
Functional Gastrointestinal Disorders with Psychiatric Symptoms: Involvement of the Microbiome–Gut–Brain Axis in the Pathophysiology and Case Management

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
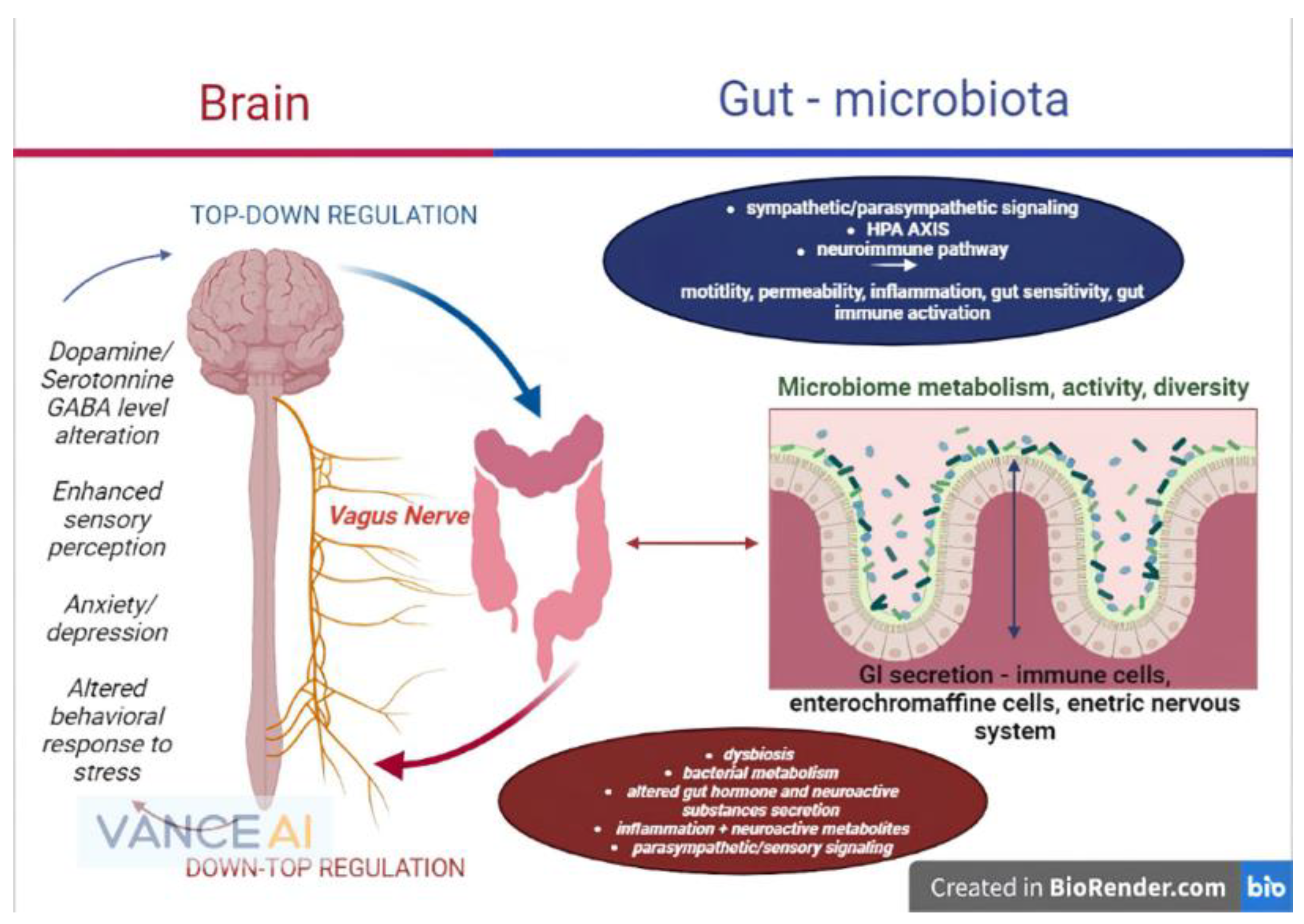
3. Review of Gut–Brain Axis and the Microbiome Physiology
3.1. The Microbiome
3.2. Brain to Gut Connection and Gut to Brain Connection
3.3. Functional Mechanisms and Pathways between Microbiome, Gut and Brain
3.3.1. The Autonomous Nervous System
3.3.2. The Enteric Nervous System
3.3.3. The Enteric Immune System
3.3.4. The Neuroendocrine System
3.4. The Hypothalamic–Pituitary–Adrenal Axis (HPA Axis)
3.5. Spinal Cord within the Gut–Brain Axis
3.6. The Neurotransmitters
3.7. Amino Acids and Microbiome
3.8. Microbial Metabolism of Short Chain Fatty Acids
4. Development and Importance of Microbiome–Gut–Brain Axis in Infants and Children
4.1. Infant Colic
4.2. Antibiotics in Functional Gastrointestinal Disorders as Modulator of Microbiome–Gut–Brain Axis in Infancy and Childhood
5. Functional Digestive Disorders—The Microbiome, the Enteric System and the Brain
5.1. Irritable Bowel Syndrome
5.2. Functional Abdominal Pain
5.3. Functional Abdominal Bloating
5.4. Functional Constipation
5.5. Functional Dyspepsia (FD)
6. Pharmacological Modulation of the Microbiome–Gut–Brain Axis
6.1. Antibiotics in Microbiome Management of FGIDs
6.2. Prebiotics, Probiotic or Psycho-Biotics?
6.2.1. Prebiotics
6.2.2. Probiotics

{kind=link}
| Bacterial Strain | Gastrointestinal Effects | Psychiatric Effects | Source |
|---|---|---|---|
| Lactobacillus casei | Anti-inflammatory effect, pain relief (children and adults) | Anxiety and depression symptom improvement, cognitive improvement | [129,130] |
| Lactobacillus acidophilus | Motility symptoms improvement, pain reduction, distension relief (children and adults) | Potential role in modulation of cannabinoid receptors | [131,132] |
| Lactobacillus brevis | Anti-inflammatory effect, motility symptoms improvement, intestinal barrier function improvement (children and adults) | Sleep, mood and affective symptoms imrpivement (important role in 1 GABA regulation) | [133,134] |
| Lactobacillus rhamnosus | Pain relief, anti-inflammatory effect (especially in children) (children and adults) | Cognitive improvement, mood and affective symptoms improvements (Dopamine, Glutamate, 2 5-Ht regulation) | [135,136] |
| Lactobacillus helveticus | Decrease in visceral hypersensitivity, pain relief, anti-inflammatory effect | Mood and behavior improvement (3 BDNF and 1 GABA regulation) | [137,138] |
| Lactococcus lactis | Under study—possible anti-inflammatory effect | Depression and anxiety score improvement (1 GABA regulation) | [139,140] |
| Saccharomyces boulardi | Inhibitory effect on inflammatory intestinal activity, reduces oxidative stress | Cognitive improvement | [141,142] |
| Bifidobacterium longum, lactis | Bloating relief, motility improvement, visceral pain relief, lactate production support via glycolysis from dietary fibers (+Firmicutes strain) | Reduces depression scores, improves cognitive and behavior symptoms (2 5-HT, 3 BDNF regulation) | [143,144,145] |
| Bacillus subtilis | Promotes bacterial diversity, improves intestinal barrier function, inflammation resolution, improves oxidative stress | Mood, cognitive and behavior improvement (2 5-HT regulation) | [146,147] |
| Akkermansia muciniphila | Inflammation resolution, intestinal barrier and enteral immune system improvement, digestive symptoms improvement | Important cognitive modulation, mood and behavior improvement | [148,149] |
6.3. Psychopharmacological Interventions in the Gut–Brain Axis
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Perlmutter, D. The Microbiome and the Brain. p. 233. Available online: https://www.perlego.com/book/1546614/the-microbiome-and-the-brain-pdf (accessed on 11 December 2021).
- Quigley, E. The gut-brain axis and the microbiome: Clues to pathophysiology and opportunities for novel management strategies in irritable bowel syndrome (IBS). J. Clin. Med. 2018, 7, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, K.V.A.; Burnet, P.W.J. Opposing effects of antibiotics and germ-free status on neuropeptide systems involved in social behaviour and pain regulation. BMC Neurosci. 2020, 21, 32. [Google Scholar] [CrossRef] [PubMed]
- Margolis, K.G.; Cryan, J.F.; Mayer, E.A. The microbiota-gut-brain axis: From motility to mood. Gastroenterology 2021, 160, 1486–1501. [Google Scholar] [CrossRef] [PubMed]
- Eguílaz, M.G.; Trapero, J.L.R.; Martínez, L.P.; Blanco, J.R. El eje microbiota-intestino-cerebro y sus grandes proyecciones. Rev. Neurol. 2019, 68, 111. [Google Scholar] [CrossRef]
- Usuda, H.; Okamoto, T.; Wada, K. Leaky gut: Effect of dietary fiber and fats on microbiome and intestinal barrier. Int. J. Mol. Sci. 2021, 22, 7613. [Google Scholar] [CrossRef] [PubMed]
- McVey Neufeld, K.A.; Mao, Y.K.; Bienenstock, J.; Foster, J.A.; Kunze, W.A. The microbiome is essential for normal gut intrinsic primary afferent neuron excitability in the mouse. Neurogastroenterol. Motil. 2013, 25, 183-e88. [Google Scholar] [CrossRef]
- Chen, Y.; Xu, J.; Chen, Y. Regulation of neurotransmitters by the gut microbiota and effects on cognition in neurological disorders. Nutrients 2021, 13, 2099. [Google Scholar] [CrossRef]
- Chen, R.; Xu, Y.; Wu, P.; Zhou, H.; Lasanajak, Y.; Fang, Y.; Tang, L.; Ye, L.; Li, X.; Cai, Z.; et al. Transplantation of fecal microbiota rich in short chain fatty acids and butyric acid treat cerebral ischemic stroke by regulating gut microbiota. Pharmacol. Res. 2019, 148, 104403. [Google Scholar] [CrossRef]
- Rao, M.; Gershon, M. The bowel and beyond: The enteric nervous system in neurological disorders. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 517–528. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Han, Y.; Du, J.; Liu, R.; Jin, K.; Yi, W. Microbiota-gut-brain axis and the central nervous system. Oncotarget 2017, 8, 32. [Google Scholar] [CrossRef]
- de Palma, G.; Collins, S.M.; Bercik, P.; Verdu, E.F. The microbiota-gut-brain axis in gastrointestinal disorders: Stressed bugs, stressed brain or both? J. Physiol. 2014, 592, 2989–2997. [Google Scholar] [CrossRef] [PubMed]
- Ford, A.C.; Lacy, B.E.; Harris, L.A.; Quigley, E.M.M.; Moayyedi, P. Effect of antidepressants and psychological therapies in irritable bowel syndrome. Am. J. Gastroenterol. 2019, 114, 1350–1365. [Google Scholar] [CrossRef] [PubMed]
- Cedeño, M.M.C.; Moreira, J.F.K.; Diaz, M.J.K.; Chávez, P.E.P.; Marquinez, S.P.M.; Falcones Espinoza, A.M.; Bravo Véliz, S.B.; Moreira Alava, R.A.; Ganchozo Mendoza, A.A. Use of antidepressant drugs in the treatment of irritable bowel syndrome. Arch. Venez. Farmacol. Ter. 2019, 38, 809–813. [Google Scholar]
- Messay, B.; Lim, A.; Marsland, A.L. Current understanding of the bi-directional relationship of major depression with inflammation. Biol. Mood Anxiety Disord. 2012, 2, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, S.M.; Surette, M.; Bercik, P. The interplay between the intestinal microbiota and the brain. Nat. Rev. Microbiol. 2012, 10, 735–742. [Google Scholar] [CrossRef]
- Keightley, P.C.; Koloski, N.A.; Talley, N.J. Pathways in gut-brain communication: Evidence for distinct gut-to-brain and brain-to-gut syndromes. Aust. N. Z. J. Psychiatry 2015, 49, 207–214. [Google Scholar] [CrossRef]
- Bonaz, B.; Bazin, T.; Pellissier, S. The vagus nerve at the interface of the microbiota-gut-brain axis. Front. Neurosci. 2018, 12, 49. [Google Scholar] [CrossRef] [Green Version]
- Goehler, L.E.; Gaykema, R.P.A.; Opitz, N.; Reddaway, R.; Badr, N.; Lyte, M. Activation in vagal afferents and central autonomic pathways: Early responses to intestinal infection with Campylobacter jejuni. Brain Behav. Immun. 2005, 19, 334–344. [Google Scholar] [CrossRef]
- Szabo, I.L.; Czimmer, J.; Mozsik, G. Cellular energetical actions of ‘chemical’ and ‘surgical’ vagotomy in gastrointestinal mucosal damage and protection: Similarities, differences and significance for brain-gut function. Curr. Neuropharmacol. 2016, 14, 901–913. [Google Scholar] [CrossRef] [Green Version]
- Perez-Burgos, A.; Wang, B.; Mao, Y.-K.; Mistry, B.; Neufeld, K.-A.M.; Bienenstock, J.; Kunze, W. Psychoactive bacteria Lactobacillus rhamnosus (JB-1) elicits rapid frequency facilitation in vagal afferents. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 304, G211–G220. [Google Scholar] [CrossRef] [Green Version]
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The gut-brain axis: Interactions between enteric microbiota, central and enteric nervous systems. Ann. Gastroenterol. 2015, 28, 203–209. [Google Scholar] [PubMed]
- Abdel-Haq, R.; Schlachetzki, J.C.M.; Glass, C.K.; Mazmanian, S.K. Microbiome–microglia connections via the gut–brain axis. J. Exp. Med. 2019, 216, 41–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, D.G.; Soto, R.; Yandamuri, S.; Stone, C.; Dickey, L.; Gomes-Neto, J.C.; Pastuzyn, E.D.; Bell, R.; Petersen, C.; Buhrke, K.; et al. The microbiota protects from viral- induced neurologic damage through microglia-intrinsic TLR signaling. eLife 2019, 8, e47117. [Google Scholar] [CrossRef] [PubMed]
- D’Mello, C.; Ronaghan, N.; Zaheer, R.; Dicay, M.; Le, T.; Macnaughton, W.K.; Surrette, M.G.; Swain, M.G. Probiotics improve inflammation-associated sickness behavior by altering communication between the peripheral immune system and the brain. J. Neurosci. 2015, 35, 10821–10830. [Google Scholar] [CrossRef] [Green Version]
- Gao, D.; Liu, Z.; Liu, F.; Chen, L.; Wang, W.; Ma, J.; Xu, C.; Jiang, Z.; Hou, J. Study of the immunoregulatory effect of Lactobacillus rhamnosus 1.0320 in immunosuppressed mice. J. Funct. Foods 2021, 79, 104423. [Google Scholar] [CrossRef]
- Cervantes-Barragan, L.; Chai, J.N.; Tianero, M.D.; Di Luccia, B.; Ahern, P.P.; Merriman, J.; Cortez, V.S.; Caparon, M.G.; Donia, M.S.; Gilfillan, S.; et al. Lactobacillus reuteri induces gut intraepithelial CD4+CD8αα+ T cells. Science 2017, 357, 806–810. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.J.; Emge, J.R.; Berzins, K.; Lung, L.; Khamishon, R.; Shah, P.; Rodrigues, D.M.; Sousa, A.J.; Reardon, C.; Sherman, P.; et al. Probiotics normalize the gut-brain-microbiota axis in immunodeficient mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 307, G793–G802. [Google Scholar] [CrossRef]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The Microbiota-Gut-Brain Axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef]
- Lund, M.L.; Egerod, K.L.; Engelstoft, M.S.; Dmytriyeva, O.; Theodorsson, E.; Patel, B.A.; Schwartz, T.W. Enterochromaffin 5-HT cells—A major target for GLP-1 and gut microbial metabolites. Mol. Metab. 2018, 11, 70–83. [Google Scholar] [CrossRef]
- Mandić, A.D.; Woting, A.; Jaenicke, T.; Sander, A.; Sabrowski, W.; Rolle-Kampcyk, U.; Von Bergen, M.; Blaut, M. Clostridium ramosum regulates enterochromaffin cell development and serotonin release. Sci. Rep. 2019, 9, 1177. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.J.; Hatoum, H.T.; Buchner, D.; Cox, D.; Balu, S. Impact of 5-HT 3 receptor antagonists on chemotherapy-induced nausea and vomiting: A retrospective cohort study. BMC Health Serv. Res. 2012, 12, 215. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.M.; Sun, E.W.; Keating, D.J. Mechanisms controlling hormone secretion in human gut and its relevance to metabolism. J. Endocrinol. 2020, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wren, A.M.; Bloom, S.R. Gut hormones and appetite control. Gastroenterology 2007, 132, 2116–2130. [Google Scholar] [CrossRef]
- Han, H.; Yi, B.; Zhong, R.; Wang, M.; Zhang, S.; Ma, J.; Yin, Y.; Yin, J.; Chen, L.; Zhang, H. From gut microbiota to host appetite: Gut microbiota-derived metabolites as key regulators. Microbiome 2021, 9, 162. [Google Scholar] [CrossRef] [PubMed]
- Jahnke, J.R.; Roach, J.; Azcarate-Peril, M.A.; Thompson, A.L. Maternal precarity and HPA axis functioning shape infant gut microbiota and HPA axis development in humans. PLoS ONE 2021, 16, e0251782. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, K.; Cowen, P.; Harmer, C.J.; Tzortzis, G.; Burnet, P.W.J.P. 3.023 Prebiotic intake reduces the waking cortisol response and alters emotional bias in healthy volunteers. Eur. Neuropsychopharmacol. 2014, 24, S74. [Google Scholar] [CrossRef]
- De Herdt, V.; Puimege, L.; De Waele, J.; Raedt, R.; Wyckhuys, T.; El Tahry, R.; Libert, C.; Wadman, W.; Boon, P.; Vonck, K. Increased rat serum corticosterone suggests immunomodulation by stimulation of the vagal nerve. J. Neuroimmunol. 2009, 212, 102–105. [Google Scholar] [CrossRef]
- Wallace, D.J.; Sayre, N.L.; Patterson, T.T.; Nicholson, S.E.; Hilton, D.; Grandhi, R. Spinal cord injury and the human microbiome: Beyond the brain-gut axis. Neurosurg. Focus 2019, 46, E11. [Google Scholar] [CrossRef]
- Mittal, R.; Debs, L.H.; Patel, A.P.; Nguyen, D.; Patel, K.; O’Connor, G.; Grati, M.; Mittal, J.; Yan, D.; Eshraghi, A.A.; et al. Neurotransmitters: The Critical Modulators Regulating Gut–Brain Axis. J. Cell. Physiol. 2017, 232, 2359–2372. [Google Scholar] [CrossRef] [Green Version]
- Mazzoli, R.; Pessione, E. The neuro-endocrinological role of microbial glutamate and GABA signaling. Front. Microbiol. 2016, 7, 1934. [Google Scholar] [CrossRef] [Green Version]
- Gao, K.; Mu, C.L.; Farzi, A.; Zhu, W.Y. Tryptophan Metabolism: A Link between the Gut Microbiota and Brain. Adv. Nutr. 2020, 11, 709–723. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.M.; Hong, T.; Van Pijkeren, J.P.; Hemarajata, P.; Trinh, D.V.; Hu, W.; Britton, R.A.; Kalkum, M.; Versalovic, J. Histamine derived from probiotic Lactobacillus reuteri suppresses tnf via modulation of pka and erk signaling. PLoS ONE 2012, 7, e31951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, C.; Major, A.; Rendon, D.; Lugo, M.; Jackson, V.; Shi, Z.; Mori-Akiyama, Y.; Versalovic, J. Histamine H2 receptor-mediated suppression of intestinal inflammation by probiotic Lactobacillus reuteri. mBio 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Rios-Covian, D.; Salazar, N.; Gueimonde, M.; de los Reyes-Gavilan, C.G. Shaping the metabolism of intestinal Bacteroides population through diet to improve human health. Front. Microbiol. 2017, 8, 376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markowiak-Kopeć, P.; Śliżewska, K. The effect of probiotics on the production of short-chain fatty acids by human intestinal microbiome. Nutrients 2020, 12, 1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frost, G.; Sleeth, M.L.; Sahuri-Arisoylu, M.; Lizarbe, B.; Cerdan, S.; Brody, L.; Anastasovska, J.; Ghourab, S.; Hankir, M.; Zhang, S.; et al. The short-chain fatty acid acetate reduces appetite via a central homeostatic mechanism. Nat. Commun. 2014, 5, 3611. [Google Scholar] [CrossRef] [Green Version]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The role of short-chain fatty acids from gut microbiota in gut-brain communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef] [Green Version]
- Hassan, A.M.; Mancano, G.; Kashofer, K.; Liebisch, G.; Farzi, A.; Zenz, G.; Claus, S.P.; Holzer, P. Anhedonia induced by high-fat diet in mice depends on gut microbiota and leptin. Nutr. Neurosci. 2020, 25, 299–312. [Google Scholar] [CrossRef] [Green Version]
- Nishiwaki, H.; Ito, M.; Ms, T.I.; Hamaguchi, T.; Maeda, T.; Kashihara, K.; Tsuboi, Y.; Ueyama, J.; Shimamura, T.; Mori, H.; et al. Meta-analysis of gut dysbiosis in Parkinson’s disease. Mov. Disord. 2020, 35, 1626–1635. [Google Scholar] [CrossRef]
- Hao, C.; Gao, Z.; Liu, X.; Rong, Z.; Jia, J.; Kang, K.; Guo, W.; Li, J. Intravenous administration of sodium propionate induces antidepressant or prodepressant effect in a dose dependent manner. Sci. Rep. 2020, 10, 19917. [Google Scholar] [CrossRef]
- Nitert, M.D.; Gomez-Arango, L.F.; Barrett, H.L.; McIntyre, H.D.; Anderson, G.J.; Frazer, D.M.; Callaway, L.K. Iron supplementation has minor effects on gut microbiota composition in overweight and obese women in early pregnancy. Br. J. Nutr. 2018, 120, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Simkin, D.R. Microbiome and mental health, specifically as it relates to adolescents. Curr. Psychiatry Rep. 2019, 21, 93. [Google Scholar] [CrossRef] [PubMed]
- Oriach, C.S.; Robertson, R.C.; Stanton, C.; Cryan, J.F.; Dinan, T.G. Food for thought: The role of nutrition in the microbiota-gut–brain axis. Clin. Nutr. Exp. 2016, 6, 25–38. [Google Scholar] [CrossRef] [Green Version]
- Seura, T.; Yoshino, Y.; Fukuwatari, T. The relationship between habitual dietary intake and gut microbiota in young Japanese women. J. Nutr. Sci. Vitaminol. 2017, 63. [Google Scholar] [CrossRef] [Green Version]
- Zackular, J.; Moore, J.; Jordan, A.T.; Juttukonda, L.J.; Noto, M.J.; Nicholson, M.; Crews, J.D.; Semler, M.W.; Zhang, Y.; Ware, L.B.; et al. Dietary zinc alters the microbiota and decreases resistance to Clostridium difficile infection. Nat. Med. 2016, 22, 1330–1334. [Google Scholar] [CrossRef] [Green Version]
- Fang, S.; Zhuo, Z.; Yu, X.; Wang, H.; Feng, J. Oral administration of liquid iron preparation containing excess iron induces intestine and liver injury, impairs intestinal barrier function and alters the gut microbiota in rats. J. Trace Elem. Med. Biol. 2018, 47, 12–20. [Google Scholar] [CrossRef]
- Chaplin, A.; Parra, P.; Laraichi, S.; Serra, F.; Palou, A. Calcium supplementation modulates gut microbiota in a prebiotic manner in dietary obese mice. Mol. Nutr. Food Res. 2016, 60, 468–480. [Google Scholar] [CrossRef]
- Trautvetter, U.; Camarinha-Silva, A.; Jahreis, G.; Lorkowski, S.; Glei, M. High phosphorus intake and gut-related parameters—Results of a randomized placebo-controlled human intervention study. Nutr. J. 2018, 17, 23. [Google Scholar] [CrossRef]
- Shao, Y.; Lei, Z.; Yuan, J.; Yang, Y.; Guo, Y.; Zhang, B. Effect of zinc on growth performance, gut morphometry, and cecal microbial community in broilers challenged with Salmonella enterica serovar typhimurium. J. Microbiol. 2014, 52, 1002–1011. [Google Scholar] [CrossRef]
- Shen, H.; Han, J.; Li, Y.; Lu, C.; Zhou, J.; Li, Y.; Su, X. Different host-specific responses in thyroid function and gut microbiota modulation between diet-induced obese and normal mice given the same dose of iodine. Appl. Microbiol. Biotechnol. 2019, 103, 3537–3547. [Google Scholar] [CrossRef]
- Zhong, S.R.; Kuang, Q.; Zhang, F.; Chen, B.; Zhong, Z.G. Functional roles of the microbiota-gut-brain axis in Alzheimer’s disease: Implications of gut microbiota-targeted therapy. Transl Neurosci. 2021, 12, 581–600. [Google Scholar] [CrossRef] [PubMed]
- Kapourchali, F.R.; Cresci, G.A.M. Early-life gut microbiome—The importance of maternal and infant factors in its establishment. Nutr. Clin. Pract. 2020, 35, 386–405. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Bello, M.G.; Costello, E.K.; Contreras, M.; Magris, M.; Hidalgo, G.; Fierer, N.; Knight, R. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc. Natl. Acad. Sci. USA 2010, 107, 11971–11975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marwarha, G.; Claycombe-Larson, K.; Schommer, J.; Ghribi, O. Maternal low-protein diet decreases brain-derived neurotrophic factor expression in the brains of the neonatal rat offspring. J. Nutr. Biochem. 2017, 45, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Braniste, V.; Al-Asmakh, M.; Kowal, C.; Anuar, F.; Abbaspour, A.; Tóth, M.; Korecka, A.; Bakocevic, N.; Ng, L.G.; Kundu, P.; et al. The gut microbiota influences blood-brain barrier permeability in mice. Sci. Transl. Med. 2014, 6, 263ra158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurokawa, S.; Kishimoto, T.; Mizuno, S.; Masaoka, T.; Naganuma, M.; Liang, K.-C.; Kitazawa, M.; Nakashima, M.; Shindo, C.; Suda, W.; et al. The effect of fecal microbiota transplantation on psychiatric symptoms among patients with irritable bowel syndrome, functional diarrhea and functional constipation: An open-label observational study. J. Affect. Disord. 2018, 235, 506–512. [Google Scholar] [CrossRef] [PubMed]
- Riezzo, G.; Orlando, A.; D’Attoma, B.; Linsalata, M.; Martulli, M.; Russo, F. Randomised double blind placebo controlled trial on Lactobacillus reuteri DSM 17938: Improvement in symptoms and bowel habit in functional constipation. Benef. Microbes 2018, 9, 51–60. [Google Scholar] [CrossRef]
- Rodriguez, D.A.; Popov, J.; Ratcliffe, E.M.; Monjaraz, E.M.T. Functional constipation and the gut microbiome in children: Preclinical and clinical evidence. Front. Pediatr. 2021, 8, 595531. Available online: https://www.frontiersin.org/articles/10.3389/fped.2020.595531/full (accessed on 20 September 2022). [CrossRef]
- Zheng, Y.-F.; Liang, S.-P.; Zhong, Z.-S.; Zhang, W.; Wu, Y.-Y.; Liu, J.-B.; Huang, S.-P. Duodenal microbiota makes an important impact in functional dyspepsia. Microb. Pathog. 2022, 162, 105297. [Google Scholar] [CrossRef]
- Faramarzi, M.; Azadfallah, P.; Book, H.E.; Tabatabai, K.R.; Taherim, H.; Kashifard, M. The effect of psychotherapy in improving physical and psychiatric symptoms in patients with functional dyspepsia. Iran. J. Psychiatry 2015, 10, 43–49. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4434428/ (accessed on 8 December 2021).
- Igarashi, M.; Nakae, H.; Matsuoka, T.; Takahashi, S.; Hisada, T.; Tomita, J.; Koga, Y. Alteration in the gastric microbiota and its restoration by probiotics in patients with functional dyspepsia. BMJ Open Gastroenterol. 2017, 4, e000144. [Google Scholar] [CrossRef] [PubMed]
- Drago, L.; Meroni, G.; Pistone, D.; Pasquale, L.; Milazzo, G.; Monica, F.; Aragona, S.; Ficano, L.; Vassallo, R.; Gastrobiota Group. Evaluation of main functional dyspepsia symptoms after probiotic administration in patients receiving conventional pharmacological therapies. J. Int. Med. Res. 2021, 49, 1–13. [Google Scholar] [CrossRef]
- Ponziani, F.R.; Scaldaferri, F.; Petito, V.; Paroni Sterbini, F.; Pecere, S.; Lopetuso, L.R.; Palladini, A.; Gerardi, V.; Masucci, L.; Pompili, M.; et al. The role of antibiotics in gut microbiota modulation: The eubiotic effects of rifaximin. Dig. Dis. 2016, 34, 269–278. [Google Scholar] [CrossRef]
- Ahyani, T.; Supriatmo, S.; Deliana, M.; Yudiyanto, A.R.; Sinuhaji, A.B. Lactobacillus probiotics for treating functional dyspepsia in children. Paediatr. Indones. 2016, 56, 37–42. [Google Scholar] [CrossRef]
- Hashash, J.G.; Azar, C.; Elhajj, I.I.; El Zahabi, L.; Chaar, H.F.; Sharara, A.I.; Abdul-Baki, H. Clinical trial: A randomized controlled cross-over study of flupenthixol + melitracen in functional dyspepsia. Aliment. Pharmacol. Ther. 2008, 27, 1148–1155. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Li, X.; Huang, Y.; Xu, X.; Liu, Y.; Wang, J.; Nie, G.; Zhou, D. Comparative efficacy and acceptability of psychotropic drugs for functional dyspepsia in adults: A systematic review and network meta-analysis. Medicine 2021, 100, e26046. [Google Scholar] [CrossRef] [PubMed]
- Shin, A.; Preidis, G.A.; Shulman, R.; Kashyap, P.C. The gut microbiome in adult and pediatric functional gastrointestinal disorders. Clin. Gastroenterol. Hepatol. 2019, 17, 256. [Google Scholar] [CrossRef]
- Savino, F.; Castagno, E.; Bretto, R.; Brondello, C.; Palumeri, E.; Oggero, R. A prospective 10-year study on children who had severe infantile colic. Acta Paediatr. Suppl. 2005, 94, 129–132. [Google Scholar] [CrossRef]
- Reis, G.; Monteiro, T. Intestinal microflora in breastfed colicky and non-colicky infants. Nascer Crescer 2004, 13, 141–144. [Google Scholar] [CrossRef]
- Niesler, B.; Kuerten, S.; Demir, I.E.; Schäfer, K.H. Disorders of the enteric nervous system—A holistic view. Nat. Rev. Gastroenterol. Hepatol. 2021, 18. [Google Scholar] [CrossRef]
- Hsiao, E.Y.; McBride, S.W.; Hsien, S.; Sharon, G.; Hyde, E.R.; McCue, T.; Codelli, J.A.; Chow, J.; Reisman, S.E.; Petrosino, J.F.; et al. Microbiota modulate behavioral and physiological abnormalities associated with neurodevelopmental disorders. Cell 2013, 155, 1451–1463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koenig, J.E.; Spor, A.; Scalfone, N.; Fricker, A.D.; Stombaugh, J.; Knight, R.; Angenent, L.T.; Ley, R.E. Succession of microbial consortia in the developing infant gut microbiome. Proc. Natl. Acad. Sci. USA 2011, 108, 4578–4585. [Google Scholar] [CrossRef]
- Pärtty, A.; Kalliomäki, M.; Wacklin, P.; Salminen, S.; Isolauri, E. A possible link between early probiotic intervention and the risk of neuropsychiatric disorders later in childhood: A randomized trial. Pediatr. Res. 2015, 77, 823–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minter, M.R.; Hinterleitner, R.; Meisel, M.; Zhang, C.; Leone, V.; Zhang, X.; Oyler-Castrillo, P.; Zhang, X.; Musch, M.W.; Shen, X.; et al. Antibiotic-induced perturbations in microbial diversity during post-natal development alters amyloid pathology in an aged APP SWE/PS1 ΔE9 murine model of Alzheimer’s disease. Sci. Rep. 2017, 7, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laue, H.E.; Karagas, M.R.; Coker, M.O.; Bellinger, D.C.; Baker, E.R.; Korrick, S.A.; Madan, J.C. Sex-specific relationships of the infant microbiome and early-childhood behavioral outcomes. Pediatr. Res. 2021, 92, 580–591. [Google Scholar] [CrossRef] [PubMed]
- Agans, R.; Rigsbee, L.; Kenche, H.; Michail, S.; Khamis, H.J.; Paliy, O. Distal gut microbiota of adolescent children is different from that of adults. FEMS Microbiol. Ecol. 2011, 77, 404–412. [Google Scholar] [CrossRef] [Green Version]
- Lasheras, I.; Seral, P.; Latorre, E.; Barroso, E.; Gracia-García, P.; Santabárbara, J. Microbiota and gut-brain axis dysfunction in autism spectrum disorder: Evidence for functional gastrointestinal disorders. Asian J. Psychiatr. 2020, 47, 101874. [Google Scholar] [CrossRef]
- Mitchell, L.K.; Davies, P.S.W. Pre- and probiotics in the management of children with autism and gut issues: A review of the current evidence. Eur. J. Clin. Nutr. 2021, 76, 913–921. [Google Scholar] [CrossRef]
- Roohafza, H.; Molaee, A.; Afshar, H.; Mazaheri, M.; Feizi, A.; Hassanzadeh-Keshteli, A.; Adibi, P. The association of psychological comorbidity with the number of functional gastrointestinal disorders. Int. J. Body Mind Cult. 2016, 3, 22–29. [Google Scholar] [CrossRef]
- Reigstad, C.S.; Kashyap, P.C. Beyond phylotyping: Understanding the impact of gut microbiota on host biology. Neurogastroenterol. Motil. 2013, 25, 358–372. [Google Scholar] [CrossRef] [Green Version]
- Touw, K.; Ringus, D.L.; Hubert, N.; Wang, Y.; Leone, V.A.; Nadimpalli, A.; Theriault, B.R.; Huang, Y.E.; Tune, J.D.; Herring, P.B.; et al. Mutual reinforcement of pathophysiological host-microbe interactions in intestinal stasis models. Physiol. Rep. 2017, 5, e13182. [Google Scholar] [CrossRef] [PubMed]
- Crouzet, L.; Gaultier, E.; Del’Homme, C.; Cartier, C.; Delmas, E.; Dapoigny, M.; Fioramonti, J.; Bernalier-Donadille, A. The hypersensitivity to colonic distension of IBS patients can be transferred to rats through their fecal microbiota. Neurogastroenterol. Motil. 2013, 25, e272–e282. [Google Scholar] [CrossRef] [PubMed]
- Kunze, W.A.; Mao, Y.; Wang, B.; Huizinga, J.D.; Ma, X.; Forsythe, P.; Bienenstock, J. Lactobacillus reuteri enhances excitability of colonic AH neurons by inhibiting calcium-dependent potassium channel opening. J. Cell. Mol. Med. 2009, 13, 2261–2270. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, S.C.; Barbara, G.; Buurman, W.; Ockhuizen, T.; Schulzke, J.-D.; Serino, M.; Tilg, H.; Watson, A.; Wells, J.M. Intestinal permeability—A new target for disease prevention and therapy. BMC Gastroenterol. 2014, 14, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desai, M.S.; Seekatz, A.M.; Koropatkin, N.M.; Kamada, N.; Hickey, C.A.; Wolter, M.; Pudlo, N.A.; Kitamoto, S.; Terrapon, N.; Muller, A.; et al. A dietary fiber-deprived gut microbiota degrades the colonic mucus barrier and enhances pathogen susceptibility. Cell 2016, 167, 1339–1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasapolli, R.; Schulz, C.; Schweden, M.; Casèn, C.; Kirubakaran, G.T.; Kirste, K.H.; Macke, L.; Link, A.; Schütte, K.; Malfertheiner, P. Gut microbiota profiles and the role of anti-CdtB and anti-vinculin antibodies in patients with functional gastrointestinal disorders (FGID). Eur. J. Clin. Investig. 2021, 51, e13666. [Google Scholar] [CrossRef] [PubMed]
- Qamra, A.; Soni, N.K.; Trivedi, H.H.; Kumar, S.; Prakash, A.; Roy, S.; Mukherjee, S. A review of digestive enzyme and probiotic supplementation for functional gastrointestinal disorders. Indian Pract. 2020, 73, 35–39. Available online: http://articles.theindianpractitioner.com/index.php/tip/article/download/944/897 (accessed on 20 September 2022).
- Bercik, P.; Park, A.J.; Sinclair, D.; Khoshdel, A.; Lu, J.; Huang, X.; Deng, Y.; Blennerhassett, P.A.; Fahnestock, M.; Moine, D.; et al. The anxiolytic effect of Bifidobacterium longum NCC3001 involves vagal pathways for gut–brain communication. Neurogastroenterol. Motil. 2011, 23, 1132–1139. [Google Scholar] [CrossRef] [Green Version]
- Rome IV Criteria—Rome Foundation. Available online: https://theromefoundation.org/rome-iv/rome-iv-criteria/ (accessed on 8 December 2021).
- Bhattarai, Y.; Pedrogo, D.A.M.; Kashyap, P.C. Irritable bowel syndrome: A gut microbiota-related disorder? Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 312, G52–G62. [Google Scholar] [CrossRef]
- De Palma, G.; Lynch, M.D.J.; Lu, J.; Dang, V.T.; Deng, Y.; Jury, J.; Umeh, G.; Miranda, P.M.; Pastor, M.P.; Sidani, S.; et al. Transplantation of fecal microbiota from patients with irritable bowel syndrome alters gut function and behavior in recipient mice. Sci. Transl. Med. 2017, 9. [Google Scholar] [CrossRef]
- Agrawal, A.; Houghton, L.A.; Morris, J.; Reilly, B.; Guyonnet, D.; Goupil Feuillerat, N.; Schlumberger, A.; Jakob, S.; Whorwell, P.J. Clinical trial: The effects of a fermented milk product containing Bifidobacterium lactis DN-173 010 on abdominal distension and gastrointestinal transit in irritable bowel syndrome with constipation. Aliment. Pharmacol. Ther. 2009, 29, 104–114. [Google Scholar] [CrossRef]
- Chen, C.H.; Lin, C.L.; Kao, C.H. Irritable bowel syndrome is associated with an increased risk of dementia: A nationwide population-based study. PLoS ONE 2016, 11, e0144589. [Google Scholar] [CrossRef]
- Liu, P.; Li, G.; Zhang, A.; Yang, C.; Liu, Z.; Sun, N.; Kerang, Z. Brain Structural and Functional Alterations in MDD Patient with Gastrointestinal Symptoms: A Resting-State MRI Study; Elsevier: Amsterdam, The Netherlands, 2020; Available online: https://www.sciencedirect.com/science/article/pii/S0165032719325339 (accessed on 8 December 2021).
- Shankar, V.; Reo, N.V.; Paliy, O. Simultaneous fecal microbial and metabolite profiling enables accurate classification of pediatric irritable bowel syndrome. Microbiome 2015, 3, 73. [Google Scholar] [CrossRef]
- Horvath, A.; Dziechciarz, P.; Szajewska, H. Meta-analysis: Lactobacillus rhamnosus GG for abdominal pain-related functional gastrointestinal disorders in childhood. Aliment. Pharmacol. Ther. 2011, 33, 1302–1310. [Google Scholar] [CrossRef]
- Abomoelak, B.; Pemberton, V.; Deb, C.; Campion, S.; Vinson, M.; Mauck, J.; Manipadam, J.; Sudakaran, S.; Patel, S.; Saps, M.; et al. The gut microbiome alterations in pediatric patients with functional abdominal pain disorders. Microorganisms 2021, 9, 2354. [Google Scholar] [CrossRef]
- Tan, V.P.Y.; Liu, K.S.H.; Lam, F.Y.F.; Hung, I.F.N.; Yuen, M.F.; Leung, W.K. Randomised clinical trial: Rifaximin versus placebo for the treatment of functional dyspepsia. Aliment. Pharmacol. Ther. 2017, 45, 767–776. [Google Scholar] [CrossRef] [Green Version]
- Schmulson, M.; Chang, L. Review article: The treatment of functional abdominal bloating and distension. Aliment. Pharmacol. Ther. 2011, 33, 1071–1086. [Google Scholar] [CrossRef]
- Guandalini, S.; Magazzù, G.; Chiaro, A.; La Balestra, V.; Di Nardo, G.; Gopalan, S.; Sibal, A.; Romano, C.; Canani, R.B.; Lionetti, P.; et al. VSL#3 improves symptoms in children with irritable bowel syndrome: A multicenter, randomized, placebo-controlled, double-blind, crossover study. J. Pediatr. Gastroenterol. Nutr. 2010, 51, 24–30. [Google Scholar] [CrossRef]
- Hosseinzadeh, S.T.; Poorsaadati, S.; Radkani, B.; Forootan, M. Psychological disorders in patients with chronic constipation. Gastroenterol. Hepatol. Bed Bench 2011, 4, 159. [Google Scholar] [CrossRef]
- Savino, F.; Bailo, E.; Oggero, R.; Tullio, V.; Roana, J.; Carlone, N.; Cuffini, A.M.; Silvestro, L. Bacterial counts of intestinal Lactobacillus species in infants with colic. Pediatr. Allergy Immunol. 2005, 16, 72–75. [Google Scholar] [CrossRef]
- Skonieczna-żydecka, K.; Marlicz, W.; Misera, A.; Koulaouzidis, A.; Łoniewski, I. Microbiome—The missing link in the gut-brain axis: Focus on its role in gastrointestinal and mental health. J. Clin. Med. 2018, 7, 521. [Google Scholar] [CrossRef] [Green Version]
- Shirazinia, R.; Golabchifar, A.A.; Fazeli, M.R. Efficacy of probiotics for managing infantile colic due to their anti-inflammatory properties: A meta-analysis and systematic review. Clin. Exp. Pediatr. 2021, 64, 642–651. [Google Scholar] [CrossRef]
- Liang, S.; Wu, X.; Jin, F. Gut-brain psychology: Rethinking psychology from the microbiota–gut–brain axis. Front. Integr. Neurosci. 2018, 12, 33. [Google Scholar] [CrossRef] [Green Version]
- Ozkul, C.; Ruiz, V.E.; Battaglia, T.; Xu, J.; Roubaud-Baudron, C.; Cadwell, K.; Perez, G.P.; Blaser, M.J. A single early-in-life antibiotic course increases susceptibility to DSS-induced colitis. Genome Med. 2020, 12, 65. [Google Scholar] [CrossRef]
- Karakan, T.; Ozkul, C.; Akkol, E.K.; Bilici, S.; Sobarzo-Sánchez, E.; Capasso, R. Gut-brain-microbiota axis: Antibiotics and functional gastrointestinal disorders. Nutrients 2021, 13, 389. [Google Scholar] [CrossRef]
- Kennedy, E.A.; King, K.Y.; Baldridge, M.T. Mouse microbiota models: Comparing germ-free mice and antibiotics treatment as tools for modifying gut bacteria. Front. Physiol. 2018, 9, 1534. [Google Scholar] [CrossRef] [Green Version]
- Jones, M.P.; Shah, A.; Walker, M.M.; Koloski, N.A.; Holtmann, G.; Talley, N.J. Antibiotic use but not gastrointestinal infection frequently precedes first diagnosis of functional gastrointestinal disorders. UEG J. 2021, 9, 1074–1080. [Google Scholar] [CrossRef]
- Kamphorst, K.; van Daele, E.; Vlieger, A.M.; Daams, J.G.; Knol, J.; van Elburg, R.M. Early life antibiotics and childhood gastrointestinal disorders: A systematic review. BMJ Paediatr. Open 2021, 5, e001028. [Google Scholar] [CrossRef]
- Mechetina, T.A.; Il’chenko, A.A.; Lychkova, A.E. Rifaximin application in the overgrowth bacterial syndrome in the small intestine in patients after cholecystectomy. Eksp. Klin. Gastroenterol. 2011, 3, 93–100. [Google Scholar]
- Cangemi, D.J.; Lacy, B.E.; Wise, J. Diagnosing small intestinal bacterial overgrowth: A comparison of lactulose breath tests to small bowel aspirates. Dig. Dis. Sci. 2021, 66, 2042–2050. [Google Scholar] [CrossRef]
- Jones, J.; Lembo, A.; Heidelbaugh, J.; Kuritzky, L.; Lacy, B. Management of irritable bowel syndrome with diarrhea: Focus on eluxadoline. Curr. Med. Res. Opin. 2021, 37, 567–578. [Google Scholar] [CrossRef] [PubMed]
- Wegh, C.A.M.; Schoterman, M.H.C.; Vaughan, E.E.; Belzer, C.; Benninga, M.A. The effect of fiber and prebiotics on children’s gastrointestinal disorders and microbiome. Expert Rev. Gastroenterol. Hepatol. 2017, 11, 1031–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, B.; Whelan, K. Prebiotic inulin-type fructans and galacto-oligosaccharides: Definition, specificity, function, and application in gastrointestinal disorders. J. Gastroenterol. Hepatol. 2017, 32, 64–68. [Google Scholar] [CrossRef] [Green Version]
- Saulnier, D.M.; Ringel, Y.; Heyman, M.B.; Foster, J.A.; Bercik, P.; Shulman, R.J.; Versalovic, J.; Verdu, E.F.; Dinan, T.G.; Hecht, G.; et al. The intestinal microbiome, probiotics and prebiotics in neurogastroenterology. Gut Microbes 2013, 4, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Chao, L.; Liu, C.; Sutthawongwadee, S.; Li, Y.; Lv, W.; Chen, W.; Yu, L.; Zhou, J.; Guo, A.; Li, Z.; et al. Effects of probiotics on depressive or anxiety variables in healthy participants under stress conditions or with a depressive or anxiety diagnosis: A meta-analysis of randomized controlled trials. Front. Neurol. 2020, 11, 421. [Google Scholar] [CrossRef]
- Messaoudi, M.; Lalonde, R.; Violle, N.; Javelot, H.; Desor, D.; Nejdi, A.; Bisson, J.F.; Rougeot, C.; Pichelin, M.; Cazaubiel, M.; et al. Assessment of psychotropic-like properties of a probiotic formulation (Lactobacillus helveticus R0052 and Bifidobacterium longum R0175) in rats and human subjects. Br. J. Nutr. 2011, 105, 755–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Compare, D.; Rocco, A.; Coccoli, P.; Angrisani, D.; Sgamato, C.; Iovine, B.; Salvatore, U.; Nardone, G. Lactobacillus casei DG and its postbiotic reduce the inflammatory mucosal response: An ex-vivo organ culture model of post-infectious irritable bowel syndrome. BMC Gastroenterol. 2017, 17, 53. [Google Scholar] [CrossRef] [Green Version]
- Paiva IH, R.; Duarte-Silva, E.; Peixoto, C.A. The role of prebiotics in cognition, anxiety, and depression. Eur. Neuropsychopharmacol. 2020, 34, 1–18. [Google Scholar] [CrossRef]
- Rafsky, H.A.; Rafsky, J.C. Clinical and bacteriological studies of a new Lactobacillus acidophilus concentrate in functional gastrointestinal disturbances. Am. J. Gastroenterol. 1955, 24, 87–92. [Google Scholar]
- Romijn, A.R.; Rucklidge, J.J. Systematic review of evidence to support the theory of psychobiotics. Nutr. Rev. 2015, 73, 675–693. [Google Scholar] [CrossRef] [Green Version]
- Lai, C.-T.; Chen, C.-Y.; She, S.-C.; Chen, W.-J.; Kuo, T.B.; Lin, H.-C.; Yang, C.C. Production of Lactobacillus brevis ProGA28 attenuates stress-related sleep disturbance and modulates the autonomic nervous system and the motor response in anxiety/depression behavioral tests in Wistar–Kyoto rats. Life Sci. 2022, 288, 120165. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Ding, S.; Ma, Y.; Fang, J.; Jiang, H.; Li, Y.; Liu, G. Lactobacillus plantarum and Lactobacillus brevis alleviate intestinal inflammation and microbial disorder induced by ETEC in a murine model. Oxidative Med. Cell. Longev. 2021, 2021, 6867962. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Qing-Jie, F.; Yin-Hui, L.; Ming, L.; Jie-Li, Y. Fermentation production of ganoderma lucidum by Bacillus subtilis ameliorated ceftriaxone-induced intestinal dysbiosis and improved intestinal mucosal barrier function in mice. Digit. Chin. Med. 2020, 3, 199–212. [Google Scholar] [CrossRef]
- Jiang, S.; Hu, J.-Y.; Cheng, H.-W. The impact of probiotic Bacillus subtilis on injurious behavior in laying hens. Animals 2022, 12, 870. [Google Scholar] [CrossRef]
- Pinto-Sanchez, M.I.; Hall, G.B.; Ghajar, K.; Nardelli, A.; Bolino, C.; Lau, J.T.; Martin, F.-P.; Cominetti, O.; Welsh, C.; Rieder, A.; et al. Probiotic Bifidobacterium longum NCC3001 reduces depression scores and alters brain activity: A pilot study in patients with irritable bowel syndrome. Gastroenterology 2017, 153, 448–459. [Google Scholar] [CrossRef]
- Vitellio, P.; Celano, G.; Bonfrate, L.; Gobbetti, M.; Portincasa, P.; De Angelis, M. Effects of Bifidobacterium longum and Lactobacillus rhamnosus on gut microbiota in patients with lactose intolerance and persisting functional gastrointestinal symptoms: A randomised, double-blind, cross-over study. Nutrients 2019, 11, 886. [Google Scholar] [CrossRef] [Green Version]
- Dalmasso, G.; Cottrez, F.; Imbert, V.; Lagadec, P.; Peyron, J.F.; Rampal, P.; Czerucka, D.; Groux, H. Saccharomyces boulardii inhibits inflammatory bowel disease by trapping T cells in mesenteric lymph nodes. Gastroenterology 2006, 131, 1812–1825. [Google Scholar] [CrossRef]
- Aghamohammadi, D.; Ayromlou, H.; Dolatkhah, N.; Jahanjoo, F.; Shakouri, S.K. The effects of probiotic Saccharomyces boulardii on the mental health, quality of life, fatigue, pain, and indices of inflammation and oxidative stress in patients with multiple sclerosis: Study protocol for a double-blind randomized controlled clinical trial. Trials 2019, 20, 379. [Google Scholar]
- Stoyanova, L.G.; Vodolazov, I.V.; Dbar, S.D.; Oleskin, A.V. Probiotic strains of Lactococcus lactis subsp. lactis produce neuroactive substances. J. Hyg. Eng. Des. 2017, 20, 25–31. [Google Scholar]
- Sałański, P.; Kowalczyk, M.; Bardowski, J.K.; Szczepankowska, A.K. Health-promoting nature of Lactococcus lactis IBB109 and Lactococcus lactis IBB417 strains exhibiting proliferation inhibition and stimulation of Interleukin-18 expression in colorectal cancer cells. Front. Microbiol. 2022, 13, 822912. [Google Scholar] [CrossRef]
- Ait-Belgnaoui, A.; Payard, I.; Rolland, C.; Harkat, C.; Braniste, V.; Théodorou, V.; Tompkins, T.A. Bifidobacterium longum and Lactobacillus helveticus synergistically suppress stress-related visceral hypersensitivity through hypothalamic-pituitary-adrenal axis modulation. J. Neurogastroenterol. Motil. 2018, 24, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louis, P.; Duncan, S.; Sheridan, P.; Walker, A.; Flint, H. Microbial lactate utilisation and the stability of the gut microbiome. Gut Microbiome 2022, 3, E3. [Google Scholar] [CrossRef]
- Liang, S.; Wang, T.; Hu, X.; Luo, J.; Li, W.; Wu, X.; Duan, Y.; Jin, F. Administration of Lactobacillus helveticus NS8 improves behavioral, cognitive, and biochemical aberrations caused by chronic restraint stress. Neuroscience 2015, 310, 561–577. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, A.; Lehto, S.M.; Harty, S.; Dinan, T.G.; Cryan, J.F.; Burnet, P.W. Psychobiotics and the manipulation of bacteria–gut–brain signals. Trends Neurosci. 2016, 39, 763–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, V.F.; Elias-Oliveira, J.; Pereira, Í.S.; Pereira, J.A.; Barbosa, S.C.; Machado, M.S.G.; Carlos, D. Akkermansia muciniphila and gut immune system: A good friendship that attenuates inflammatory bowel disease, obesity, and diabetes. Front. Immunol. 2022, 7, 934695. [Google Scholar] [CrossRef]
- Xu, R.; Zhang, Y.; Chen, S.; Zeng, Y.; Fu, X.; Chen, T.; Luo, S.; Zhang, X. The role of the probiotic Akkermansia muciniphila in brain functions: Insights underpinning therapeutic potential. Crit. Rev. Microbiol. 2022, 1–26. [Google Scholar] [CrossRef]
- Pärtty, A.; Rautava, S.; Kalliomäki, M. Probiotics on pediatric functional gastrointestinal disorders. Nutrients 2018, 10, 1836. [Google Scholar] [CrossRef]
- Skonieczna-żydecka, K.; Janda, K.; Kaczmarczyk, M.; Marlicz, W.; Łoniewski, I.; Łoniewska, B. The effect of probiotics on symptoms, gut microbiota and inflammatory markers in infantile colic: A systematic review, meta-analysis and meta-regression of randomized controlled trials. J. Clin. Med. 2020, 9, 999. [Google Scholar] [CrossRef] [Green Version]
- Tait, C.; Sayuk, G.S. The brain-gut-microbiotal axis: A framework for understanding functional GI illness and their therapeutic interventions. Eur. J. Intern. Med. 2021, 84, 1–9. [Google Scholar] [CrossRef]
- Staller, K.; Thurler, A.H.; Reynolds, J.S.; Dimisko, L.R.; McGovern, R.; Skarbinski, K.F.; Kuo, B. Gabapentin improves symptoms of functional dyspepsia in a retrospective, open-label cohort study. J. Clin. Gastroenterol. 2019, 53, 379–384. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Șchiopu, C.G.; Ștefănescu, C.; Boloș, A.; Diaconescu, S.; Gilca-Blanariu, G.-E.; Ștefănescu, G. Functional Gastrointestinal Disorders with Psychiatric Symptoms: Involvement of the Microbiome–Gut–Brain Axis in the Pathophysiology and Case Management. Microorganisms 2022, 10, 2199. https://doi.org/10.3390/microorganisms10112199
Șchiopu CG, Ștefănescu C, Boloș A, Diaconescu S, Gilca-Blanariu G-E, Ștefănescu G. Functional Gastrointestinal Disorders with Psychiatric Symptoms: Involvement of the Microbiome–Gut–Brain Axis in the Pathophysiology and Case Management. Microorganisms. 2022; 10(11):2199. https://doi.org/10.3390/microorganisms10112199
Chicago/Turabian StyleȘchiopu, Cristina Gabriela, Cristinel Ștefănescu, Alexandra Boloș, Smaranda Diaconescu, Georgiana-Emmanuela Gilca-Blanariu, and Gabriela Ștefănescu. 2022. "Functional Gastrointestinal Disorders with Psychiatric Symptoms: Involvement of the Microbiome–Gut–Brain Axis in the Pathophysiology and Case Management" Microorganisms 10, no. 11: 2199. https://doi.org/10.3390/microorganisms10112199