
Screening of Lactiplantibacillus plantarum Strains from Sourdoughs for Biosuppression of Pseudomonas syringae pv. syringae and Botrytis cinerea in Table Grapes

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Material and Methods
2.1. LAB Strains and Plant Pathogens
2.2. Plant Material
2.3. Molecular Analyses
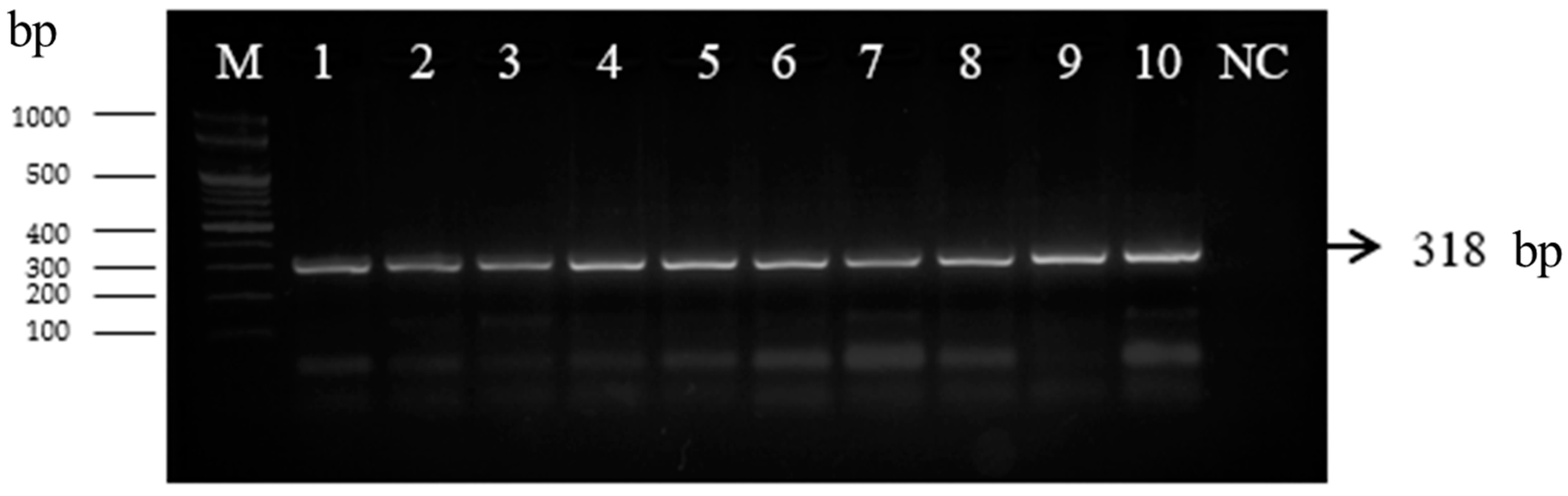
2.3.1. PCR Analysis with recA Gene
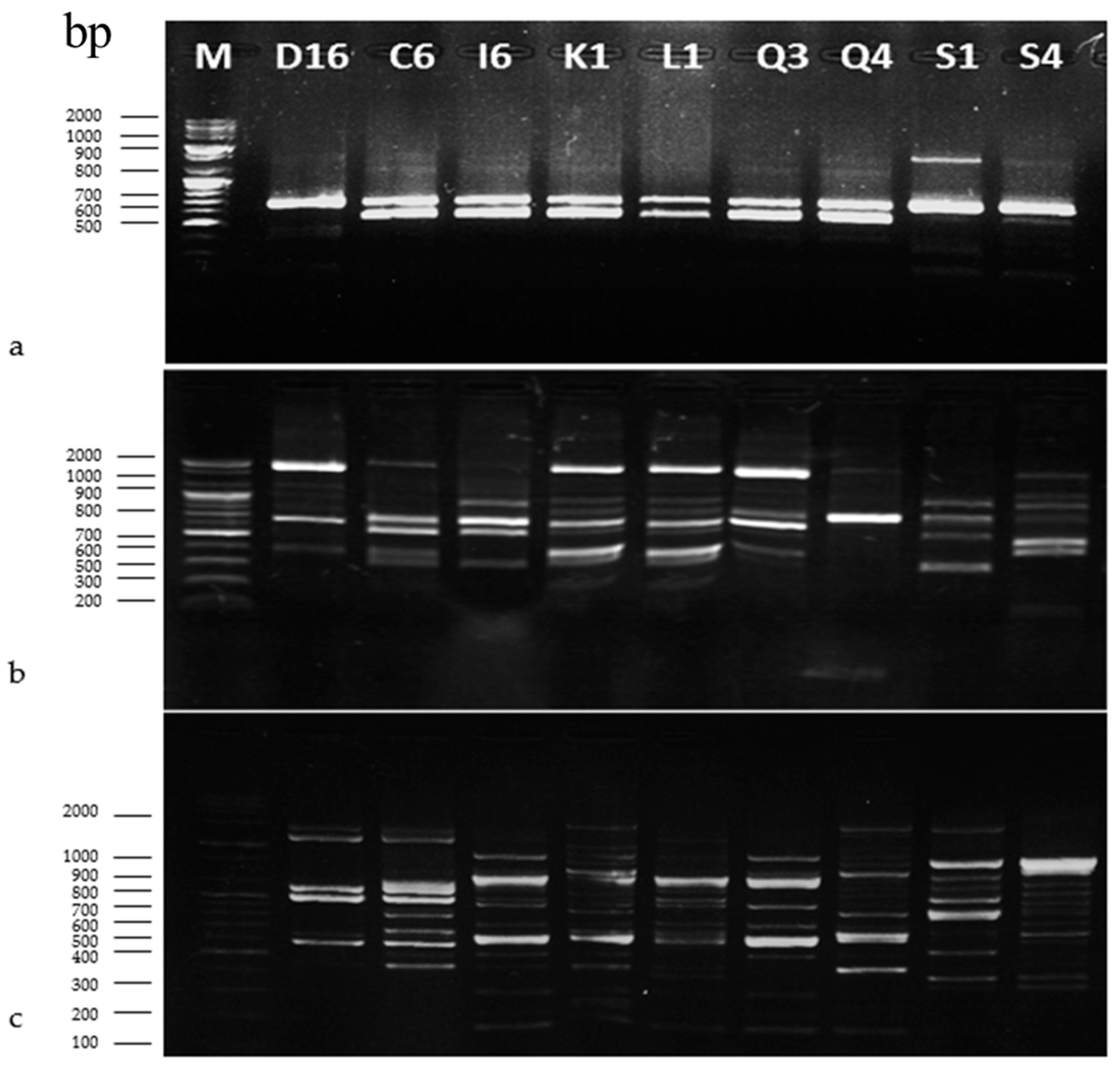
2.3.2. Genotyping of LAB Isolated from Sourdoughs by RAPD-PCR
2.3.3. Screening of Lp. plantarum Strains for the Production of Known Plantaricin Genes
2.4. In Vitro Antimicrobial Activity of LAB Strains
2.4.1. Antibacterial Activity Screening of Cell-Free Supernatants (CFSs) from LAB against Ps. syringae pv. syringae
2.4.2. Antifungal Activity Screening of Cell-Free Supernatants (CFSs) from Tested LAB against B. cinerea
2.5. In Vivo Antimicrobial Activity of Lp. plantarum Strains
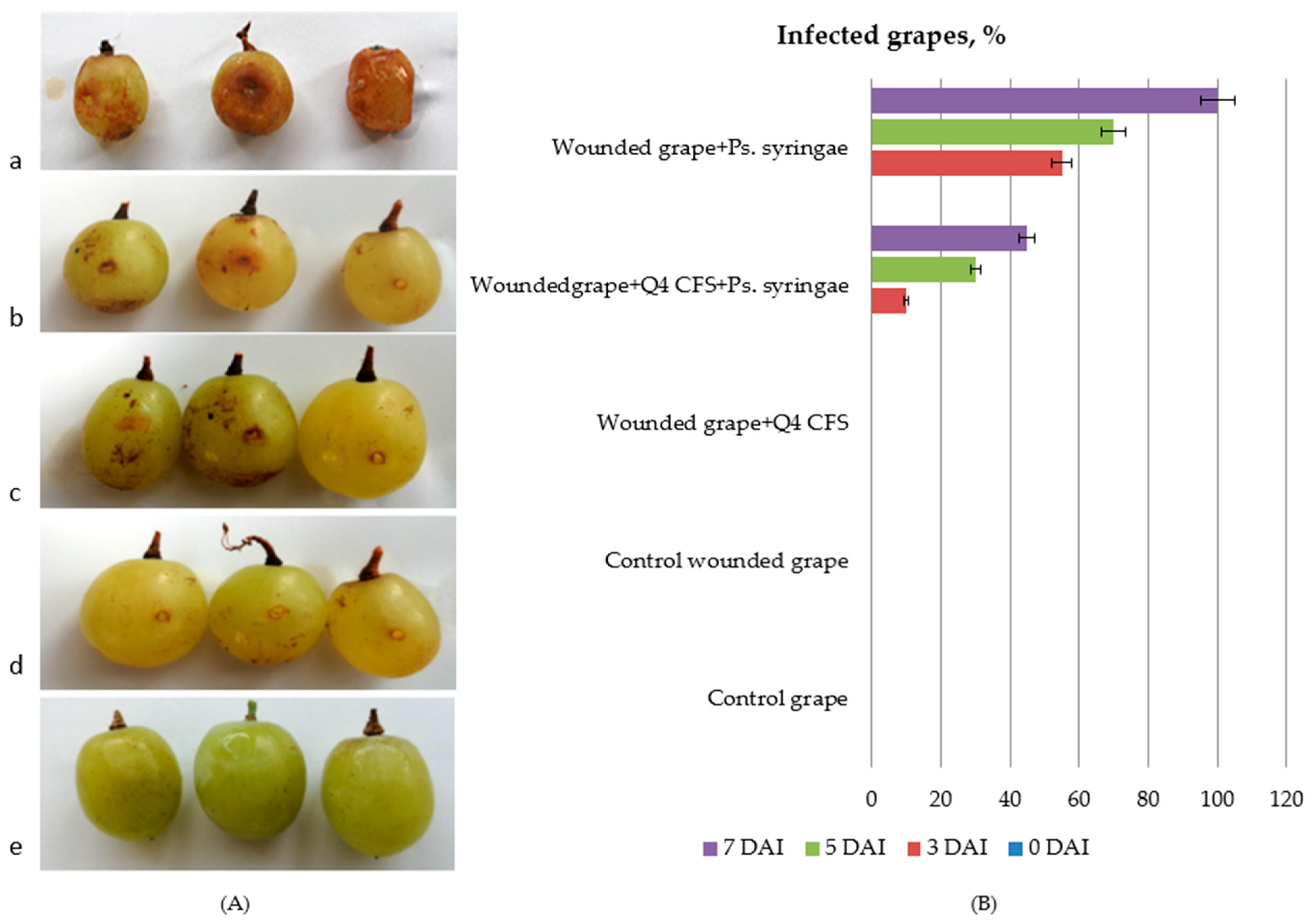
2.5.1. In Vivo Antibacterial Activity of Lp. plantarum Q4 against Ps. syringae pv. syringae in Grapes
2.5.2. In Vivo Antimicrobial Activity of Lp. plantarum Q4 against B. cinerea in Grapes
2.6. Statistical Analysis
3. Results and Discussion
3.1. Plans Material and Plant Pathogenicity Tests
3.2. Molecular Analysis of Lp. plantarum Strains
3.2.1. Microbiological Analysis
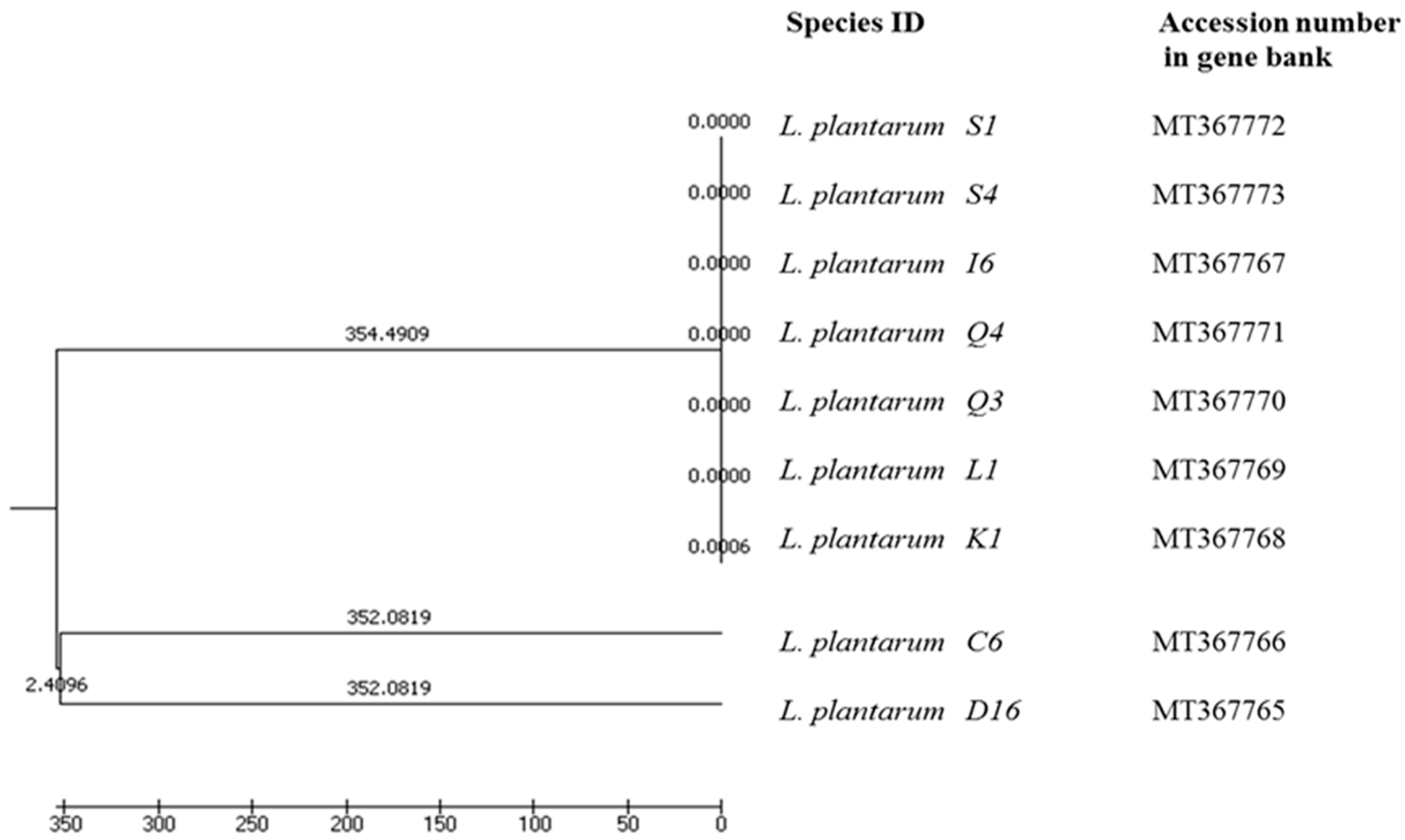
3.2.2. RAPD-PCR Analysis
3.2.3. Detection of Plantaricin Production Genes
3.3. Antimicrobial Activity of LAB Strains
3.3.1. Determination of the Spectrum of Antibacterial Activity of Lp. plantarum Strains against Ps. syringae
3.3.2. Antifungal Activity of Lp. plantarum Strains against B. cinerea
3.4. In Vivo Antimicrobial Activity of Lp. plantarum Q4 against Ps. syringae in Table Grapes
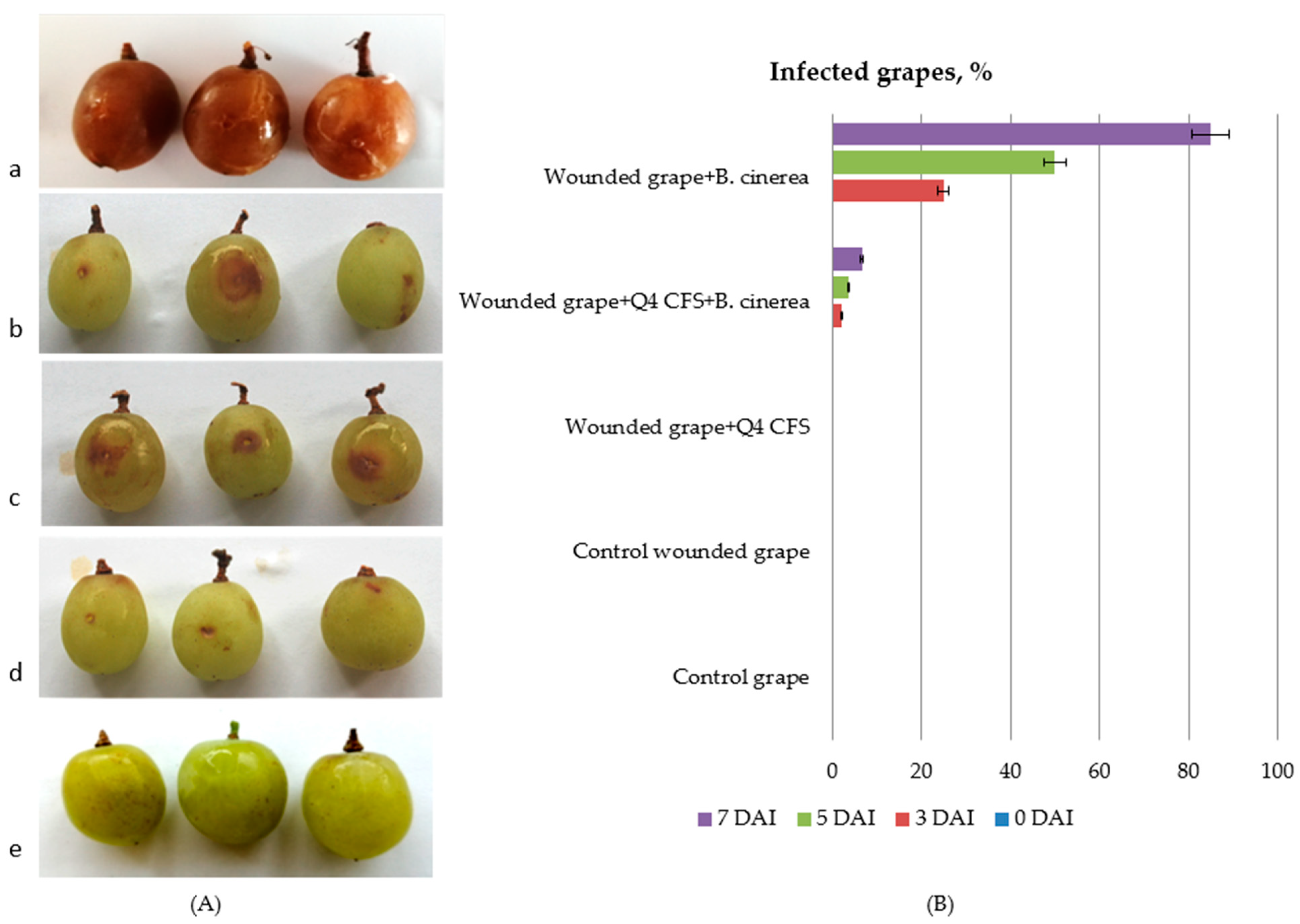
3.5. In Vivo Antimicrobial Activity of Lp. plantarum Q4 against Botrytis cinerea in Grapes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pezzuto, J.M. Grapes and human health: A perspective. J. Agric. Food Chem. 2008, 56, 6777–6784. [Google Scholar] [CrossRef] [PubMed]
- Ilieva, F.; Petrov, K.; Kostadinović-Veličkovska, S.; Gunova, N.; Dimovska, V.; Rocha, J.; Esatbeyoglu, T. Influence of autochthonous and commercial yeast strains on fermentation and quality of wines produced from Vranec and Cabernet Sauvignon grape varieties from Tikveš wine-growing region, Republic of North Macedonia. Appl. Sci. 2021, 11, 6135. [Google Scholar] [CrossRef]
- De Simone, N.; Pace, B.; Grieco, F.; Chimienti, M.; Tyibilika, V.; Santoro, V.; Capozzi, V.; Colelli, G.; Spano, G.; Russo, P. Botrytis cinerea and Table Grapes: A Review of the Main Physical, Chemical, and Bio-Based Control Treatments in Post-Harvest. Foods 2020, 9, 1138. [Google Scholar] [CrossRef] [PubMed]
- Tzanova, M.; Atanasov, V.; Ivanov, M.; Iliev, A.; Atanassova, S.; Peeva, P.; Grozeva, N.; Gerdzhikova, M.; Dinev, T. Antioxidant constituents and antioxidant activity of some red wine and red table grape varieties, cultivated in different regions of Bulgaria. Bulg. J. Agric. Sci. 2019, 25, 3–9. [Google Scholar]
- Holz, G.; Coertze, S.; Williamson, B. The ecology of Botrytis on plant surfaces. In Botrytis: Biology, Pathology and Control; Elad, Y., Williamson, B., Tudzynski, P., Delen, N., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 9–27. [Google Scholar]
- Droby, S.; Lichter, A. Post-harvest Botrytis infection: Etiology, development and management. In Botrytis: Biology, Pathology and Control; Elad, Y., Williamson, B., Tudzynski, P., Delen, N., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 349–367. [Google Scholar]
- Whitelaw-Weckert, M.A.; Whitelaw, E.S.; Rogiers, S.Y.; Quirk, L.; Clark, A.C.; Huang, C.X. Bacterial inflorescence rot of grapevine caused by Pseudomonas syringae pv. syringae. Plant Pathol. 2011, 60, 325–337. [Google Scholar] [CrossRef]
- Gerin, D.; Cariddi, C.; de Miccolis Angelini, R.M.; Rotolo, C.; Dongiovanni, C.; Faretra, F.; Pollastro, S. First report of Pseudomonas grapevine bunch rot caused by Pseudomonas syringae pv. syringae. Plant Dis. 2019, 103, 1954–1960. [Google Scholar] [CrossRef]
- Elmer, P.A.G.; Reglinski, T. Biosuppression of Botrytis cinerea in grapes. Plant Pathol. 2006, 55, 155–177. [Google Scholar] [CrossRef]
- Muri, S.D.; van der Voet, H.; Boon, P.E.; van Klaveren, J.D.; Brüschweiler, B.J. Comparison of human health risks resulting from exposure to fungicides and mycotoxins via food. Food Chem. Toxicol. 2009, 47, 2963–2974. [Google Scholar] [CrossRef] [PubMed]
- Meena, R.S.; Kumar, S.; Datta, R.; Lal, R.; Vijayakumar, V.; Brtnicky, M.; Sharma, M.P.; Yadav, G.S.; Jhariya, M.K.; Jangir, C.K. Impact of agrochemicals on soil microbiota and management: A review. Land 2020, 9, 34. [Google Scholar] [CrossRef] [Green Version]
- Russo, P.; Fragasso, M.; Berbegal, C.; Grieco, F.; Spano, G.; Capozzi, V. Microorganisms able to produce biogenic amines and factors affecting their activity. In Biogenic Amines in Food: Analysis, Occurrence and Toxicity; Saad, B., Tofalo, R., Eds.; RSC Publishing: Cambridge, UK, 2019; pp. 18–40. [Google Scholar]
- Russo, P.; Arena, M.P.; Fiocco, D.; Capozzi, V.; Drider, D.; Spano, G. Lactobacillus plantarum with broad antifungal activity: A promising approach to increase safety and shelf-life of cereal-based products. Int. J. Food Microbiol. 2017, 247, 48–54. [Google Scholar] [CrossRef]
- Linares-Morales, J.R.; Gutiérrez-Méndez, N.; Rivera-Chavira, B.E.; Pérez-Vega, S.B.; Nevárez-Moorillón, G.V. Biocontrol processes in fruits and fresh produce, the use of lactic acid bacteria as a sustainable option. Front. Sustain. Food Syst. 2018, 2, 50. [Google Scholar] [CrossRef]
- Gálvez, A.; Abriouel, H.; López, R.L.; Ben Omar, N. Biological control of pathogens and post-processing spoilage microorganisms in fresh and processed fruit and vegetables. In Protective Cultures, Antimicrobial Metabolites and Bacteriophages for Food and Beverage Biopreservation; Lacroix, C., Ed.; Woodhead Publishing: Sawston, UK, 2011; pp. 403–432. [Google Scholar]
- Hwanhlem, N.; Biscola, V.; El-Ghaish, S.; Jaffrès, E.; Dousset, X.; Haertlé, T.; Chobert, J.M. Bacteriocin-producing lactic acid bacteria isolated from mangrove forests in Southern Thailand as potential bio-control agents: Purification and characterization of bacteriocin produced by Lactococcus lactis subsp. lactis KT2W2L. Probiotics Antimicrob. Proteins 2013, 5, 264–278. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.H.; Liao, M.J.; Wang, H.K.; Zheng, M.Z.; Xu, J.J.; Guo, J.H. Bacillus velezensis, a potential and efficient biocontrol agent in control of pepper gray mold caused by Botrytis cinerea. Biol. Control 2018, 126, 147–157. [Google Scholar] [CrossRef]
- Hamaoka, K.; Aoki, Y.; Suzuki, S. Isolation and characterization of endophyte Bacillus velezensis KOF112 from grapevine shoot xylem as biological control agent for Fungal Diseases. Plants 2021, 10, 1815. [Google Scholar] [CrossRef] [PubMed]
- Petkova, M.; Stefanova, P.; Gotcheva, V.; Kuzmanova, I.; Angelov, A. Microbiological and physicochemical characterization of traditional Bulgarian sourdoughs and screening of lactic acid bacteria for amylolytic activity. J. Chem. Technol. Metall. 2020, 55, 921–934. [Google Scholar]
- Petkova, M.; Stefanova, P.; Gotcheva, V.; Angelov, A. Isolation and characterization of lactic acid bacteria and yeasts from typical Bulgarian sourdoughs. Microorganisms 2021, 9, 1346. [Google Scholar] [CrossRef] [PubMed]
- Chochkov, R.; Savova-Stoyanova, D.; Papageorgiou, M.; Rocha, J.M.; Gotcheva, V.; Angelov, A. Effects of Teff-Based Sourdoughs on Dough Rheology and Gluten-Free Bread Quality. Foods 2022, 11, 1012. [Google Scholar] [CrossRef] [PubMed]
- Kaya, Y.; Erten, T.; Vurmaz, M.; İspirli, H.; Şimşek, Ö.; Dertli, E. Comparison of the probiotic characteristics of Lactic Acid Bacteria (LAB) isolated from sourdough and infant feces. Food Biosci. 2022, 47, 101722. [Google Scholar] [CrossRef]
- Bukvicki, D.; Siroli, L.; D’Alessandro, M.; Cosentino, S.; Fliss, I.; Said, L.B.; Hassan, H.; Lanciotti, R.; Patrignani, F. Unravelling the potential of Lactococcus lactis strains to be used in cheesemaking production as biocontrol agents. Foods 2020, 9, 1815. [Google Scholar] [CrossRef]
- Dukare, A.; Paul, S. Biological control of Fusarium wilt and growth promotion in pigeon pea (Cajanus cajan) by antagonistic rhizobacteria, displaying multiple modes of pathogen inhibition. Rhizosphere 2021, 17, 100278. [Google Scholar] [CrossRef]
- Novotni, D.; Gänzle, M.; Rocha, J.M. Composition and Activity of Microbiota in Sourdough and Their Effect on Bread Quality and Safety. In Trends in Wheat and Bread Making; Galanakis, C.M., Ed.; Elsevier-Academic Press: Cambridge, MA, USA, 2020; p. 469. [Google Scholar]
- Yu, A.; Goldman, E.; Brooks, J.; Golomb, B.; Yim, I.; Gotcheva, V.; Angelov, A.; Kim, E.; Marco, M. Strain diversity of plant-associated Lactiplantibacillus plantarum. Microb. Biotechnol. 2021, 14, 1990–2008. [Google Scholar] [CrossRef]
- Rocha, J.M. Microbiological and Lipid Profiles of Broa: Contributions for the Characterization of a Traditional Portuguese Bread. Ph.D. Thesis, Instituto Superior de Agronomia, Universidade de Lisboa, Lisbon, Portugal, 2011; p. 705. [Google Scholar]
- Niku-Paavola, M.; Laitila, L.A.; Mattila-Sandholm, T.; Hikara, A. New types of antimicrobial compounds produced by Lactobacillus plantarum. J. Appl. Microbiol. 1999, 86, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Okkers, D.J.; Dicks, L.M.T.; Silvester, M.; Joubert, J.J.; Odendaal, H.J. Characterization of pentocin TV35b, a bacteriocin-like peptide isolated from Lactobacillus pentosus with a fungistatic effect on Candida albicans. J. Appl. Microbiol. 1999, 87, 726–734. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Cassone, A.; Coda, R.; Gobbetti, M. Antifungal activity of sourdough fermented wheat germ used as an ingredient for bread making. Food Chem. 2011, 127, 952–959. [Google Scholar] [CrossRef] [PubMed]
- Bartkiene, E.; Lele, V.; Starkute, V.; Zavistanaviciute, P.; Zokaityte, E.; Varinauskaite, I.; Pileckaite, G.; Paskeviciute, L.; Rutkauskaite, G.; Kanaporis, T.; et al. Plants and Lactic Acid Bacteria Combination for New Antimicrobial and Antioxidant Properties Product Development in a Sustainable Manner. Foods 2020, 9, 433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karpiński, T.M.; Szkaradkiewicz, A.K. Characteristic of bacteriocines and their application. Pol. J. Microbiol. 2013, 62, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Nes, I.F.; Diep, D.B.; Havarstein, L.S.; Brurberg, M.B.; Eijsink, V.; Holo, H. Biosynthesis of bacteriocins in lactic acid bacteria. Antonie Van Leeuwenhoek 1996, 70, 113–128. [Google Scholar] [CrossRef] [PubMed]
- Diep, D.B.; Havarstein, L.S.; Nes, I.F. Characterization of the locus responsible for the bacteriocin production in Lactobacillus plantarum C11. J. Bacteriol. 1996, 178, 4472–4483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Favaro, L.; Penna, A.L.B.; Todorov, S.D. Bacteriocinogenic LAB from cheeses–application in biopreservation? Trends Food Sci Technol. 2015, 41, 37–48. [Google Scholar] [CrossRef]
- Vescovo, M.; Torriani, S.; Orsi, C.; Macchiarolo, F.; Scolari, G. Application of antimicrobial-producing lactic acid bacteria to control pathogens in ready-to-use vegetables. J. Appl. Microbiol. 1996, 81, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Torriani, S.; Orsi, C.; Vescovo, M. Potential of Lactobacillus casei, culture permeate, and lactic acid to control microorganisms in ready-to-use vegetables. J. Food Prot. 1997, 60, 1564–1567. [Google Scholar] [CrossRef] [PubMed]
- Alegre, I.; Viñas, I.; Usall, J.; Anguera, M.; Altisent, R.; Abadias, M. Antagonistic effect of Pseudomonas graminis CPA-7 against foodborne pathogens in fresh-cut apples under simulated commercial conditions. Food Microbiol. 2013, 33, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Abadias, M.; Altisent, R.; Usall, J.; Torres, R.; Oliveira, M.; Viñas, I. Biopreservation of fresh-cut melon using the strain Pseudomonas graminis CPA-7. Postharvest Biol. Technol. 2014, 96, 69–77. [Google Scholar] [CrossRef]
- Rocha, J.M.; Malcata, F.X. On the microbiological profile of traditional Portuguese sourdough. J. Food Prot. 1999, 62, 1416–1429. [Google Scholar] [CrossRef] [PubMed]
- Zokaityte, E.; Cernauskas, D.; Klupsaite, D.; Lele, V.; Starkute, V.; Zavistanaviciute, P.; Ruzauskas, M.; Gruzauskas, R.; Juodeikiene, G.; Rocha, J.M.; et al. Bioconversion of milk permeate with selected lactic acid bacteria strains and apple by-products into beverages with antimicrobial properties and enriched with galactooligosaccharides. Microorganisms 2020, 8, 1182. [Google Scholar] [CrossRef]
- Todorov, S.D.; Dicks, L.M.T. Effect of medium components on bacteriocin production by Lactobacillus pentosus ST151BR, a strain isolated from beer produced by the fermentation of maize, barley and soy flour. World J. Microbiol. Biotechnol. 2004, 20, 643–650. [Google Scholar] [CrossRef]
- Sood, S.K.; Sinha, P.R. Analysis of structure of YGNGV motif containing bacteriocins: A model for pore formation. Indian J. Biotech. 2003, 2, 227–235. [Google Scholar]
- Klein, M.; Burla, B.; Martinoia, E. The multidrug resistance-associated protein (MRP/ABCC) subfamily of ATP-binding cassette transporters in plants. FEBS Lett. 2006, 580, 1112–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michiels, J.; Dirix, G.; Vanderleyden, J.; Xi, C. Processing and export of peptide pheromones and bacteriocins in Gram-negative bacteria. Trends Microbiol. 2001, 9, 164–168. [Google Scholar] [CrossRef]
- Daeschel, M.A.; McKenney, M.C.; McDonald, L.C. Bacteriocidal activity of Lactobacillus plantarum C-11. Food Microbiol. 1990, 7, 91–98. [Google Scholar] [CrossRef]
- Todorov, S.D.; de Melo Franco, B.D.G.; Tagg, J.R. Bacteriocins of Gram-positive bacteria having activity spectra extending beyond closely-related species. Benef. Microbes 2019, 10, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Fathia, B.A.H.R.I.; Noreddine, K.C. Partial characterization of bacteriocin-like substance produced by probiotic Lactobacillus plantarum F12 isolated from Algerian children faeces. Afr. J. Microbiol. Res. 2016, 10, 1798–1805. [Google Scholar] [CrossRef] [Green Version]
- Lavilla-Lerma, L.; Pérez-Pulido, R.; Martínez-Bueno, M.; Maqueda, M.; Valdivia, E. Characterization of functional, safety, and gut survival related characteristics of Lactobacillus strains isolated from farmhouse goat’s milk cheeses. Int. J. Food Microbiol. 2013, 163, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Grinter, R.; Milner, J.; Walker, D. Bacteriocins active against plant pathogenic bacteria. Biochem. Soc. Trans. 2012, 40, 1498–1502. [Google Scholar] [CrossRef] [PubMed]
- Rooney, W.M.; Grinter, R.W.; Correia, A.; Parkhill, J.; Walker, D.C.; Milner, J.J. Engineering bacteriocin-mediated resistance against the plant pathogen Pseudomonas syringae. Plant Biotechnol. J. 2019, 18, 1296–1306. [Google Scholar]
- Crowley, S.; Mahony, J.; van Sinderen, D. Broad-spectrum antifungal-producing lactic acid bacteria and their application in fruit models. Folia Microbiol. 2012, 58, 291–299. [Google Scholar] [CrossRef]
- Agriopoulou, S.; Stamatelopoulou, E.; Sachadyn-Król, M.; Varzakas, T. Lactic acid bacteria as antibacterial agents to extend the shelf life of fresh and minimally processed fruits and vegetables: Quality and safety aspects. Microorganisms 2020, 8, 952. [Google Scholar] [CrossRef]
- Küley, E.; Özyurt, G.; Özogul, I.; Boga, M.; Akyol, I.; Rocha, J.M.; Özogul, F. The Role of Selected Lactic Acid Bacteria on Organic Acid Accumulation during Wet and Spray-Dried Fish-based Silages. Contributions to the Winning Combination of Microbial Food Safety and Environmental Sustainability. Microorganisms 2020, 8, 172. [Google Scholar] [CrossRef] [Green Version]
- Bultreys, A.; Kaluzna, M. Bacterial cankers caused by Pseudomonas syringae on stone fruit species with special emphasis on the pathovars syringae and morsprunorum race 1 and race 2. J. Plant Pathol. 2010, 92, 21–33. [Google Scholar]
- King, E.O.; Ward, M.K.; Raney, D.E. Two simple media for the demonstration of pyocyanin and fluorescin. J. Lab. Clin. Med. 1954, 44, 301–307. [Google Scholar]
- Torriani, S.; Felis, G.E.; Dellaglio, F. Differentiation of Lactobacillus plantarum, L. pentosus, and L. paraplantarum by recA gene sequence analysis and multiplex PCR assay with recA gene-derived primers. Appl. Environ. Microbiol. 2001, 67, 3450–3454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Angelis, M.; Siragusa, S.; Berloco, M.; Caputo, L.; Settanni, L.; Alfonsi, G.; Grandi, A.; Ragni, A.; Gobbetti, M. Selection of potential probiotic lactobacilli from pig feces to be used as additives in pelleted feeding. Res. Microbiol. 2006, 157, 792–801. [Google Scholar] [CrossRef] [PubMed]
- Siragusa, S.; Di Cagno, R.; Ercolini, D.; Minervini, F.; Gobbetti, M.; De Angelis, M. Taxonomic structure and monitoring of the dominant population of lactic acid bacteria during wheat flour sourdough type I propagation using Lactobacillus sanfranciscensis starters. Appl. Environ. Microbiol. 2009, 75, 1099–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehrmann, M.T.; Vogel, R.F. Molecular taxonomy and genetics of sourdough lactic acid bacteria. Trends Food Sci. Technol. 2005, 16, 31–42. [Google Scholar] [CrossRef]
- Omar, N.B.; Abriouel, H.; Lucas, R.; Martinez-Canamero, M.; Guyot, J.P.; Galvez, A. Bacteriocin-producing Lactobacillus strains isolated from poto poto, a Congolese fermented maize product, and genetic fingerprinting of their plantaricin operons. Int. J. Food Microbiol. 2008, 127, 18–25. [Google Scholar]
- Pal, G.; Srivastava, S. Inhibitory effect of plantaricin peptides (Pln E/F and J/K) against Escherichia coli. World J. Microbiol. Biotechnol. 2014, 30, 2829–2837. [Google Scholar] [CrossRef]
- Maldonado, A.; Ruiz-Barba, J.L.; Floriano, B.; Jiménez-Díaz, R. The locus responsible for production of plantaricin S, a class IIb bacteriocin produced by Lactobacillus plantarum LPCO10, is widely distributed among wild-type Lact. plantarum strains isolated from olive fermentations. Int. J. Food Microbiol. 2002, 77, 117–124. [Google Scholar] [CrossRef]
- Hall, S.J.; Dry, I.B.; Blanchard, C.L.; Whitelaw-Weckert, M.A. Phylogenetic relationships of Pseudomonas syringae pv. syringae isolates associated with bacterial inflorescence rot in grapevine. Plant Dis. 2016, 100, 607–616. [Google Scholar] [CrossRef] [Green Version]
- Ogunremi, O.R.; Leischtfeld, S.F.; Mischler, S.; Schwenninger, S.M. Antifungal activity of lactic acid bacteria isolated from kunu-zaki, a cereal-based Nigerian fermented beverage. Food Biosci. 2022, 49, 101648. [Google Scholar] [CrossRef]
- Tryfinopoulou, P.; Kizis, D.; Nychas, G.J.E.; Panagou, E.Z. Quantification of Aspergillus carbonarius in grapes using a real time PCR assay. Food Microbiol. 2015, 51, 139–143. [Google Scholar] [CrossRef]
- Chen, C.; Zhang, X.; Wei, X.; Zhu, Y.; Chen, W.; Han, Y. Postharvest biological control of Botrytis cinerea and the mechanisms underlying the induction of disease resistance in grapes by Lactobacillus plantarum CM-3. Biol. Control 2022, 172, 104982. [Google Scholar] [CrossRef]
- Kasfi, K.; Taheri, P.; Jafarpour, B.; Tarighi, S. Identification of epiphytic yeasts and bacteria with potential for biocontrol of grey mold disease on table grapes caused by Botrytis cinerea. Span. J. Agric. Res. 2018, 16, 23. [Google Scholar] [CrossRef]
- De Vuyst, L.; Schrijvers, V.; Paramithiotis, S.; Hoste, B.; Vancanneyt, M.; Swings, J.; Messens, W. The biodiversity of lactic acid bacteria in Greek traditional wheat sourdoughs is reflected in both composition and metabolite formation. Appl. Environ. Microbiol. 2002, 68, 6059–6069. [Google Scholar] [CrossRef] [Green Version]
- Minervini, F.; Di Cagno, R.; Lattanzi, A.; De Angelis, M.; Antonielli, L.; Cardinali, G.; Gobbetti, M. Lactic acid bacterium and yeast microbiotas of 19 sourdoughs used for traditional/typical Italian breads: Interactions between ingredients and microbial species diversity. Appl. Environ. Microbiol. 2012, 78, 1251–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banwo, K.; Sanni, A.; Tan, H.; Tian, Y. Phenotypic and genotypic characterization of lactic acid bacteria isolated from some Nigerian traditional fermented foods. Food Biotechnol. 2012, 26, 124–142. [Google Scholar] [CrossRef]
- Diep, D.B.; Myhre, R.; Johnsborg, O.; Aakra, Å.; Nes, I.F. Inducible bacteriocin production in Lactobacillus is regulated by differential expression of the pln operons and by two antagonizing response regulators, the activity of which is enhanced upon phosphorylation. Molecular Microbiology 2003, 47, 483–494. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; Leroy, F. Bacteriocins from lactic acid bacteria: Production, purification, and food applications. J. Mol. Microbiol. Biotechnol. 2007, 13, 194–199. [Google Scholar] [CrossRef]
- Todorov, S.D. Bacteriocins from Lactobacillus plantarum production, genetic organization and mode of action: Braz. J. Microbiol. 2009, 40, 209–221. [Google Scholar]
- Brurberg, M.B.; Nes, I.F.; Eijsink, V.G.H. Pheromone-induced production of antimicrobial peptides in Lactobacillus. Mol. Microbiol. 1997, 26, 347–360. [Google Scholar] [CrossRef]
- Rojo-Bezares, B.; Sáenz, Y.; Navarro, L.; Jiménez-Díaz, R.; Zarazaga, M.; Ruiz-Larrea, F.; Torres, C. Characterization of a new organization of the plantaricin locus in the inducible bacteriocin-producing Lactobacillus plantarum J23 of grape must origin. Arch. Microbiol. 2008, 189, 491–499. [Google Scholar] [CrossRef]
- Holo, H.; Jeknic, Z.; Daeschel, M.; Stevanovic, S.; Nes, I.F. Plantaricin W from Lactobacillus plantarum belongs to a new family of two-peptide lantibiotics. Microbiology 2001, 147, 643–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haghshenas, B.; Abdullah, N.; Nami, Y.; Radiah, D.; Rosli, R.; Khosroushahi, A.Y. Different effects of two newly-isolated probiotic Lactobacillus plantarum 15HN and Lactococcus lactis subsp. lactis 44 Lac strains from traditional dairy products on cancer cell lines. Anaerobe 2014, 30, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, I.; Kabadjova, P.; Pantev, A.; Danova, S.; Dousset, X. Detection, purification and partial characterization of a novel bacteriocin substance produced by Lactococcus lactis subsp. lactis B14 isolated from boza-Bulgarian traditional cereal beverage. Biocatalysis 2000, 41, 47–53. [Google Scholar]
- Todorov, S.; Onno, B.; Sorokine, O.; Chobert, J.M.; Ivanova, I.; Dousset, X. Detection and characterization of a novel antibacterial substance produced by Lactobacillus plantarum ST 31 isolated from sourdough. Int. J. Food Microbiol. 1999, 48, 167–177. [Google Scholar] [CrossRef]
- Todorov, S.D.; Dicks, L.M. Characterization of bacteriocins produced by lactic acid bacteria isolated from spoiled black olives. J. Basic Microbiol. 2005, 45, 312–322. [Google Scholar] [CrossRef]
- von Mollendorff, J.W.; Todorov, S.D.; Dicks, L.M. Comparison of bacteriocins produced by lactic-acid bacteria isolated from boza, a cereal-based fermented beverage from the Balkan Peninsula. Curr. Microbiol. 2006, 53, 209–216. [Google Scholar] [CrossRef]
- Lavermicocca, P.; Valerio, F.; Evidente, A.; Lazzaroni, S.; Corsetti, A.; Gobetti, M. Purification and characterization of Novel antifungal compounds from the sourdough Lactobacillus plantarum strain 21B. Appl. Environ. Microbiol. 2000, 66, 4084–4090. [Google Scholar] [CrossRef] [Green Version]
- Ogunbanwo, S.T.; Sanni, A.I.; Onilude, A.A. Characterization of bacteriocin produced by Lactobacillus plantarum F1 and Lactobacillus brevis OG1. Afr. J. Biotechnol. 2003, 2, 219–227. [Google Scholar]
- Zamani-Zadeh, M.; Soleimanian-Zad, S.; Sheikh-Zeinoddin, M.; Goli, S.A.H. Integration of Lactobacillus plantarum A7 with thyme and cumin essential oils as a potential biocontrol tool for gray mold rot on strawberry fruit. Postharvest Biol. Technol. 2014, 92, 149–156. [Google Scholar] [CrossRef]
- Dopazo, V.; Luz, C.; Quiles, J.M.; Calpe, J.; Romano, R.; Mañes, J.; Meca, G. Potential application of lactic acid bacteria in the biopreservation of red grape from mycotoxigenic fungi. J. Sci. Food Agric. 2022, 102, 898–907. [Google Scholar] [CrossRef]
- Marín, A.; Plotto, A.; Atarés, L.; Chiralt, A. Lactic acid bacteria incorporated into edible coatings to control fungal growth and maintain postharvest quality of grapes. HortScience 2019, 54, 337–343. [Google Scholar] [CrossRef] [Green Version]
- Tsuda, K.; Tsuji, G.; Higashiyama, M.; Ogiyama, H.; Umemura, K.; Mitomi, M.; Kubo, Y.; Kosaka, Y. Biological control of bacterial soft rot in Chinese cabbage by Lactobacillus plantarum strain BY under field conditions. Biol. Control 2016, 100, 63–69. [Google Scholar] [CrossRef]
- Crowley, S.; Mahony, J.; Van Sinderen, D. Comparative analysis of two antifungal Lactobacillus plantarum isolates and their application as bioprotectants in refrigerated foods. J. Appl. Microbiol. 2012, 113, 1417–1427. [Google Scholar] [CrossRef]
- Wang, H.; Yan, Y.; Wang, J.; Zhang, H.; Qi, W. Production and characterization of antifungal compounds produced by Lactobacillus plantarum IMAU10014. PLoS ONE 2012, 7, e29452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, W.T.; Chen, Y.S.; Wu, H.C.; Yanagida, F. Bio-protective potential of lactic acid bacteria isolated from fermented wax gourd. Folia Microbiol. 2012, 5, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Cao, Z.; Li, J.; Tao, C.; Feng, Y.; Han, Y. A novel endophytic strain of Lactobacillus plantarum CM-3 with antagonistic activity against Botrytis cinerea on strawberry fruit. Biol. Control 2020, 148, 104306. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteriocins/ Lp. plantarum Strains | C6 | D16 | I6 | K1 | L1 | Q3 | Q4 | S1 | S4 |
|---|---|---|---|---|---|---|---|---|---|
| plnA | + | ND | ND | + | + | + | + | + | + |
| plnB | + | + | + | + | + | + | + | + | + |
| plnC | + | ND | + | + | ND | + | + | + | + |
| plnD | + | + | + | + | + | + | + | + | + |
| plnEF | + | + | + | + | + | + | + | + | + |
| plnG | + | + | + | + | + | + | + | + | + |
| plnI | + | + | + | + | + | + | + | + | + |
| plnJ | + | + | ND | + | + | + | + | + | + |
| plnK | + | ND | ND | ND | + | + | ND | + | + |
| plnN | + | + | + | + | + | + | + | + | + |
| plnNC8 | + | + | + | + | + | + | + | ND | + |
| plnS | ND | + | + | + | + | + | + | ND | + |
| plnW | ND | + | ND | ND | ND | ND | ND | ND | ND |
| LAB Strain | Treatment | |||||
|---|---|---|---|---|---|---|
| CFS | Trypsin | pH | Proteinase K | Catalase | Boiling | |
| Inhibition Zone, mm | ||||||
| C6 | 16.00 ± 1.02 d** | 3.10 ± 0.79 c | 1.17 ± 0.36 b | 0.00 a | 0.00 a | 3.03 ± 0.33 c** |
| D16 | 23.67 ± 0.93 d** | 0.00 a | 0.00 a | 0.00 a | 18.00 ± 1.36 d** | 0.00 a |
| I6 | 16.00 ± 0.85 d** | 0.00 a | 2.03 ± 0.26 d** | 1.67 ± 0.59 b | 0.00 a | 0.00 a |
| L1 | 12.17 ± 0.76 c | 26.67 ± 0.92 e** | 0.00 a | 0.00 a | 1.33 ± 0.23 b | 0.00 a |
| K1 | 2.33 ± 0.65 a | 0.00 a | 1.63 ± 0.37 c | 0.00 a | 0.00 a | 0.00 a |
| Q3 | 15.67 ± 0.83 d** | 5.33 ± 0.46 d** | 1.77 ± 0.59 cd** | 2.50 ± 0.74 c** | 0.00 a | 0.00 a |
| Q4 | 23.67 ± 0.98 e** | 3.17 ± 0.85 c | 2.53 ± 0.83 e** | 0.00 a | 2.33 ± 0.47 c** | 1.90 ± 0.42 b** |
| S1 | 5.80 ± 0.57 b | 2.00 ± 0.45 b | 2.00 ± 0.72 d** | 0.00 a | 2.23 ± 0.49 c** | 0.00 a |
| S4 | 1.70 ± 0.32 a | 0.00 a | 0.00 a | 1.330 ± 0.46 b | 1.70 ± 0.39 bc | 0.00 a |
| LAB Strain | Treatment | |||||
|---|---|---|---|---|---|---|
| CFS | Trypsin | pH | Proteinase K | Catalase | Boiling | |
| Mycelial Growth, % | ||||||
| C6 | 81.62 ± 1.32 d | 45.86 ± 2.15 a** | 72.87 ± 1.26 c | 82.04 ± 0.99 d** | 68.53 ± 1.28 c** | 49.47 ± 1.02 b** |
| D16 | 83.34 ± 1.50 d | 81.94 ± 0.61 d | 74.37 ± 1.76 c | 83.04 ± 0.58 d | 80.93 ± 0.93 cd | 46.45 ± 0.97 ab |
| I6 | 82.04 ± 1.93 d | 64.65 ± 1.00 d | 72.51 ± 0.87 c | 71.33 ± 1.27 c | 79.28 ± 0.63 cd | 62.29 ± 1.38 ab |
| L1 | 82.22 ± 0.57 d | 82.51 ± 1.26 d | 68.51 ± 2.01 cd** | 52.43 ± 0.72 b | 45.86 ± 0.98 b** | 70.58 ± 0.88 c |
| K1 | 79.61 ± 1.30 cd** | 55.24 ± 2.04 b | 64.11 ± 1.52 c | 71.48 ± 1.55 c | 78.12 ± 1.30 cd | 72.09 ± 1.12 c |
| Q3 | 82.87 ± 0.81 d | 78.37 ± 1.99 cd** | 58.97 ± 0.73 b** | 60.35 ± 0.73 bc** | 65.14 ± 1.22 c** | 30.31 ± 0.75 a** |
| Q4 | 83.42 ± 1.28 d | 52.24 ± 1.63 b** | 67.57 ± 0.65 c | 31.93 ± 1.28 a | 73.06 ± 0.96 c** | 30.15 ± 1.54 a** |
| S1 | 80.93 ± 0.668 d | 66.35 ± 2.04 cd** | 79.62 ± 1.00 cd** | 23.75 ± 1.49 a | 52.35 ± 1.66 b** | 49.45 ± 0.73 b** |
| S4 | 81.21 ± 1.53 d | 82.92 ± 0.94 d | 71.82 ± 0.92 c | 69.28 ± 0.94 bc** | 68.45 ± 1.11 bc** | 73.36 ± 1.62 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petkova, M.; Gotcheva, V.; Dimova, M.; Bartkiene, E.; Rocha, J.M.; Angelov, A. Screening of Lactiplantibacillus plantarum Strains from Sourdoughs for Biosuppression of Pseudomonas syringae pv. syringae and Botrytis cinerea in Table Grapes. Microorganisms 2022, 10, 2094. https://doi.org/10.3390/microorganisms10112094
Petkova M, Gotcheva V, Dimova M, Bartkiene E, Rocha JM, Angelov A. Screening of Lactiplantibacillus plantarum Strains from Sourdoughs for Biosuppression of Pseudomonas syringae pv. syringae and Botrytis cinerea in Table Grapes. Microorganisms. 2022; 10(11):2094. https://doi.org/10.3390/microorganisms10112094
Chicago/Turabian StylePetkova, Mariana, Velitchka Gotcheva, Milena Dimova, Elena Bartkiene, João Miguel Rocha, and Angel Angelov. 2022. "Screening of Lactiplantibacillus plantarum Strains from Sourdoughs for Biosuppression of Pseudomonas syringae pv. syringae and Botrytis cinerea in Table Grapes" Microorganisms 10, no. 11: 2094. https://doi.org/10.3390/microorganisms10112094