
Are Reactive Oxygen Species (ROS) the Main Mechanism by Which Copper Ion Treatment Degrades the DNA of Mycobacterium avium subsp. paratuberculosis Suspended in Milk?
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. The MAP Strains and Inoculum Preparation
2.3. Reagents
2.4. Copper Ion Challenge
2.5. Selective Separation of MAP in Milk Samples
2.6. The MAP Total Quantification and Viability Assessment
2.7. Evaluation of the Production of Reactive Oxygen Intermediates (ROS)
2.8. Evaluation of Physicochemical Milk Properties
2.9. Assessing Copper Concentration in Milk
2.10. Statistical Analysis
3. Results
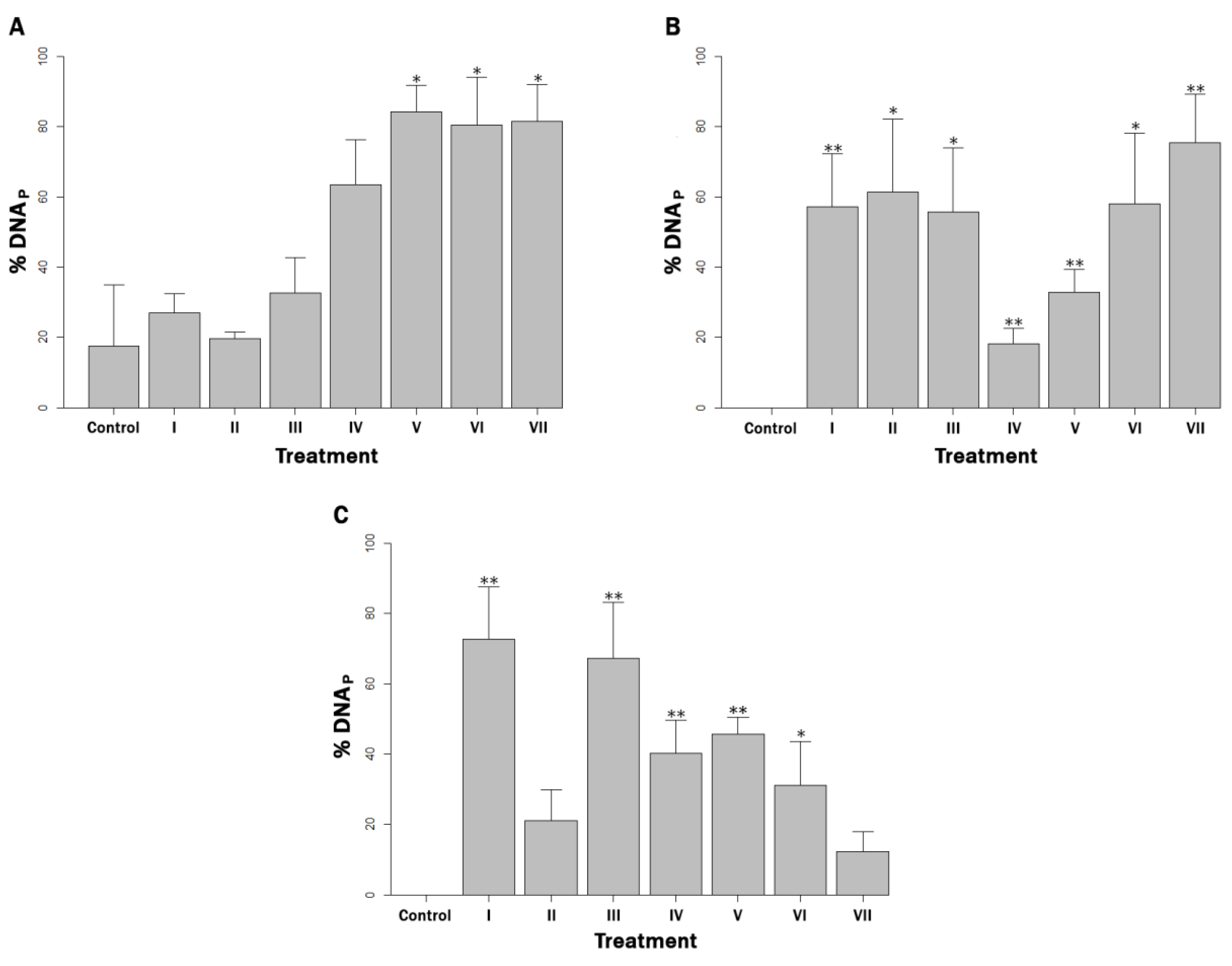
3.1. Estimation of Bacterial Load of Protected MAP Cells
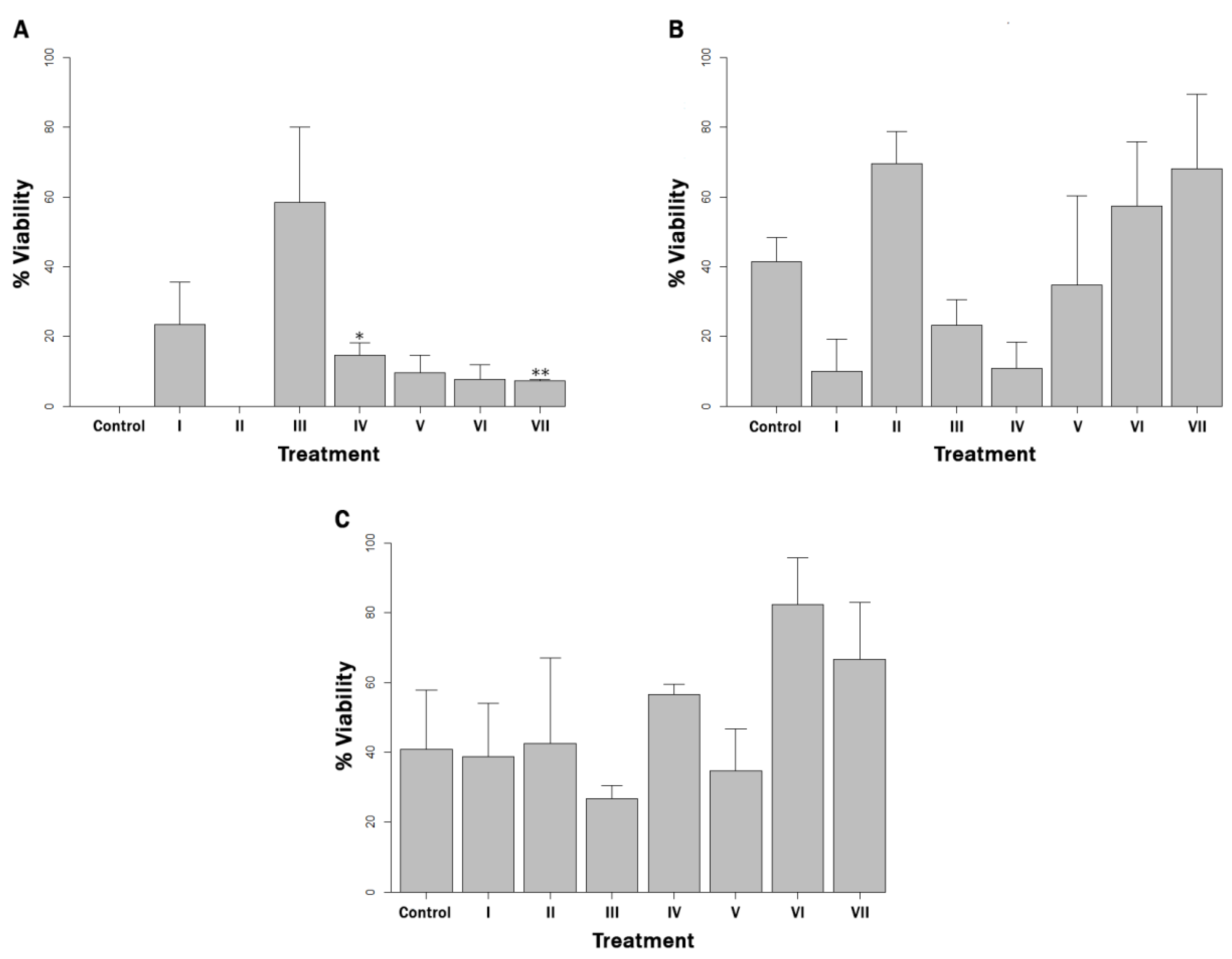
3.2. Estimation of Viable Bacterial Load
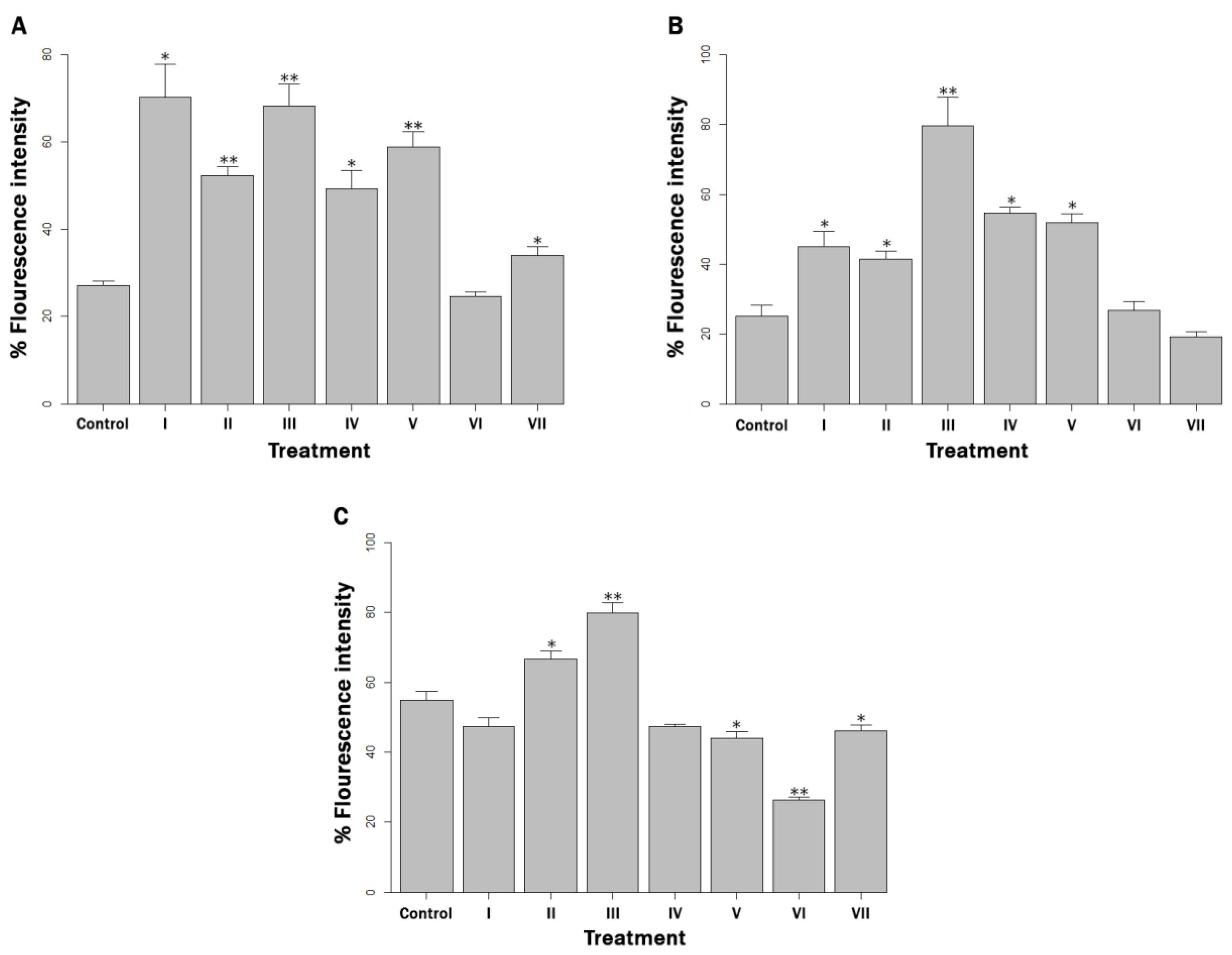
3.3. Reactive Oxygen Species Production
3.4. Physicochemical Changes in Milk Treated with Chelators/ROS Quenchers and Challenged with Copper
3.5. Determination of Copper Concentration in Milk
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fecteau, M.E. Paratuberculosis in cattle. Vet. Clin. Food Anim. Pract. 2018, 34, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, P.; Barkema, H.W.; Mason, S.; Beaulieu, E.; Hall, D.C. Economic losses due to Johne’s disease (paratuberculosis) in dairy cattle. J. Dairy Sci. 2021, 104, 3123–3143. [Google Scholar] [CrossRef] [PubMed]
- Eslami, M.; Shafiei, M.; Ghasemian, A.; Valizadeh, S.; Al-Marzoqi, A.H.; Shokouhi Mostafavi, S.K.; Nojoomi, F.; Mirforughi, S.A. Mycobacterium avium paratuberculosis and Mycobacterium avium complex and related subspecies as causative agents of zoonotic and occupational diseases. J. Cell Physiol. 2019, 234, 12415–12421. [Google Scholar] [CrossRef] [PubMed]
- Whittington, R.; Donat, K.; Weber, M.F.; Kelton, D.; Nielsen, S.S.; Eisenberg, S.; Arrigoni, N.; Juste, R.; Sáez, J.L.; Dhand, N.; et al. Control of paratuberculosis: Who, why and how. A review of 48 countries. BMC Vet. Res. 2019, 15, 198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steuer, P.; Collado, B.; Avilez, C.; Tejeda, C.; Soto, J.P.; Salgado, M. Is the transmission of Mycobacterium avium subspecies paratuberculosis (MAP) infection through milk intended to feed calves an overlooked item in paratuberculosis control programs? Trop. Anim. Health Prod. 2020, 52, 89–94. [Google Scholar] [CrossRef]
- Grant, I.R.; Ball, H.J.; Neill, S.D.; Rowe, M.T. Inactivation of Mycobacterium paratuberculosis in cows’ milk at pasteurization temperatures. Appl. Environ. Microbiol. 1996, 62, 631–636. [Google Scholar] [CrossRef] [Green Version]
- Grant, I.R.; Ball, H.J.; Rowe, M.T. Effect of high-temperature, short-time (HTST) pasteurization on milk containing low numbers of Mycobacterium paratuberculosis. Lett. Appl. Microbiol. 1998, 26, 166–170. [Google Scholar] [CrossRef]
- Salah, I.; Parkin, I.P.; Allan, E. Copper as an antimicrobial agent: Recent advances. RSC Adv. 2021, 11, 18179–18186. [Google Scholar] [CrossRef]
- Vincent, M.; Duval, R.E.; Hartemann, P.; Engels-Deutsch, M. Contact killing and antimicrobial properties of copper. J. Appl. Microbiol. 2018, 124, 1032–1046. [Google Scholar] [CrossRef] [Green Version]
- Mehtar, S.; Wiid, I.; Todorov, S.D. The antimicrobial activity of copper and copper alloys against nosocomial pathogens and Mycobacterium tuberculosis isolated from healthcare facilities in the Western Cape: An in-vitro study. J. Hosp. Infect. 2008, 68, 45–51. [Google Scholar] [CrossRef]
- Lin, Y.-S.E.; Vidic, R.D.; Stout, J.E.; McCartney, C.A.; Victor, L.Y. Inactivation of Mycobacterium avium by copper and silver ions. Water Res. 1998, 32, 1997–2000. [Google Scholar] [CrossRef]
- Steuer, P.; Avilez, C.; Tejeda, C.; González, N.; Ramirez-Reveco, A.; Ulloa, F.; Mella, A.; Grant, I.; Collins, M.T.; Salgado, M. In vitro inactivation of Mycobacterium avium subsp. paratuberculosis (MAP) by use of copper ions. BMC Microbiol. 2018, 18, 172. [Google Scholar] [CrossRef] [PubMed]
- Steuer, P.; Tejeda, C.; Martinez, O.; Ramirez-Reveco, A.; González, N.; Grant, I.R.; Foddai, A.C.G.; Collins, M.T.; Salgado, M. Effectiveness of copper ions against Mycobacterium avium subsp. paratuberculosis and bacterial communities in naturally contaminated raw cow’s milk. J. Appl. Microbiol. 2021, 131, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Lemire, J.A.; Harrison, J.J.; Turner, R.J. Antimicrobial activity of metals: Mechanisms, molecular targets and applications. Nat. Rev. Microbiol. 2013, 11, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, A.K.; Chakraborty, R.; Basu, T. Mechanism of antibacterial activity of copper nanoparticles. Nanotechnology 2014, 25, 135101. [Google Scholar] [CrossRef]
- Solioz, M. Copper and Bacteria: Evolution, Homeostasis and Toxicity, 1st ed.; Springer: Cham, Switzerland, 2018; pp. 49–80. [Google Scholar]
- Yang, B.; Chen, Y.; Shi, J. Reactive oxygen species (ROS)-based nanomedicine. Chem. Rev. 2019, 119, 4881–4985. [Google Scholar] [CrossRef] [PubMed]
- Tejeda, C.; Villegas, M.; Steuer, P.; Iranzo, E.; Gonzalez, N.; Ramirez-Reveco, A.; Salgado, M. Understanding the antibacterial mechanisms of copper ion treatment on Mycobacterium avium subsp. Paratuberculosis. Vet. Microbiol. 2022, 268, 109412. [Google Scholar] [CrossRef]
- Tejeda, C.; Steuer, P.; Villegas, M.; Reyes-Jara, A.; Iranzo, E.C.; Umaña, R.; Salgado, M. More Insights about the Efficacy of Copper Ion Treatment on Mycobacterium avium subsp paratuberculosis (MAP): A Clue for the Observed Tolerance. Pathogens 2022, 11, 272. [Google Scholar] [CrossRef]
- Shin, S.J.; Han, J.H.; Manning, E.J.; Collins, M.T. Rapid and reliable method for quantification of Mycobacterium paratuberculosis by use of the BACTEC MGIT 960 system. J. Clin. Microbiol. 2007, 45, 1941–1948. [Google Scholar] [CrossRef] [Green Version]
- Dzieciol, M.; Volgger, P.; Khol, J.; Baumgartner, W.; Wagner, M.; Hein, I. A novel real-time PCR assay for specific detection and quantification of Mycobacterium avium subsp. paratuberculosis in milk with the inherent possibility of differentiation between viable and dead cells. BMC Res. Notes 2010, 3, 1–8. [Google Scholar] [CrossRef]
- Foddai, A.; Elliott, C.T.; Grant, I.R. Maximizing capture efficiency and specificity of magnetic separation for Mycobacterium avium subsp. paratuberculosis cells. Appl. Environ. Microbiol. 2010, 76, 7550–7558. [Google Scholar] [CrossRef] [Green Version]
- Salgado, M.; Alfaro, M.; Salazar, F.; Badilla, X.; Troncoso, E.; Zambrano, A.; Gonzalez, M.; Mitchell, R.M.; Collins, M.T. Application of cattle slurry containing Mycobacterium avium subsp. paratuberculosis (MAP) to grassland soil and its effect on the relationship between MAP and free-living amoeba. Vet. Microbiol. 2015, 175, 26–34. [Google Scholar] [CrossRef]
- Foddai, A.C.; Grant, I.R. A novel one-day phage-based test for rapid detection and enumeration of viable Mycobacterium avium subsp. paratuberculosis in cows’ milk. Appl. Microbiol. Biotechnol. 2020, 104, 9399–9412. [Google Scholar] [CrossRef]
- Choi, O.; Hu, Z. Size dependent and reactive oxygen species related nanosilver toxicity to nitrifying bacteria. Environ. Sci. Technol. 2008, 42, 4583–4588. [Google Scholar] [CrossRef]
- Jorhem, L.; Afthan, G.; Cumont, G.; Dypdahl, H.P.; Gadd, K.; Havre, G.N.; Julshamn, K.; Kåverud, K.; Lind, B.; Loimaranta, J.; et al. Determination of Metals in Foods by Atomic Absorption Spectrometry after Dry Ashing: NMKL1 Collaborative Study. J. AOAC Int. 2000, 83, 1204–1211. [Google Scholar] [CrossRef] [Green Version]
- Warnes, S.L.; Keevil, C.W. Mechanism of copper surface toxicity in vancomycin-resistant enterococci following wet or dry surface contact. Appl. Environ. Microbiol. 2011, 77, 6049–6059. [Google Scholar] [CrossRef] [Green Version]
- Warnes, S.L.; Keevil, C.W. Lack of involvement of Fenton chemistry in death of methicillin-resistant and methicillin-sensitive strains of Staphylococcus aureus and destruction of their genomes on wet or dry copper alloy surfaces. Appl. Environ. Microbiol. 2016, 82, 2132–2136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, Y.; Meng, Q.; Mu, W.; Zhang, T. Recent advances in the applications and biotechnological production of mannitol. J. Funct. Foods 2017, 36, 404–409. [Google Scholar] [CrossRef]
- Yuting, C.; Rongliang, Z.; Zhongjian, J.; Yong, J. Flavonoids as superoxide scavengers and antioxidants. Free Radic. Biol. Med. 1990, 9, 19–21. [Google Scholar] [CrossRef]
- Bai, R.; Yong, H.; Zhang, X.; Liu, J.; Liu, J. Structural characterization and protective effect of gallic acid grafted O-carboxymethyl chitosan against hydrogen peroxide-induced oxidative damage. Int. J. Biol. Macromol. 2020, 143, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Sroka, Z.; Cisowski, W. Hydrogen peroxide scavenging, antioxidant and anti-radical activity of some phenolic acids. Food Chem. Toxicol. 2003, 41, 753–758. [Google Scholar] [CrossRef]
- Apak, R.; Güçlü, K.; Demirata, B.; Özyürek, M.; Çelik, S.E.; Bektaşoğlu, B.; Berker, K.; Özyurt, D. Comparative evaluation of various total antioxidant capacity assays applied to phenolic compounds with the CUPRAC assay. Molecules 2007, 12, 1496–1547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apak, R.; Güclü, K.; Özyürek, M.; Celik, S.E. Mechanism of antioxidant capacity assays and the CUPRAC (cupric ion reducing antioxidant capacity) assay. Microchim. Acta 2008, 160, 413–419. [Google Scholar] [CrossRef]
- Donaghy, J.A.; Johnston, J.; Rowe, M.T. Detection of Mycobacterium avium ssp. paratuberculosis in cheese, milk powder and milk using IS900 and f57-based qPCR assays. J. Appl. Microbiol. 2011, 110, 479–489. [Google Scholar] [CrossRef]
- Ryman, V.E.; Packiriswamy, N.; Sordillo, L.M. Role of endothelial cells in bovine mammary gland health and disease. Anim. Health Res. Rev. 2015, 16, 135–149. [Google Scholar] [CrossRef]
- Lam, P.L.; Wong, R.M.; Lam, K.H.; Hung, L.K.; Wong, M.M.; Yung, L.H.; Ho, Y.W.; Wong, E.Y.; Hau, D.K.P.; Gambaru, R.; et al. The role of reactive oxygen species in the biological activity of antimicrobial agents: An updated mini review. Chem. Biol. Interact. 2020, 320, 109023. [Google Scholar] [CrossRef]
- Dryden, M. Reactive oxygen species: A novel antimicrobial. Int. J. Antimicrob. Agents 2018, 51, 299–303. [Google Scholar] [CrossRef]
- Circu, M.L.; Aw, T.Y. Reactive oxygen species, cellular redox systems, and apoptosis. Free Radic. Biol. Med. 2010, 48, 749–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rastogi, R.P.; Singh, S.P.; Häder, D.P.; Sinha, R.P. Detection of reactive oxygen species (ROS) by the oxidant-sensing probe 2′, 7′-dichlorodihydrofluorescein diacetate in the cyanobacterium Anabaena variabilis PCC 7937. Biochem. Biophys. 2010, 397, 603–607. [Google Scholar] [CrossRef]
- Vaishampayan, A.; Grohmann, E. Antimicrobials functioning through ros-mediated mechanisms: Current insights. Microorganisms 2021, 10, 61. [Google Scholar] [CrossRef]
- Cheng, J.H.; Lv, X.; Pan, Y.; Sun, D.W. Foodborne bacterial stress responses to exogenous reactive oxygen species (ROS) induced by cold plasma treatments. Trends Food Sci. Technol. 2020, 103, 239–247. [Google Scholar] [CrossRef]
- Basu, D.; Khare, G.; Singh, S.; Tyagi, A.; Khosla, S.; Mande, S.C. A novel nucleoid-associated protein of Mycobacterium tuberculosis is a sequence homolog of GroEL. Nucleic. Acids Res. 2009, 37, 4944–4954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weigoldt, M.; Meens, J.; Bange, F.C.; Pich, A.; Gerlach, G.F.; Goethe, R. Metabolic adaptation of Mycobacterium avium subsp. paratuberculosis to the gut environment. Microbiology 2013, 159, 380–391. [Google Scholar] [CrossRef] [Green Version]
- Argüello, J.M.; Raimunda, D.; Padilla-Benavides, T. Mechanisms of copper homeostasis in bacteria. Front. Cell Infect. Microbio. 2013, 3, 73. [Google Scholar] [CrossRef] [Green Version]
- Khan, I.T.; Nadeem, M.; Imran, M.; Ullah, R.; Ajmal, M.; Jaspal, M.H. Antioxidant properties of Milk and dairy products: A comprehensive review of the current knowledge. Lipids Health Dis. 2019, 18, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro, V.M.R.; da Mota Silva, M.; Prudêncio de Souza, E.R.; Guerra, A.F.; Riger, C.J.; Laureano-Melo, R.; Luchese, R.H. Role of milk and honey in the tolerance of lactobacilli to oxidative stress. Braz. J. Microbiol. 2021, 52, 883–893. [Google Scholar] [CrossRef]
- Cervato, R.C.; Benvenuto, G. Studies on the antioxidant activity of milk caseins. Int. J. Food Sci. Nutr. 1999, 50, 291–296. [Google Scholar] [PubMed]
- Steuer, P.; Tejeda, C.; Moroni, M.; Verdugo, C.; Collins, M.T.; Salgado, M. Attempted Control of Paratuberculosis in Dairy Calves by Only Changing the Quality of Milk Fed to Calves. Animals 2021, 11, 2569. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatment | pH | EC | [O2] | |||
|---|---|---|---|---|---|---|
| No Cu | with Cu | No Cu | with Cu | No Cu | with Cu | |
| (Mean ± SD) | (Mean ± SD) | (Mean ± SD) | (Mean ± SD) | (Mean ± SD) | (Mean ± SD) | |
| EDTA | 5.3 ± 0.0 | 9.2 ± 0.1 | 7.1 ± 0.0 | 6.3 ± 0.1 | 8.5 ± 0.1 | 3.1 ± 0.2 |
| BCS | 6.6 ± 0.0 | 9.7 ± 0.0 | 4.9 ± 0.1 | 4.0 ± 0.0 | 8.2 ± 0.0 | 2.6 ± 0.0 |
| EDTA + BCS | 5.3 ± 0.0 | 9.6 ± 0.1 | 6.5 ± 0.1 | 5.9 ± 0.1 | 8.0 ± 0.1 | 1.7 ± 0.3 |
| D-mannitol | 6.7 ± 0.0 | 9.5 ± 0.0 | 5.0 ± 0.0 | 4.1 ± 0.0 | 8.0 ± 0.1 | 3.8 ± 0.1 |
| gallic acid | 6.6 ± 0.0 | 9.3 ± 0.1 | 5.0 ± 0.1 | 4.1 ± 0.1 | 7.9 ± 0.0 | 4.6 ± 0.0 |
| quercetin | 6.8 ± 0.1 | 9.6 ± 0.0 | 4.9 ± 0.0 | 4.1 ± 0.0 | 9.0 ± 0.0 | 1.9 ± 0.2 |
| D-mannitol + gallic acid + quercetin | 6.7 ± 0.0 | 9.2 ± 0.0 | 4.9 ± 0.1 | 4.0 ± 0.0 | 7.7 ± 0.0 | 2.6 ± 0.0 |
| control (+) | 6.6 ± 0.1 | 9.7 ± 0.0 | 5.3 ± 0.3 | 4.2 ± 0.1 | 9.3 ± 0.2 | 5.5 ± 0.1 |
| Treatment | No Cu | with Cu |
|---|---|---|
| EDTA | 1.11 | 846.56 |
| BCS | 0.48 | 544.06 |
| EDTA + BCS | 0.77 | 1255.82 |
| D-mannitol | 0.50 | 278.28 |
| gallic acid | 0.19 | 378.47 |
| quercetin | 0.82 | 205.23 |
| D-mannitol + gallic acid + quercetin | 0.42 | 156.90 |
| control (+) | 1.47 | 624.98 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villegas, M.; Tejeda, C.; Umaña, R.; Iranzo, E.C.; Salgado, M. Are Reactive Oxygen Species (ROS) the Main Mechanism by Which Copper Ion Treatment Degrades the DNA of Mycobacterium avium subsp. paratuberculosis Suspended in Milk? Microorganisms 2022, 10, 2272. https://doi.org/10.3390/microorganisms10112272
Villegas M, Tejeda C, Umaña R, Iranzo EC, Salgado M. Are Reactive Oxygen Species (ROS) the Main Mechanism by Which Copper Ion Treatment Degrades the DNA of Mycobacterium avium subsp. paratuberculosis Suspended in Milk? Microorganisms. 2022; 10(11):2272. https://doi.org/10.3390/microorganisms10112272
Chicago/Turabian StyleVillegas, Marcela, Carlos Tejeda, Reydoret Umaña, Esperanza C. Iranzo, and Miguel Salgado. 2022. "Are Reactive Oxygen Species (ROS) the Main Mechanism by Which Copper Ion Treatment Degrades the DNA of Mycobacterium avium subsp. paratuberculosis Suspended in Milk?" Microorganisms 10, no. 11: 2272. https://doi.org/10.3390/microorganisms10112272