
Vaginal Lactobacillus Impair Candida Dimorphic Switching and Biofilm Formation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Lactobacilli Culture Conditions and Preparation of Planktonic/Biofilm Cell-Free Supernatants
2.2. Candida spp. Culture Conditions and Treatment with Lactobacilli Supernatants
2.3. Evaluation of Candida Dimorphic Switching
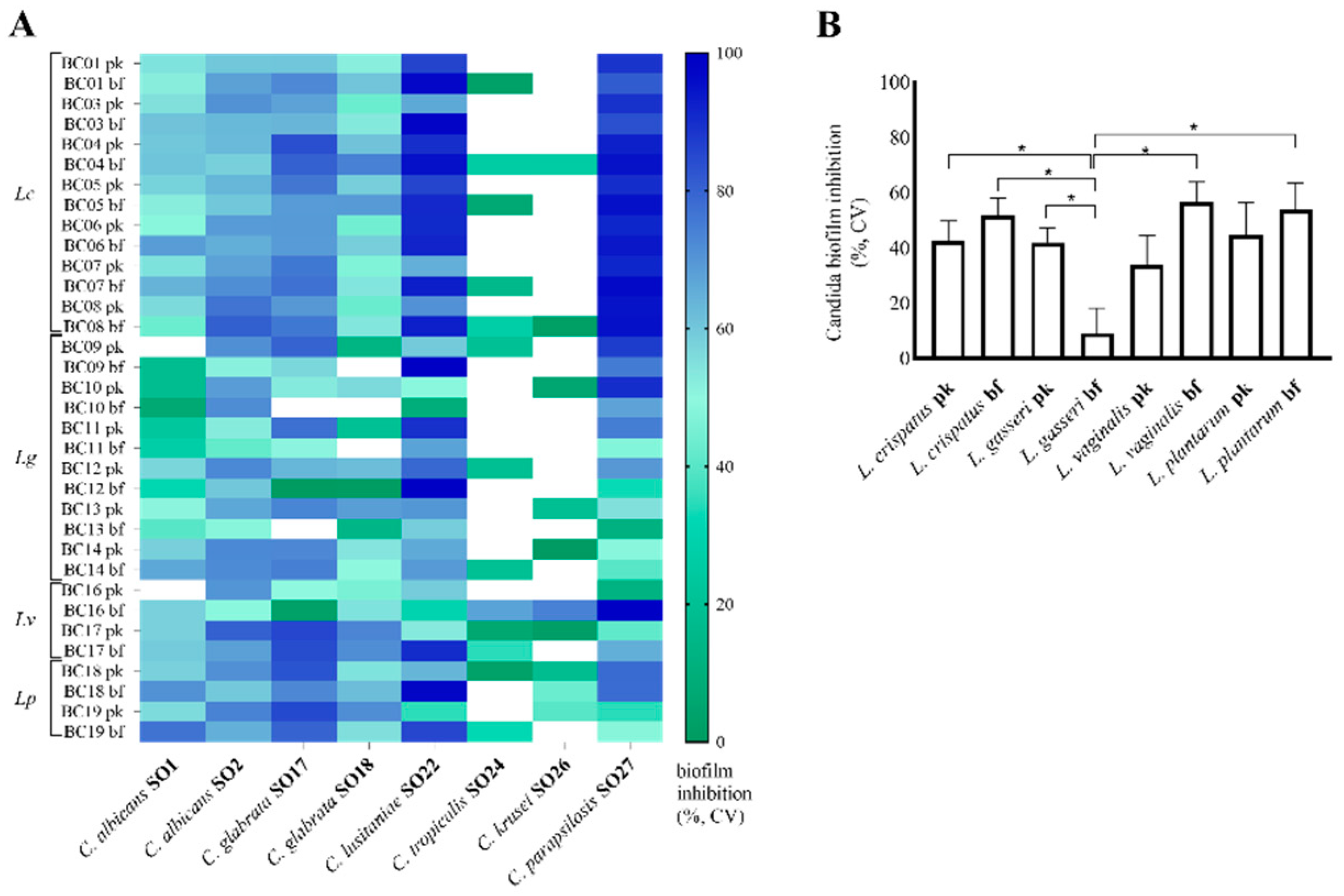
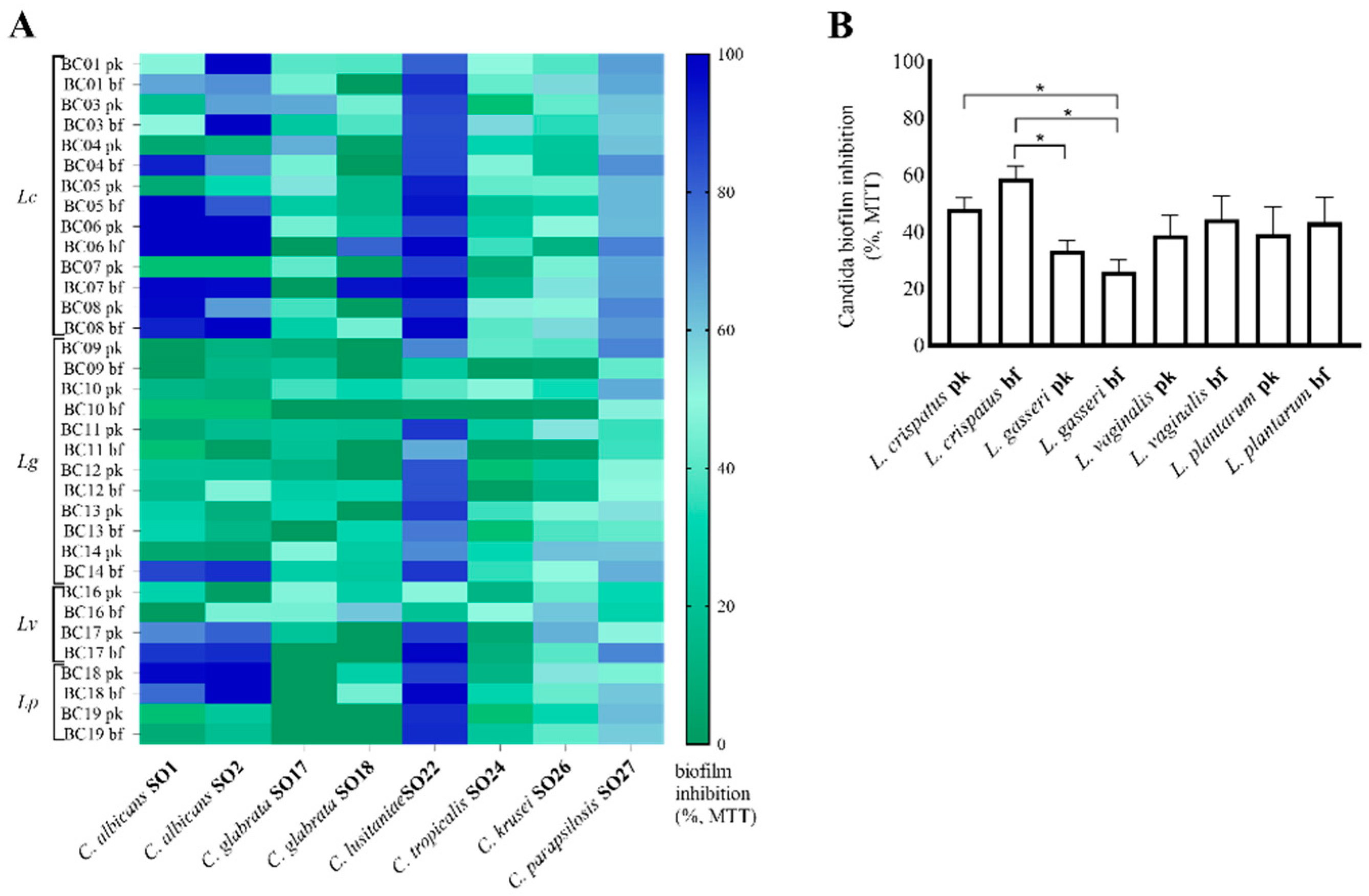
2.4. Evaluation of Candida Biofilms
2.4.1. Candida Biofilm Quantification by Crystal Violet Staining
2.4.2. Candida Biofilm Quantification by MTT Assay
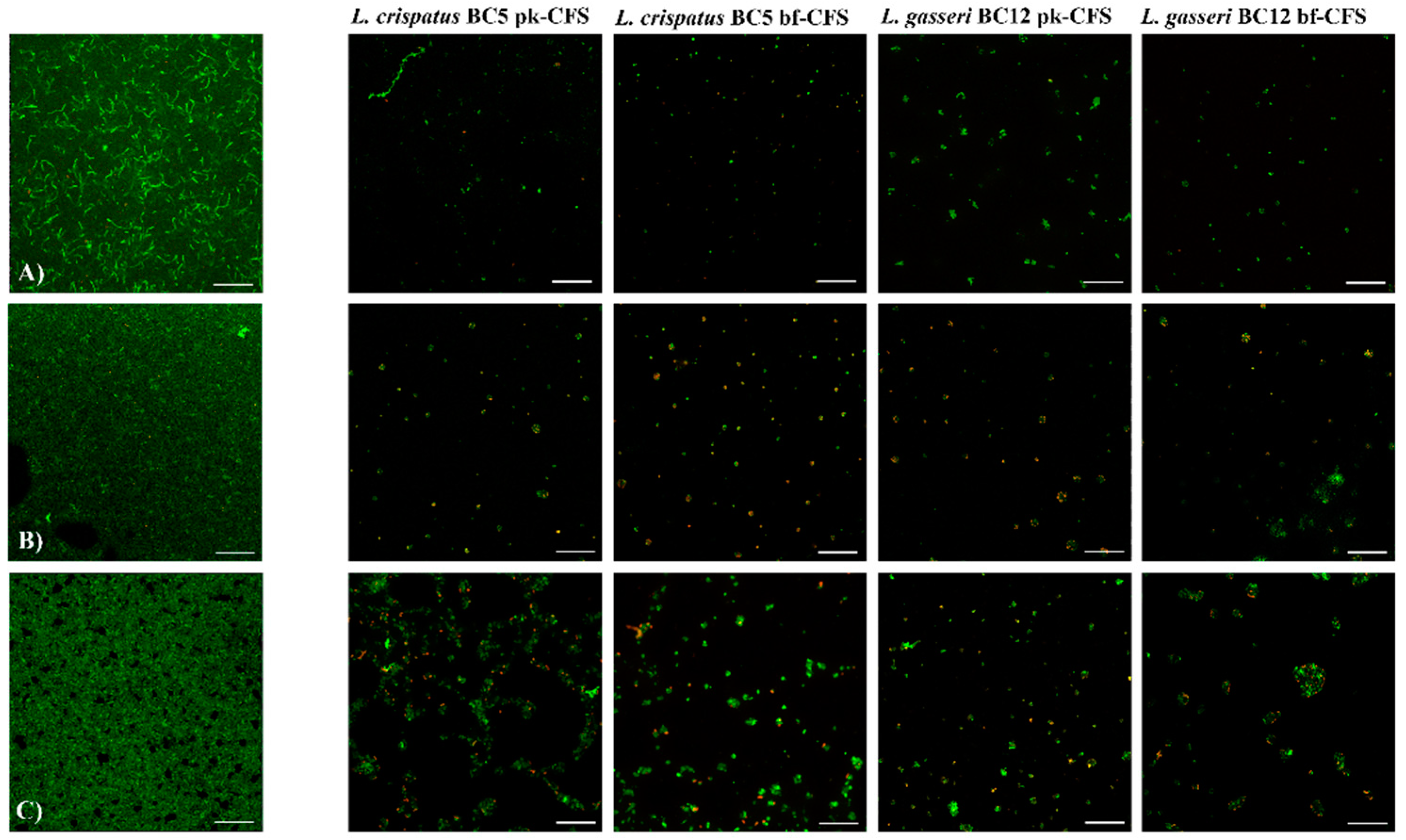
2.4.3. Candida Biofilm Visualization by Confocal Microscopy
2.5. Data Analysis
3. Results
3.1. Lactobacilli CFS Impair Candida Dimorphic Switching
3.2. Lactobacilli CFS Impact on Candida Biofilms
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nobile, C.J.; Johnson, A.D. Candida albicans Biofilms and Human Disease. Annu. Rev. Microbiol. 2015, 69, 71–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willems, H.M.E.; Ahmed, S.S.; Liu, J.; Xu, Z.; Peters, B.M. Vulvovaginal Candidiasis: A Current Understanding and Burning Questions. J. Fungi 2020, 6, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Cerdeira, C.; Gregorio, M.C.; Molares-Vila, A.; López-Barcenas, A.; Fabbrocini, G.; Bardhi, B.; Sinani, A.; Sánchez-Blanco, E.; Arenas-Guzmán, R.; Hernandez-Castro, R. Biofilms and vulvovaginal candidiasis. Colloids Surf. B. Biointerfaces 2019, 174, 110–125. [Google Scholar] [CrossRef] [PubMed]
- Blostein, F.; Levin-Sparenberg, E.; Wagner, J.; Foxman, B. Recurrent vulvovaginal candidiasis. Ann. Epidemiol. 2017, 27, 575–582.e3. [Google Scholar] [CrossRef]
- Ardizzoni, A.; Wheeler, R.T.; Pericolini, E. It Takes Two to Tango: How a Dysregulation of the Innate Immunity, Coupled with Candida Virulence, Triggers VVC Onset. Front. Microbiol. 2021, 12, 692491. [Google Scholar] [CrossRef]
- Atiencia-Carrera, M.B.; Cabezas-Mera, F.S.; Tejera, E.; Machado, A. Prevalence of biofilms in Candida spp. bloodstream infections: A meta-analysis. PLoS ONE 2022, 17, e0263522. [Google Scholar] [CrossRef]
- Tsui, C.; Kong, E.F.; Jabra-Rizk, M.A. Pathogenesis of Candida albicans biofilm. Pathog. Dis. 2016, 74, ftw018. [Google Scholar] [CrossRef] [Green Version]
- Kean, R.; Delaney, C.; Rajendran, R.; Sherry, L.; Metcalfe, R.; Thomas, R.; McLean, W.; Williams, C.; Ramage, G. Gaining insights from Candida biofilm heterogeneity: One size does not fit all. J. Fungi 2018, 4, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tulasidas, S.; Rao, P.; Bhat, S.; Manipura, R. A study on biofilm production and antifungal drug resistance among Candida species from vulvovaginal and bloodstream infections. Infect. Drug Resist. 2018, 11, 2443–2448. [Google Scholar] [CrossRef] [Green Version]
- Sobel, J.D. Editorial Commentary: Vaginal Biofilm: Much Ado about Nothing, or a New Therapeutic Challenge? Clin. Infect. Dis. 2015, 61, 607–608. [Google Scholar] [CrossRef]
- Jacobsen, I.D.; Wilson, D.; Wächtler, B.; Brunke, S.; Naglik, J.R.; Hube, B. Candida albicans dimorphism as a therapeutic target. Expert Rev. Anti. Infect. Ther. 2012, 10, 85–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borges, S.; Silva, J.; Teixeira, P. The role of lactobacilli and probiotics in maintaining vaginal health. Arch. Gynecol. Obstet. 2014, 289, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Pacha-Herrera, D.; Erazo-Garcia, M.P.; Cueva, D.F.; Orellana, M.; Borja-Serrano, P.; Arboleda, C.; Tejera, E.; Machado, A. Clustering Analysis of the Multi-Microbial Consortium by Lactobacillus Species Against Vaginal Dysbiosis Among Ecuadorian Women. Front. Cell. Infect. Microbiol. 2022, 12, 863208. [Google Scholar] [CrossRef] [PubMed]
- Petrova, M.I.; Lievens, E.; Malik, S.; Imholz, N.; Lebeer, S. Lactobacillus species as biomarkers and agents that can promote various aspects of vaginal health. Front. Physiol. 2015, 6, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parolin, C.; Marangoni, A.; Laghi, L.; Foschi, C.; Palomino, R.A.Ñ.; Calonghi, N.; Cevenini, R.; Vitali, B. Isolation of vaginal lactobacilli and characterization of anti-candida activity. PLoS ONE 2015, 10, e0131220. [Google Scholar] [CrossRef]
- Parolin, C.; Abruzzo, A.; Giordani, B.; Oliver, J.C.; Marangoni, A.; Luppi, B.; Vitali, B. Anti-candida activity of hyaluronic acid combined with lactobacillus crispatus lyophilised supernatant: A new antifungal strategy. Antibiotics 2021, 10, 628. [Google Scholar] [CrossRef]
- De Gregorio, P.R.; Parolin, C.; Abruzzo, A.; Luppi, B.; Protti, M.; Mercolini, L.; Silva, J.A.; Giordani, B.; Marangoni, A.; Nader-Macías, M.E.F.; et al. Biosurfactant from vaginal Lactobacillus crispatus BC1 as a promising agent to interfere with Candida adhesion. Microb. Cell Fact. 2020, 19, 133. [Google Scholar] [CrossRef]
- Abruzzo, A.; Giordani, B.; Parolin, C.; Vitali, B.; Protti, M.; Mercolini, L.; Cappelletti, M.; Fedi, S.; Bigucci, F.; Cerchiara, T.; et al. Novel mixed vesicles containing lactobacilli biosurfactant for vaginal delivery of an anti- Candida agent. Eur. J. Pharm. Sci. 2018, 112, 95–101. [Google Scholar] [CrossRef]
- Abruzzo, A.; Giordani, B.; Parolin, C.; De Gregorio, P.R.; Foschi, C.; Cerchiara, T.; Bigucci, F.; Vitali, B.; Luppi, B. Lactobacillus crispatus BC1 biosurfactant delivered by hyalurosomes: An advanced strategy to counteract candida biofilm. Antibiotics 2021, 10, 33. [Google Scholar] [CrossRef]
- Parolin, C.; Croatti, V.; Laghi, L.; Giordani, B.; Tondi, M.R.; De Gregorio, P.R.; Foschi, C.; Vitali, B. Lactobacillus Biofilms Influence Anti-Candida Activity. Front. Microbiol. 2021, 12, 1–11. [Google Scholar] [CrossRef]
- Oliver, J.C.; Laghi, L.; Parolin, C.; Foschi, C.; Marangoni, A.; Liberatore, A.; Dias, A.L.T.; Cricca, M.; Vitali, B. Metabolic profiling of Candida clinical isolates of different species and infection sources. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- EUCAST Committee. Method for the determination of broth dilution minimum inhibitory concentrations of antifungal agents for yeasts. EUCAST Defin. Doc. E.DEF 7.3.2 2020.
- Rutering, J.; Ilmer, M.; Recio, A.; Coleman, M.; Vykoukal, J.; Alt, E.; Orleans, N. Quantitative and Qualitative Assessment Methods for Biofilm Growth: A Mini-review Christina. Nat. Rev. Drug Discov. 2016, 5, 1–8. [Google Scholar]
- Aballéa, S.; Guelfucci, F.; Wagner, J.; Khemiri, A.; Dietz, J.P.; Sobel, J.; Toumi, M. Subjective health status and health-related quality of life among women with Recurrent Vulvovaginal Candidosis (RVVC) in Europe and the USA. Health Qual. Life Outcomes 2013, 11, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, C.; Li, H.; Han, L.; Wang, C.; Yan, Y.; Qi, W.; Fan, A.; Wang, Y.; Xue, F. Aerobic vaginitis in late pregnancy and outcomes of pregnancy. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 233–239. [Google Scholar] [CrossRef]
- Dover, S.E.; Aroutcheva, A.A.; Faro, S.; Chikindas, M.L. Natural antimicrobials and their role in vaginal health: A short review. Int. J. Probiotics Prebiotics 2008, 3, 219–230. [Google Scholar]
- Fanning, S.; Mitchell, A.P. Fungal biofilms. PLoS Pathog. 2012, 8, e1002585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, K.F.; Zarnowski, R.; Andes, D.R. Fungal Super Glue: The Biofilm Matrix and Its Composition, Assembly, and Functions. PLoS Pathog. 2016, 12, e1005828. [Google Scholar] [CrossRef] [Green Version]
- Desai, J.V. Candida albicans hyphae: From growth initiation to invasion. J. Fungi 2018, 4, 10. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Guo, S.; Chen, X.; Yang, S.; Deng, X.; Tu, M.; Tao, Y.; Xiang, W.; Rao, Y. Metabolic profiles of Lactobacillus paraplantarum in biofilm and planktonic states and investigation of its intestinal modulation and immunoregulation in dogs. Food Funct. 2021, 12, 5317–5332. [Google Scholar] [CrossRef]
- Lohsea, M.; Megha, G.; Arevalo, A.V.; Fishburn, A.; Johnson, A.; Nobile, C. Assessment and Optimizations of Candida albicans In Vitro Biofilm Assays. Antimicrob. Agents Chemother. 2017, 61, 1–13. [Google Scholar] [CrossRef] [Green Version]
- O’Hanlon, D.E.; Moench, T.R.; Cone, R.A. Vaginal pH and microbicidal lactic acid when lactobacilli dominate the microbiota. PLoS ONE 2013, 8, e80074. [Google Scholar] [CrossRef] [PubMed]
- De Gregorio, P.; Silva, J.; Marchesi, A.; Nader-Macías, M. Anti-Candida activity of beneficial vaginal lactobacilli in in vitro assays and in a murine experimental model. FEMS Yeast Res. 2019, 19, foz008. [Google Scholar] [CrossRef] [PubMed]
- Parolin, C.; Frisco, G.; Foschi, C.; Giordani, B.; Salvo, M.; Vitali, B.; Marangoni, A.; Calonghi, N. Lactobacillus crispatus BC5 interferes with Chlamydia trachomatis infectivity through integrin modulation in cervical cells. Front. Microbiol. 2018, 9, 2630. [Google Scholar] [CrossRef] [PubMed]
- Argentini, C.; Fontana, F.; Alessandri, G.; Lugli, G.A.; Mancabelli, L.; Ossiprandi, M.C.; van Sinderen, D.; Ventura, M.; Milani, C.; Turroni, F. Evaluation of Modulatory Activities of Lactobacillus crispatus Strains in the Context of the Vaginal Microbiota. Microbiol. Spectr. 2022, 10, 1–18. [Google Scholar] [CrossRef]
- Salas-Jara, M.; Ilabaca, A.; Vega, M.; García, A. Biofilm Forming Lactobacillus: New Challenges for the Development of Probiotics. Microorganisms 2016, 4, 35. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C. albicans SO1 | C. albicans SO2 | C. lusitaniae SO22 | C. tropicalis SO24 | C. krusei SO26 | C. parapsilosis SO27 | ||
|---|---|---|---|---|---|---|---|
| L. crispatus | BC1pk | + | + | + | - | + | - |
| BC1bf | + | + | + | - | + | + | |
| BC3pk | + | - | - | - | - | - | |
| BC3bf | + | + | + | - | - | + | |
| BC4pk | + | + | - | - | + | - | |
| BC4bf | + | + | + | - | + | + | |
| BC5pk | + | + | + | - | + | - | |
| BC5bf | + | + | + | - | + | + | |
| BC6pk | + | + | + | - | + | - | |
| BC6bf | + | + | + | - | + | + | |
| BC7pk | + | + | - | - | + | - | |
| BC7bf | + | + | - | - | + | + | |
| BC8pk | + | - | - | - | - | - | |
| BC8bf | + | + | + | - | - | + | |
| L. gasseri | BC9pk | + | + | + | - | - | + |
| BC9bf | - | - | - | - | - | - | |
| BC1pk | + | + | + | - | + | - | |
| BC1bf | + | + | + | - | + | - | |
| BC11pk | + | + | - | - | - | - | |
| BC11bf | + | + | - | - | + | - | |
| BC12pk | + | + | - | - | - | - | |
| BC12bf | + | + | - | - | - | - | |
| BC13pk | + | - | + | - | - | - | |
| BC13bf | + | - | + | - | - | - | |
| BC14pk | + | + | - | - | + | - | |
| BC14bf | + | + | - | - | - | - | |
| L. vaginalis | BC16pk | - | - | - | - | - | - |
| BC16bf | - | - | - | - | - | - | |
| BC17pk | - | + | - | - | + | - | |
| BC17bf | - | + | - | - | - | - | |
| L. plantarum | BC18pk | + | + | - | - | + | - |
| BC18bf | + | + | - | - | + | + | |
| BC19pk | - | + | - | - | + | - | |
| BC19bf | + | + | - | - | - | + | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parolin, C.; Croatti, V.; Giordani, B.; Vitali, B. Vaginal Lactobacillus Impair Candida Dimorphic Switching and Biofilm Formation. Microorganisms 2022, 10, 2091. https://doi.org/10.3390/microorganisms10102091
Parolin C, Croatti V, Giordani B, Vitali B. Vaginal Lactobacillus Impair Candida Dimorphic Switching and Biofilm Formation. Microorganisms. 2022; 10(10):2091. https://doi.org/10.3390/microorganisms10102091
Chicago/Turabian StyleParolin, Carola, Vanessa Croatti, Barbara Giordani, and Beatrice Vitali. 2022. "Vaginal Lactobacillus Impair Candida Dimorphic Switching and Biofilm Formation" Microorganisms 10, no. 10: 2091. https://doi.org/10.3390/microorganisms10102091