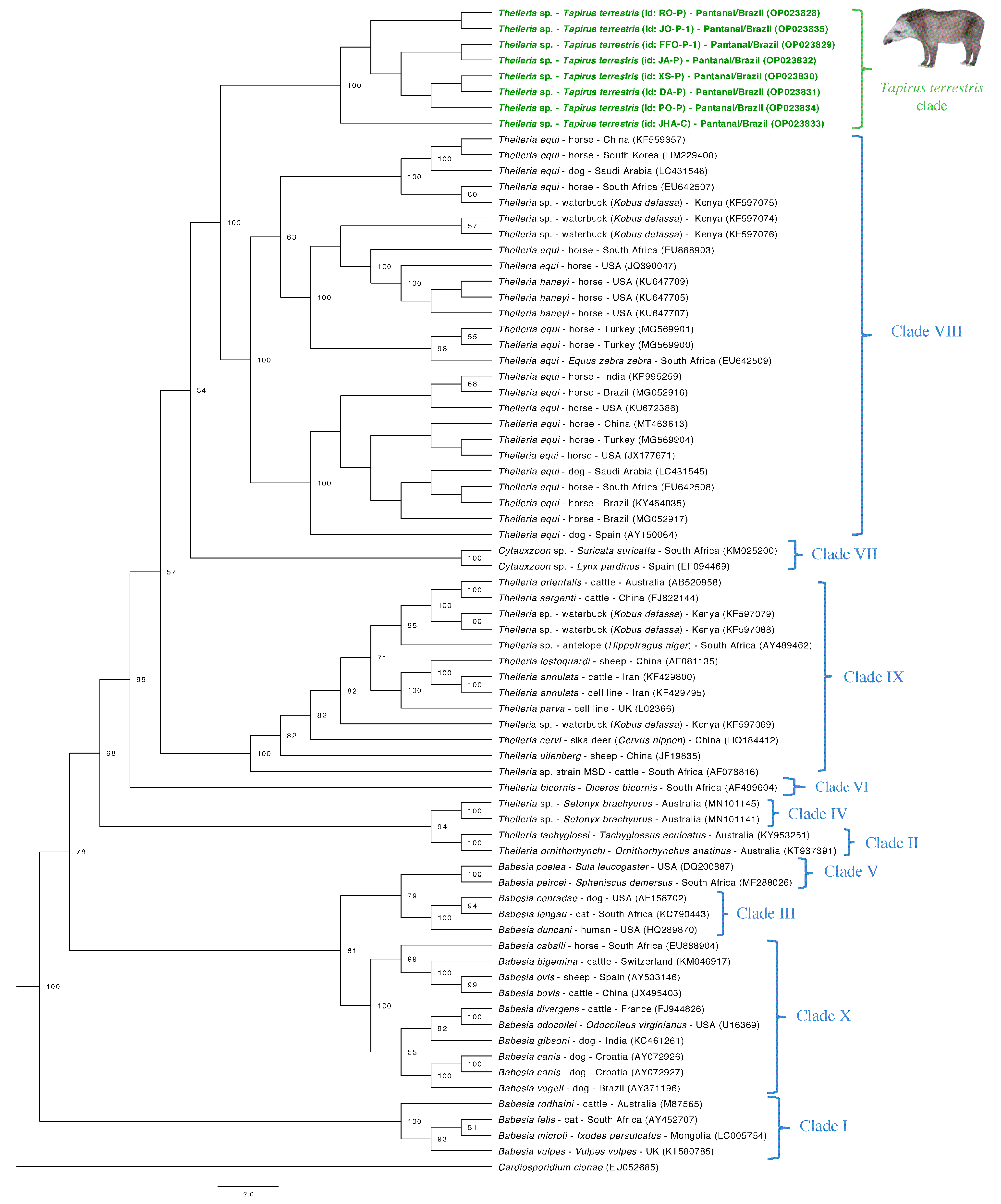
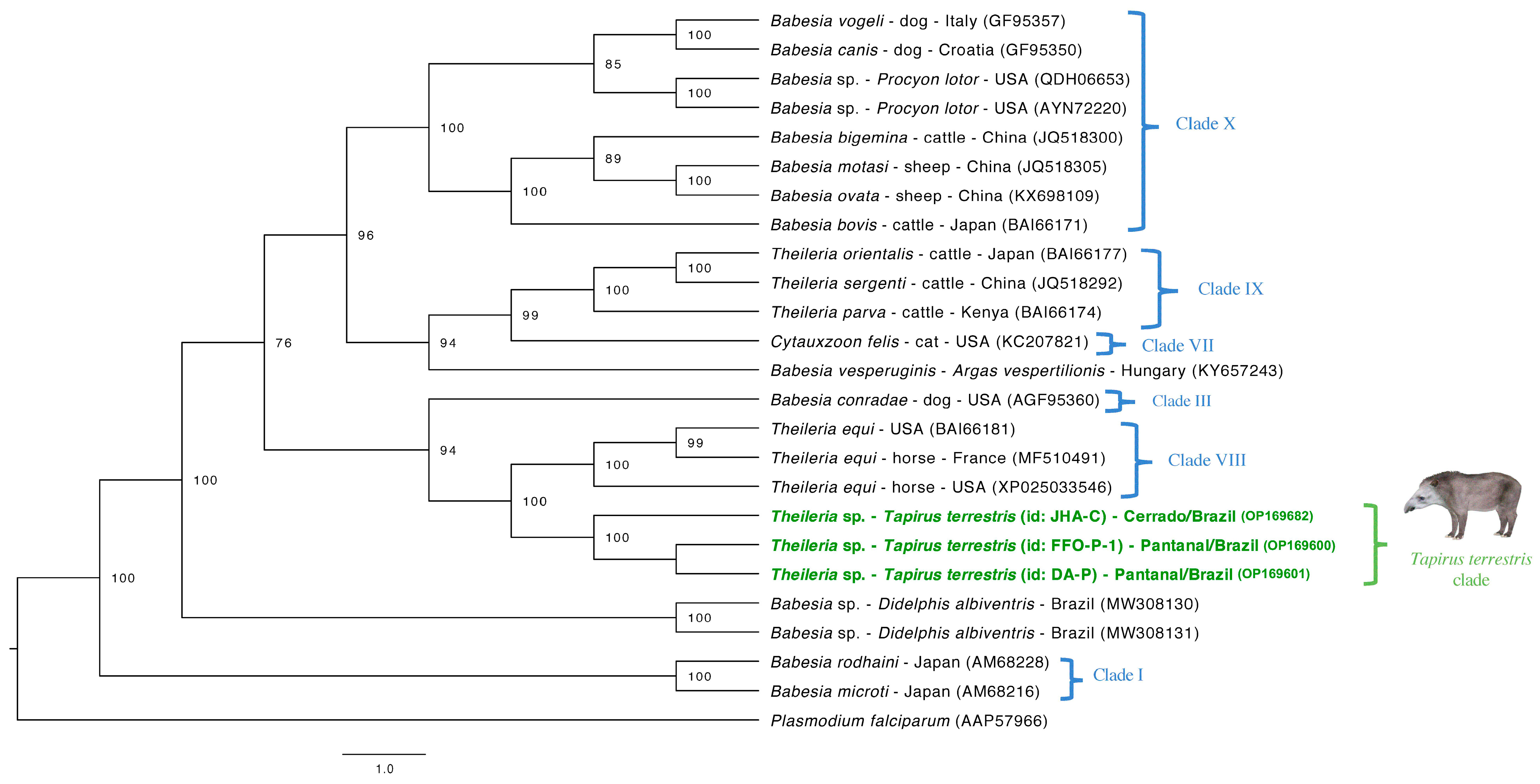
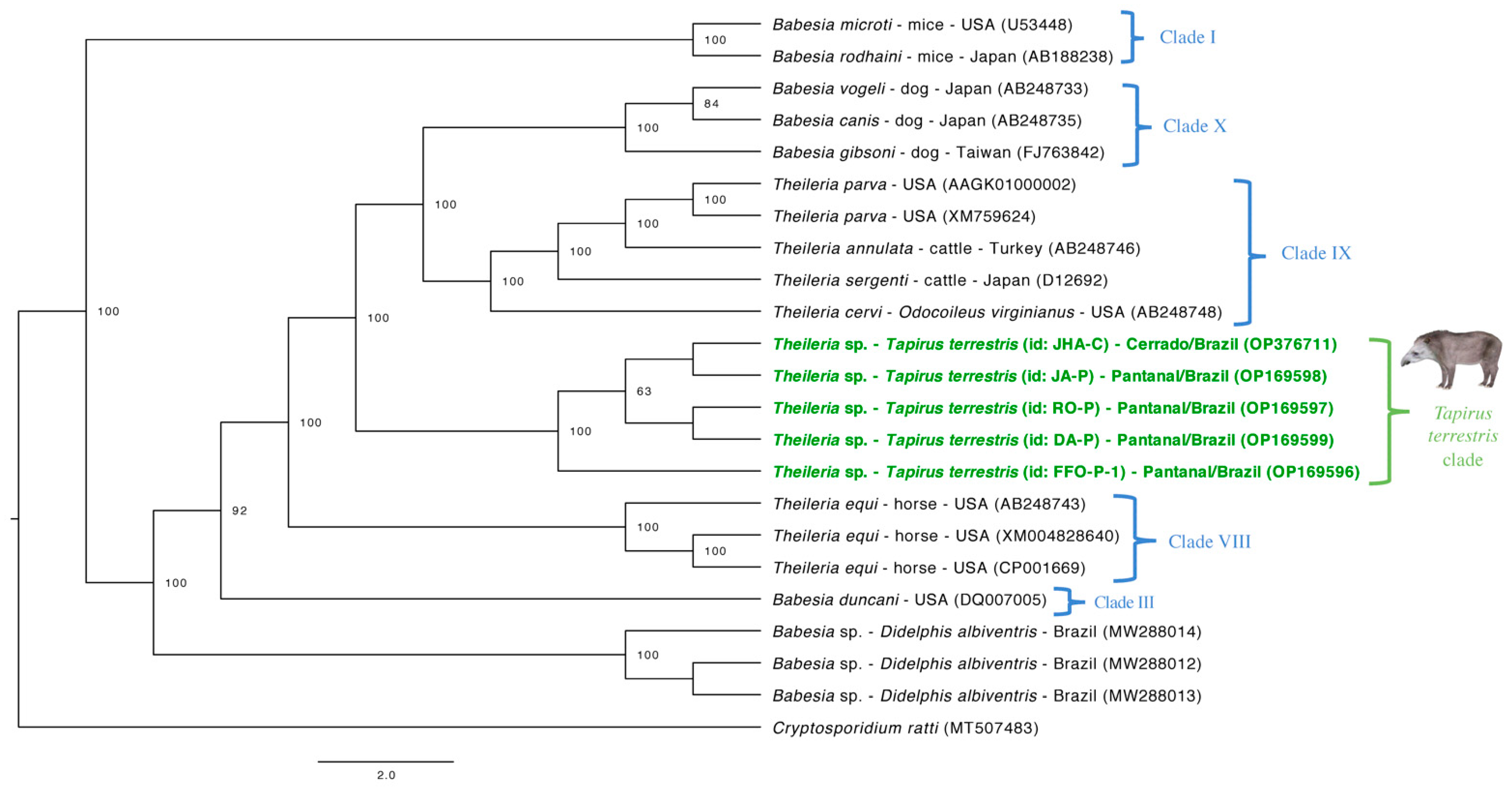
3.5. Phylogenetic Analyses
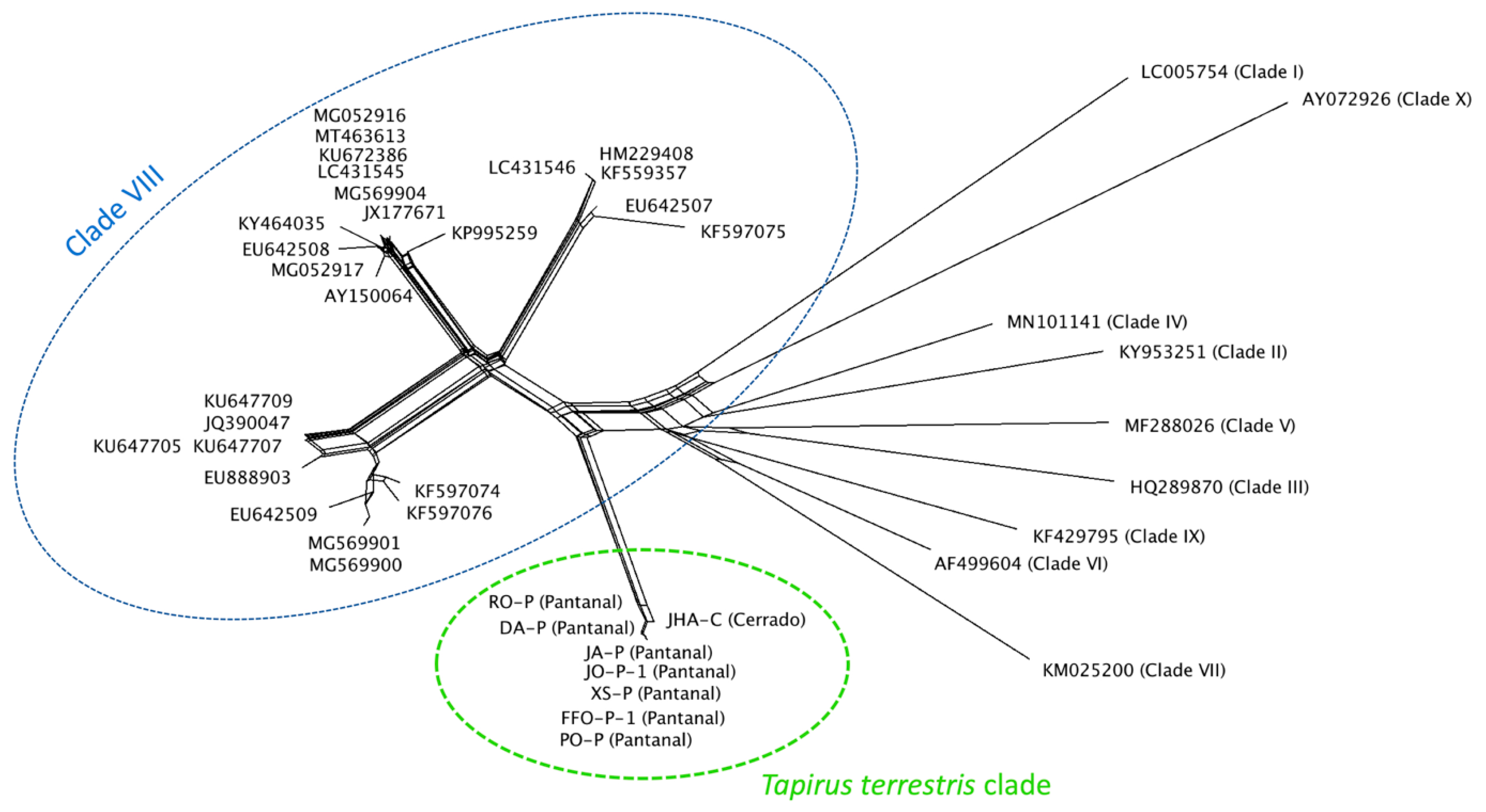
3.6. Distance Analysis by Splistree Software
3.7. Blood Smears Analysis
3.8. Description of a New Species: Theileria terrestris nov. sp.
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schnittger, L.; Ganzinelli, S.; Bhoora, R.; Omondi, D.; Nijhof, A.M.; Florin-Christensen, M. The Piroplasm Babesia, Cytauxzoon, and Theileria in farm and companion animals: Species compilation, molecular phylogeny, and evolutionary inshights. Parasitol. Res. 2022, 121, 1207–1245. [Google Scholar] [CrossRef] [PubMed]
- Acosta, I.C.L.; Costa, A.P.; Nunes, P.H.; Gondim, M.F.N.; Gatti, A.; Rossi, J.L., Jr.; Gennari, S.M.; Marcili, A. Morphological and molecular characterization and phylogenetic relationship of a new species of trypanosome in Tapirus terrestris (lowland tapir), Trypanosoma terrestris sp. nov., from Atlantic Rainforest of southeastern Brazil. Parasit. Vectors 2013, 6, 349. [Google Scholar] [CrossRef] [PubMed]
- Mongruel, A.C.B.; Medici, E.P.; Canena, A.D.C.; Calchi, A.C.; Machado, R.Z.; André, M.R. Expanding the Universe of Hemoplasmas: Multi-Locus Sequencing Reveals Putative Novel Hemoplasmas in Lowland Tapirs (Tapirus terrestris), the Largest Land Mammals in Brazil. Microorganisms 2022, 10, 614. [Google Scholar] [CrossRef]
- Silveira, A.W.; Oliveira, G.G.; Santos, L.M.; Azuaga, L.B.S.; Coutinho, C.R.M.; Echeverria, J.T.; Antunes, T.R.; Ramos, C.A.N.; Souza, A.I. Natural Infection of the South American Tapir (Tapirus terrestris) by Theileria equi. J. Wildl. Dis. 2017, 53, 411–413. [Google Scholar] [CrossRef]
- Gonçalves, T.S.; Barros, F.N.L.; Inoue, L.S.; Farias, D.M.; Lima, J.S.; Nobre, A.V.; Aidar, E.S.A.; Diniz, R.F.R.; Gering, A.P.; Scofield, A. Natural Theileria equi infection in captive Tapirus terrestris (Perissodactyla: Tapiridae) in the Brazilian Amazon. Ticks Tick-Borne Dis. 2020, 11, 101452. [Google Scholar] [CrossRef] [PubMed]
- Scoles, G.A.; Ueti, M.W. Vector Ecology of Equine Piroplasmosis. Annu. Rev. Entomol. 2015, 7, 561–580. [Google Scholar] [CrossRef] [PubMed]
- Santos, T.M.; Roier, E.C.R.; Pires, M.S.; Santos, H.A.; Vilela, J.A.R.; Peckle, M.; Paulino, P.G.; Baldani, C.D.; Massard, C.L. Molecular evidence of Anaplasma phagocytophilum and Theileria equi coinfection in horses from Rio de Janeiro, Brazil. Vet. Anim. Sci. 2019, 20, 100055. [Google Scholar] [CrossRef]
- Campos, J.B.V.; André, M.R.; Gonçalves, L.R.; Freschi, C.R.; Santos, F.M.; Oliveira, C.E.; Piranda, E.M.; Andrade, G.B.; Macedo, G.C.; Machado, R.Z.; et al. Assessment of equine piroplasmids in the Nhecolândia sub-region of Brazilian Pantanal wetland using serological, parasitological, molecular, and hematological approaches. Ticks Tick-Borne Dis. 2019, 10, 714–721. [Google Scholar] [CrossRef]
- Ribeiro, M.F.; da Silveira, J.A.; Bastos, C.V. Failure of the Amblyomma cajennense nymph to become infected by Theileria equi after feeding on acute or chronically infected horses. Exp. Parasitol. 2011, 28, 324–327. [Google Scholar] [CrossRef] [Green Version]
- Labruna, M.B.; Martins, T.F.; Acosta, I.C.L.; Serpa, M.C.A.; Soares, H.S.; Teixeira, R.H.F.; Fernandes-Santos, R.C.; Medici, E.P. Ticks and rickettsial exposure in lowland tapirs (Tapirus terrestris) of three Brazilian biomes. Ticks Tick Borne Dis. 2021, 12, 101648. [Google Scholar] [CrossRef] [PubMed]
- Birkenheuer, A.J.; Levy, M.G.; Breitschewerdt, E.B. Development and evaluation of a seminested PCR for detection and differentiation of Babesia gibsoni (Asian genotype) and B. canis DNA in canine blood samples. J. Clin. Microbiol. 2003, 41, 4172–4177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, E.C.; Contijo, C.M.; Cruz, I.; Melo, M.N.; Silva, A.M. Alternative PCR protocol using a single primer set for assessing DNA quality in several tissues from a large variety of mammalian species living in areas endemic for leishmaniasis. Mem. Inst. Oswaldo Cruz 2010, 105, 895–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jefferies, R.; Ryan, U.M.; Irwin, P.J. PCR–RFLP for the detection and differentiation of the canine piroplasm species and its use with filter paper-based technologies. Vet. Parasitol. 2007, 144, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Gou, H.; Guan, G.; Liu, A.; Ma, M.; Xu, Z.; Liu, Z.; Ren, Q.; Li, Y.; Yang, J.; Chen, Z.; et al. A DNA barcode for Piroplasmea. Acta Trop. 2012, 124, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Corduneanu, A.; Hrazdilov, K.; Sandor, A.D.; Matei, I.A.; Ionică, A.M.; Barti, L.; Ciocănău, M.A.; Măntoiu, D.S.; Coroiu, I.; Hornok, S.; et al. Babesia vesperuginis, a neglected piroplasmid: New host and geographical records, and phylogenetic relations. Parasit. Vectors 2017, 10, 598–606. [Google Scholar] [CrossRef] [Green Version]
- Schreeg, M.E.; Marr, H.S.; Griffith, E.H.; Tarigo, J.L.; Bird, D.M.; Reichard, M.V.; Birkenheuer, A.J. PCR amplification of a multi-copy mitochondrial gene (cox3) improves detection of Cytauxzoon felis infection as compared to a ribosomal gene (18S). Vet. Parasitol. 2016, 225, 123–130. [Google Scholar] [CrossRef]
- Barbosa, A.D.; Austen, J.; Portas, T.J.; Friend, A.; Ahlstrom, L.A.; Oskam, C.L.; Ryan, U.M.; Irwin, P.J. Sequence analyses at mitochondrial and nuclear loci reveal a novel Theileria sp. and aid in the phylogenetic resolution of piroplasms from Australian marsupials and ticks. PLoS ONE 2019, 4, e0225822. [Google Scholar] [CrossRef] [Green Version]
- Soares, J.F.; Girotto, A.; Brand, P.E.; Da Silva, A.S.; França, R.T.; Lopes, S.T.A.; Labruna, M.B. Detection and molecular characterization of a canine piroplasm from Brazil. Vet. Parasitol. 2011, 180, 203–208. [Google Scholar] [CrossRef]
- Zamoto, A.; Tsuji, M.; Wei, Q.; Cho, S.H.; Shin, E.H.; Kim, T.S.; Leonova, G.N.; Hagiwara, K.; Asakawa, M.; Kariwa, H.; et al. Epizootiologic survey for Babesia microti among small wild mammals in northeastern Eurasia and a geographic diversity in the beta-tubulin gene sequences. J. Vet. Med. Sci. 2004, 66, 785–792. [Google Scholar] [CrossRef] [Green Version]
- Shock, B.C.; Birkenheuer, A.J.; Patton, L.L.; Olfenbuttel, C.; Beringer, J.; Grove, D.M.; Peek, M.; Butfiloski, J.W.; Hughes, D.W.; Lockhart, J.M.; et al. Variation in the ITS-1 and ITS-2 rRNA genomic regions of Cytauxzoon felis from bobcats and pumas in the eastern United States and comparison with sequences from domestic cats. Vet. Parasitol. 2012, 190, 29–35. [Google Scholar] [CrossRef]
- Nicolaiewsky, T.B.; Richter, M.F.; Lunge, V.R.; Cunha, C.W.; Delagostin, O.; Ikuta, N.; Fonseca, A.S.; Da Silva, S.S.; Ozaki, L.S. Detection of Babesia equi (Laveran, 1901) by nested polymerase chain reaction. Vet. Parasitol. 2001, 101, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Kawabuchi, T.; Tsuji, M.; Sado, A.; Matoba, Y.; Asakawa, M.; Ishihara, C. Babesia microti-like parasites detected in feral raccoons (Procyon lotor) captured in Hokkaido, Japan. J. Vet. Med. Sci. 2005, 67, 825–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oosthuizen, M.C.; Zweygarth, E.; Collins, N.E.; Troskie, M.; Penzhorn, B.L. Identification of a Novel Babesia sp. from a Sable Antelope (Hippotragus niger Harris, 1838). J. Clin. Microbiol. 2008, 46, 2247–2251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greay, T.L.; Zahedi, A.; Krige, A.S.; Owens, J.M.; Rees, R.L.; Ryan, U.M.; Oskam, C.L.; Irwin, P.J. Endemic, exotic and novel apicomplexan parasites detected during a national study of ticks from companion animals in Australia. Parasit. Vectors 2018, 11, 197. [Google Scholar] [CrossRef] [Green Version]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef] [Green Version]
- Hirata, H.; Kawai, S.; Maeda, M.; Jinnai, M.; Fujisawa, K.; Katakai, Y.; Hikosaka, K.; Tanabe, K.; Yasutomi, Y.; Ishihara, C. Identification and phylogenetic analysis of Japanese Macaque Babesia-1 (JM-1) detected from a Japanese Macaque (Macaca fuscata fuscata). Am. J. Trop. Med. Hyg. 2011, 85, 635–638. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Sela, I.; Ashkenazy, H.; Katoh, K.; Pupko, T. GUIDANCE2: Accurate detection of unreliable alignment regions accounting for the uncertainty of multiple parameters. Nucleic Acids Res. 2015, 43, W7–W14. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. ModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Jalovecka, M.; Sojka, D.; Ascencio, M.; Schinittger, L. Babesia Life Cycle—When Phylogeny Meets Biology. Trends Parasitol. 2019, 35, 365–368. [Google Scholar] [CrossRef] [PubMed]
- Silva, V.L.B.; Almeida, S.L.H.; Maia, M.O.; Santos, T.A.; Pavelegini, L.A.D.; Zaffalon, G.B.; Marcili, A.; Morgado, T.O.; Dutra, V.; Nakazato, L.; et al. Post mortem protozoan hemoparasites detection in wild mammals from Mato Grosso state. Rev. Bras. Parasitol. Vet. 2021, 30, e013021. [Google Scholar] [CrossRef]
- André, M.R.; Baccarim, N.C.D.; Sousa, K.C.M.; Gonçalves, L.R.; Henrique, P.C.; Ontivero, G.R.C.R.; Gonzalez, L.I.H.; Nery, C.C.V.; Chagas, F.C.R.; Monticelli, C.; et al. Arthropod-borne pathogens circulating in free-roaming domestic cats in a zoo environment in Brazil. Ticks Tick Borne Dis. 2014, 5, 545–551. [Google Scholar] [CrossRef] [PubMed]
- André, M.R.; Herrera, H.M.; Fernandes, S.J.; de Sousa, K.C.; Gonçalves, L.R.; Domingos, I.H.; de Macedo, G.C.; Machado, R.Z. Tick-borne agents in domesticated and stray cats from the city of Campo Grande, state of Mato Grosso do Sul, midwestern Brazil. Ticks Tick Borne Dis. 2015, 6, 779–786. [Google Scholar] [CrossRef]
- Bishop, R.P.; Kappmeyer, L.S.; Onzere, C.K.; Odongo, D.O.; Githaka, N.; Sears, K.P.; Knowles, D.P.; Fry, L.M. Equide infective Theileria cluster in distinct 18S rRNA gene clades comprinsing multiple taxa with unusually broad mammalian host ranges. Parasit. Vectors 2020, 13, 2020. [Google Scholar] [CrossRef]
- Inácio, E.L.; Pérez-Macchi, S.; Alabi, A.; Bittencourt, P.; Miller, A. Prevalence and molecular characterization of piroplasmids in domestic dogs from Paraguay. Ticks Tickborne Dis. 2019, 10, 321–327. [Google Scholar] [CrossRef]
- Azmi, K.; Al-Jawabreh, A.; Abdeen, Z. Molecular Detection of Theileria ovis and Theileria equi in Livestock from Palestine. Sci. Rep. 2019, 9, 11557. [Google Scholar] [CrossRef] [Green Version]
- Bhoora, R.V.; Collins, N.E.; Schnittger, L.; Troskie, C.; Marumo, R.; Labuschagne, K.; Smith, R.M.; Dalton, D.L.; Mbizeni, S. Molecular genotyping and epidemiology of equine piroplasmids in South Africa. Ticks Tick Borne Dis. 2020, 11, 101358. [Google Scholar] [CrossRef]
- Wu, S.; Xiong, J.; Yu, Y. Taxonomic resolutions based on 18S rRNA genes: A case study of subclass copepoda. PLoS ONE 2015, 10, e0131498. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, P.; Menezes, T.R.; Torres, J.M.; de Oliveira, C.E.; Lourenço, E.C.; Herrera, H.M.; Machado, R.Z.; André, M.R. First molecular detection of piroplasmids in non-hematophagous bats from Brazil, with evidence of putative novel species. Parasitol. Res. 2021, 120, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, L.R.; Paludo, G.; Bisol, T.B.; Perles, L.; de Oliveira, L.B.; de Oliveira, C.M.; da Silva, T.M.V.; Nantes, W.A.G.; Duarte, M.A.; Santos, F.M.; et al. Molecular detection of piroplasmids in synanthropic rodents, marsupials, and associated ticks from Brazil, with phylogenetic inference of a putative novel Babesia sp. from white-eared opossum (Didelphis albiventris). Parasitol. Res. 2021, 120, 3537–3546. [Google Scholar] [CrossRef] [PubMed]
- Hornok, S.; Corduneanu, A.; Kontschán, J.; Bekő, K.; Szőke, K.; Görföl, T.; Gyuranecz, M.; Sándor, A.D. Analyses of separate and concatenated cox1 and 18S rRNA gene sequences indicate that the bat piroplasm Babesia vesperuginis is phylogenetically close to Cytauxzoon felis and the ‘prototheilerid’ Babesia conradae. Acta Vet. Hung. 2018, 66, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Katzer, F.; Ngugi, D.; Walker, A.R.; McKeever, D.J. Genotypic diversity, a survival strategy for the apicomplexan parasite Theileria parva. Vet. Parasitol. 2010, 167, 236–243. [Google Scholar] [CrossRef] [Green Version]
- Al-Hamidhi, S.; Tageldin, H.M.; Weir, W.; Al-Fahdi, A.; Johnson, E.H.; Bobade, P.; Alqamashoui, B.; Beja-Pereira, A.; Thompson, J.; Kinnaird, J.; et al. Genetic Diversity and Population Structure of Theileria annulata in Oman. PLoS ONE 2015, 10, e0139581. [Google Scholar] [CrossRef] [Green Version]
- Vitari, G.L.V.; Costa, R.L.; Abreu, A.P.M.; Peckle, M.; Silva, C.B.; Paulino, P.G.; Pires, M.S.; Massard, C.L.; Santos, H.A. Genetic Diversity of Theileria equi From Horses in Different Regions of Brazil Based On the 18S rRNA Gene. J. Parasitol. 2019, 105, 186–194. [Google Scholar] [CrossRef]
- Hare, M.P. Prospects for nuclear gene phylogeography. Trends Ecol. Evol. 2001, 16, 700–706. [Google Scholar] [CrossRef]
- Yamasaki, M.; Inokuma, H.; Sugimoto, C.; Shaw, S.E.; Aktas, M.; Yabsley, M.J.; Yamato, O.; Maede, Y. Comparison and phylogenetic analysis of the heat shock protein 70 gene of Babesia parasites from dogs. Vet. Parasitol. 2007, 145, 217–227. [Google Scholar] [CrossRef]
- Yokoyama, N.; Bork, S.; Nishisaka, M.; Hirata, H.; Matsuo, T.; Inoue, N.; Xuan, X.; Suzuki, H.; Sugimoto, C.; Igarashi, I. Roles of the Maltese cross form in the development of parasitemia and protection against Babesia microti infection in mice. Infect. Immun. 2003, 71, 411–417. [Google Scholar] [CrossRef] [Green Version]
- Knowles, D.P.; Kappmeyer, L.S.; Haney, D.; Herndon, D.R.; Fry, L.M.; Munro, J.B.; Sears, K.; Ueti, M.W.; Wise, L.N.; Silva, M.; et al. Discovery of a novel species, Theileria haneyi n. sp., infective to equids, highlights exceptional genomic diversity within the genus Theileria: Implications for apicomplexan parasite surveillance. Int. J. Parasitol. 2018, 48, 679–690. [Google Scholar] [CrossRef] [PubMed]
- Fawcett, D.W.; Conrad, P.A.; Grootenhuis, J.G.; Morzaria, S.P. Ultrastructure of the intra-erythrocytic stage of Theileria species from cattle and waterbuck. Tissue Cell 1987, 19, 643–655. [Google Scholar] [CrossRef] [PubMed]
- Camacho, T. Roles of the Maltese Cross Form of Babesia microti in the development of parasitemia in B. microti infection. ASM Infect. Immun. 2004, 72, 4929–4930. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primers | Fragment Size (bp) | References |
|---|---|---|---|
| 18S rRNA | 1st reaction: BTF1 (5′-GGCTCATTACAACAGTTATAG-3′) and BTR21 (5′-CCCAAAGACTTTGATTTCTCTC-3′); 2nd reation: BTF2 (5′-CCGTGCTAATTGTAGGGCTAATAC-3′) and BTR2 (5′-GGACTACGACGGTATCTGATCG-3′). | ~800 | [13] |
| 18S rRNA | Nbab_1F (5′- AAGCCATGCATGTCTAAGTATAAGCTTTT-3′) and 18Sapir (5′-GGATCACTCGATCGGTAGGAG-3′) | ~1500 | [23,24] |
| 18S rRNA | 1st reaction: Piro0F (5′-GCCAGTAGTCATATGCTTGTGTTA-3′) and Piro6R (5′-CTCCTTCCTYTAAGTGATAAGGTTCAC-3′); 2nd reaction: Piro1F (5′-CCATGCATGTCTWAGTAYAARCTTTTA-3′) and Piro5.5R (5′-CCTYTAAGTGATAAGGTTCACAAAACTT-3′) | ~1500 | [22] |
| cox-1 | 1st reaction: Bab_for1 (5′-ATWGGATTYTATATGAGTAT-3′) and Bab_Rev1 (5′-ATAATCWGGWATYCTCCTTGG-3′), Bab_for2; 2nd reation: (5′-TCTCTWCATGGWTTAATTATGATAT-3′) and Bab_Rev2 (5′-TAGCTCCAATTGAHARWACAAAGTG-3′) | ~924 | [14,15] |
| cox-3 | COX3F (5′-ACTGTCAGCTAAAACGTATC-3′) and COX3R (5′-ACAGGATTAGATACCCTGG-3′) | ~600 | [16,17] |
| hsp70 | hsp70F1 (5′-CATGAAGCACTGGCCHTTCAA-3′) and hsp70R2 (5′-GBAGGTTGTTGTCCTTVGTCAT-3) | ~740 | [18] |
| cytb | cytbF (5′-TTAGTGAAGGAACTTGACAGGT-3′) and cytbR (5′-CGGTTAATCTTTCCTATTCCTTACG-3′) | ~1000 | [16,17] |
| β-tubulin | Tubu-63F (5′-CAAATWGGYGCMAARTTYTGGGA-3′) and Tubu-3F (5′-TCGTCCATACCTTCWCCSGTRTACCAGTG-3′) | ~1200 | [19] |
| ITS-1 | 1st reaction: ITS15C (5′-CGATCGAGTGATCCGGTGAATTA-3′) and ITS13B (5′-GCTGCGTCCTTCATCGTTGTG-3′); 2nd reaction: (5′-AAGGAAGGAGAAGTCGTAACAAGG-3′) and ITS15C (5′-TTGTGTGAGCCAAGACATCCA-3′) | ~450 | [20] |
| ema-1 | 1st reaction: EMAE-F (5′-CCGCCCTTCACCTCGTTCTCAA-3′) and EMAE-R (5′-TCTCGGCGGCATCCTTGACCTC-3′); 2nd reaction: EMAI-F (5′-CCGTCTCCGTTGACTTGGCCG-3′) and EMAIR (5′-GGACGCGCTTGCCTGGAGCCT-3′) | ~396 | [21] |
| Primers | Primers | Annealing Temperature | Primers Reference |
|---|---|---|---|
| Pair 1 | Piro1F (5′-CCA TGC ATG TCT WAG TAY AAR CTT TTA-3′) and Piro5.5R (5′-CCT YTA AGT GAT AAG GTT CAC AAA ACT T-3′) | 59 °C | [26] |
| Pair 2 | BabF2 (5′-CCG TGC TAA TTG TAG GGC TAA TAC A-3′) and BabR2 (5′-GCT TGA AAC ACT CTA RTT TTC TCA A-3′) | 59 °C | [18] |
| Single 1 | Bab2F2 (5′-CTT TGA GAA ATT AGA GTG TTT-3′) | 59 °C | Present study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mongruel, A.C.B.; Medici, E.P.; da Costa Canena, A.; Calchi, A.C.; Perles, L.; Rodrigues, B.C.B.; Soares, J.F.; Machado, R.Z.; André, M.R. Theileria terrestris nov. sp.: A Novel Theileria in Lowland Tapirs (Tapirus terrestris) from Two Different Biomes in Brazil. Microorganisms 2022, 10, 2319. https://doi.org/10.3390/microorganisms10122319
Mongruel ACB, Medici EP, da Costa Canena A, Calchi AC, Perles L, Rodrigues BCB, Soares JF, Machado RZ, André MR. Theileria terrestris nov. sp.: A Novel Theileria in Lowland Tapirs (Tapirus terrestris) from Two Different Biomes in Brazil. Microorganisms. 2022; 10(12):2319. https://doi.org/10.3390/microorganisms10122319
Chicago/Turabian StyleMongruel, Anna Claudia Baumel, Emília Patrícia Medici, Ariel da Costa Canena, Ana Cláudia Calchi, Lívia Perles, Bianca Cardenal Balla Rodrigues, João Fabio Soares, Rosangela Zacarias Machado, and Marcos Rogério André. 2022. "Theileria terrestris nov. sp.: A Novel Theileria in Lowland Tapirs (Tapirus terrestris) from Two Different Biomes in Brazil" Microorganisms 10, no. 12: 2319. https://doi.org/10.3390/microorganisms10122319