
Translocation and Dissemination of Gut Bacteria after Severe Traumatic Brain Injury
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice and Modeling
2.1.1. Mice and Housing
2.1.2. Animal Model
2.1.3. Sample Collection
2.2. Protein and Messenger Ribonucleic Acid (mRNA) Detection
2.2.1. Cleaved Caspase-3 and Lysozyme Assay by Western Blot
2.2.2. mLyz1 and mDefa6 Assay by Quantitative RT-PCR
2.2.3. Small Intestinal Immunohistochemistry
2.2.4. Hematoxylin and Eosin (HE) Staining
2.2.5. Myeloperoxidase (MPO) Assay
2.2.6. Plasma Markers of Bacterial Translocation
2.3. The 16S rRNA Gene Amplicon Sequencing and Bioinformatics
2.4. Statistical Analysis Methods
3. Results
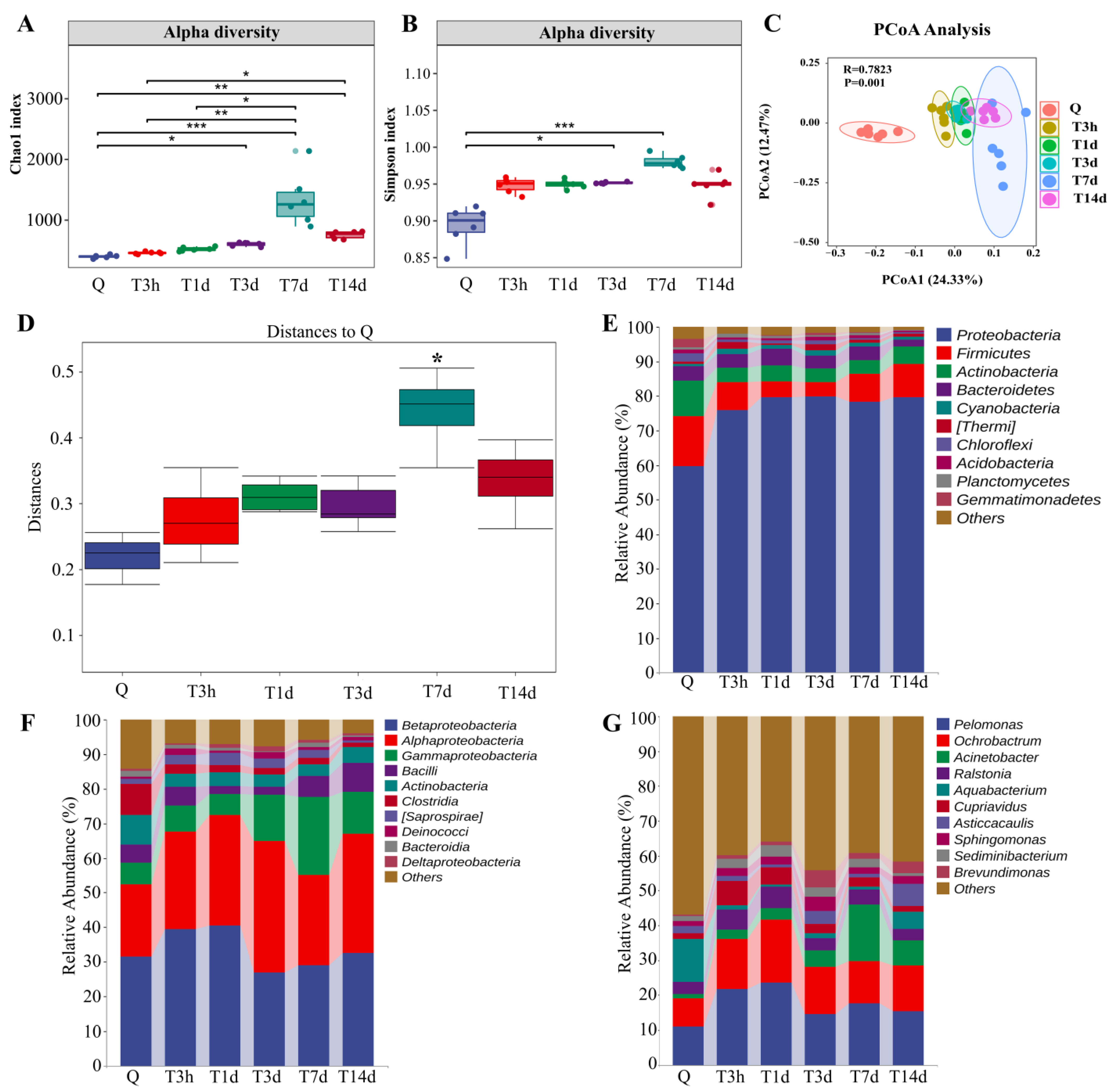
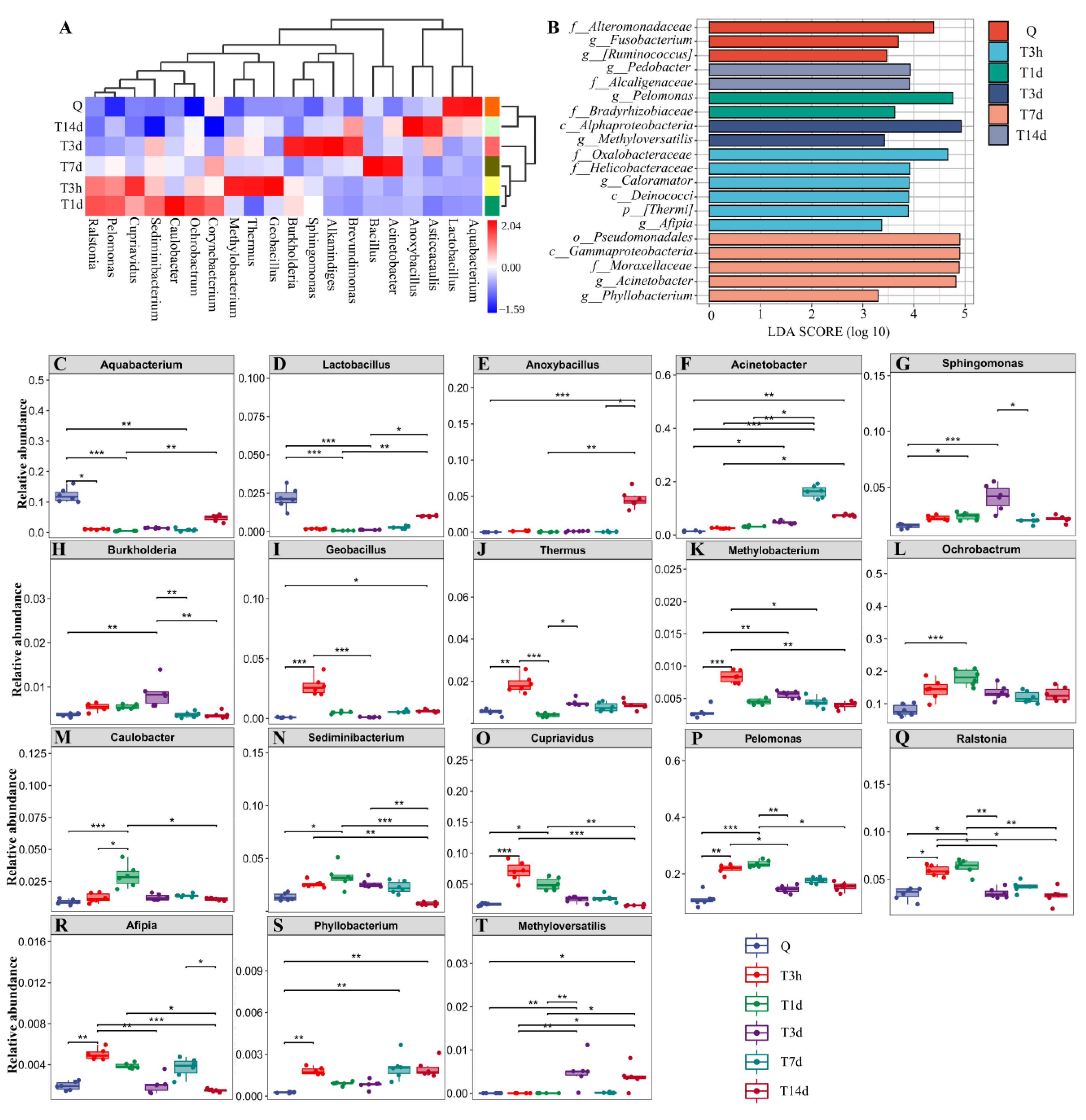
3.1. sTBI Alters the Lung Microbial Structure
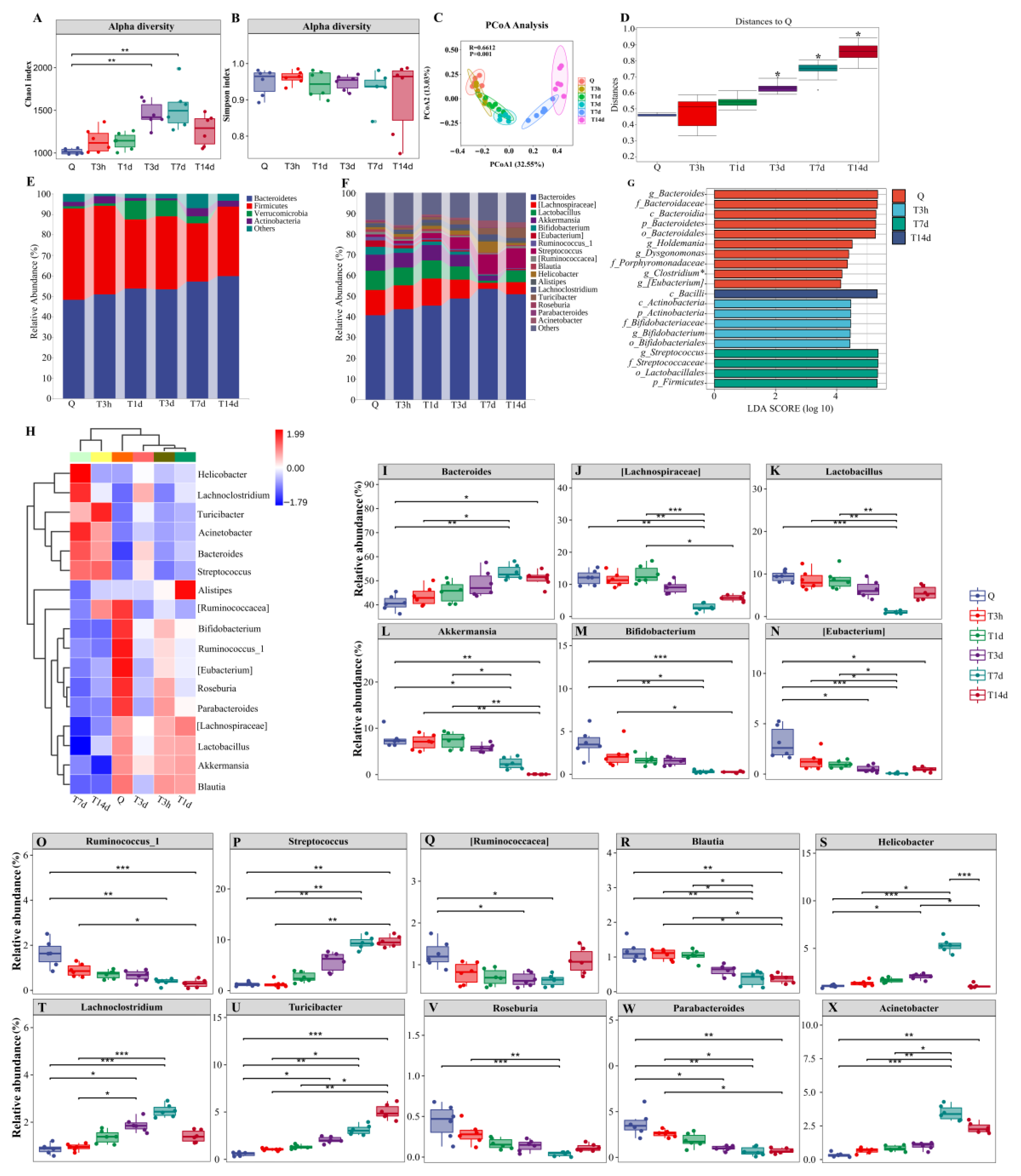
3.2. sTBI Induces Gut Microbiota Dysbiosis
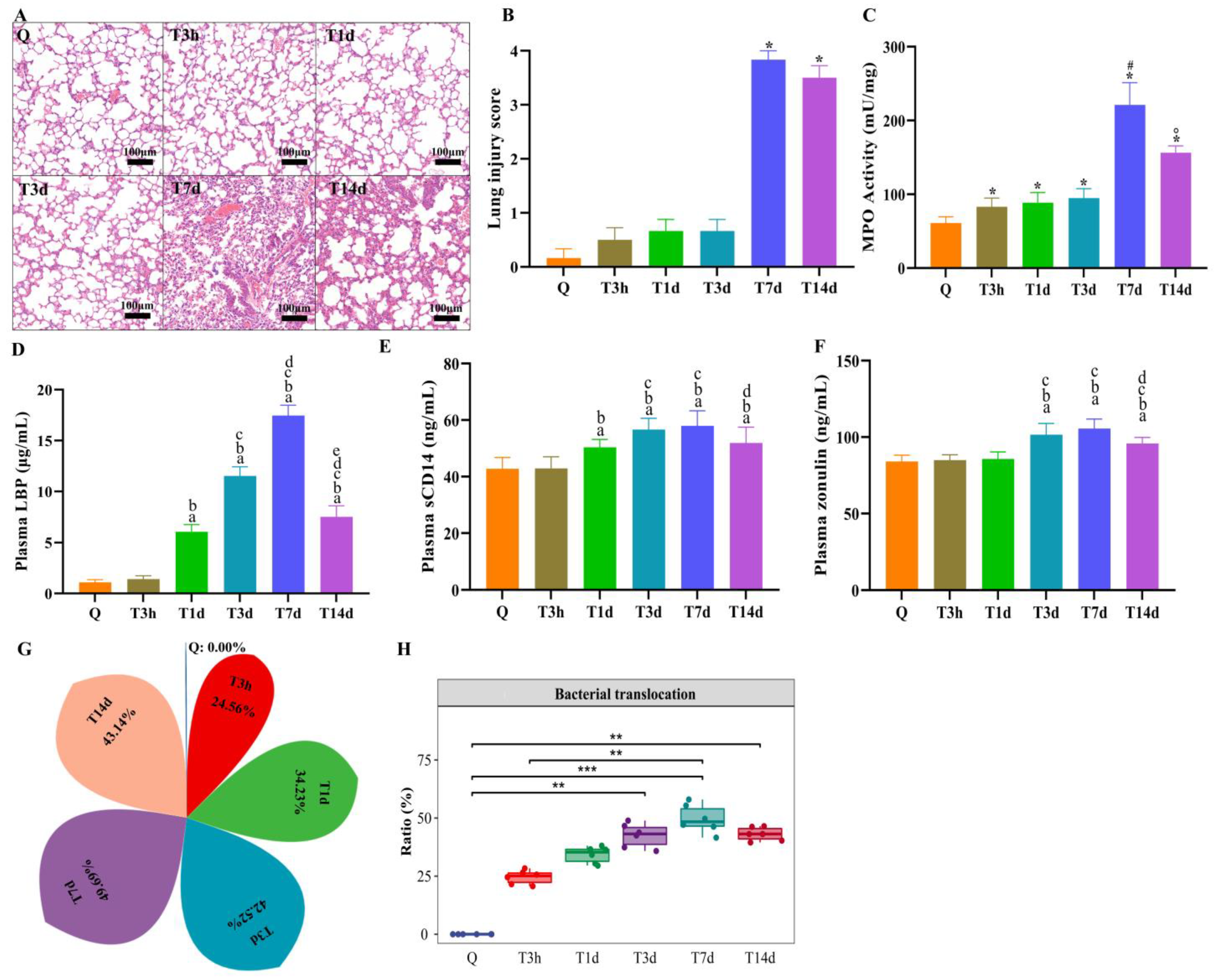
3.3. Lung Injury and Bacterial Translocation in Mice after sTBI
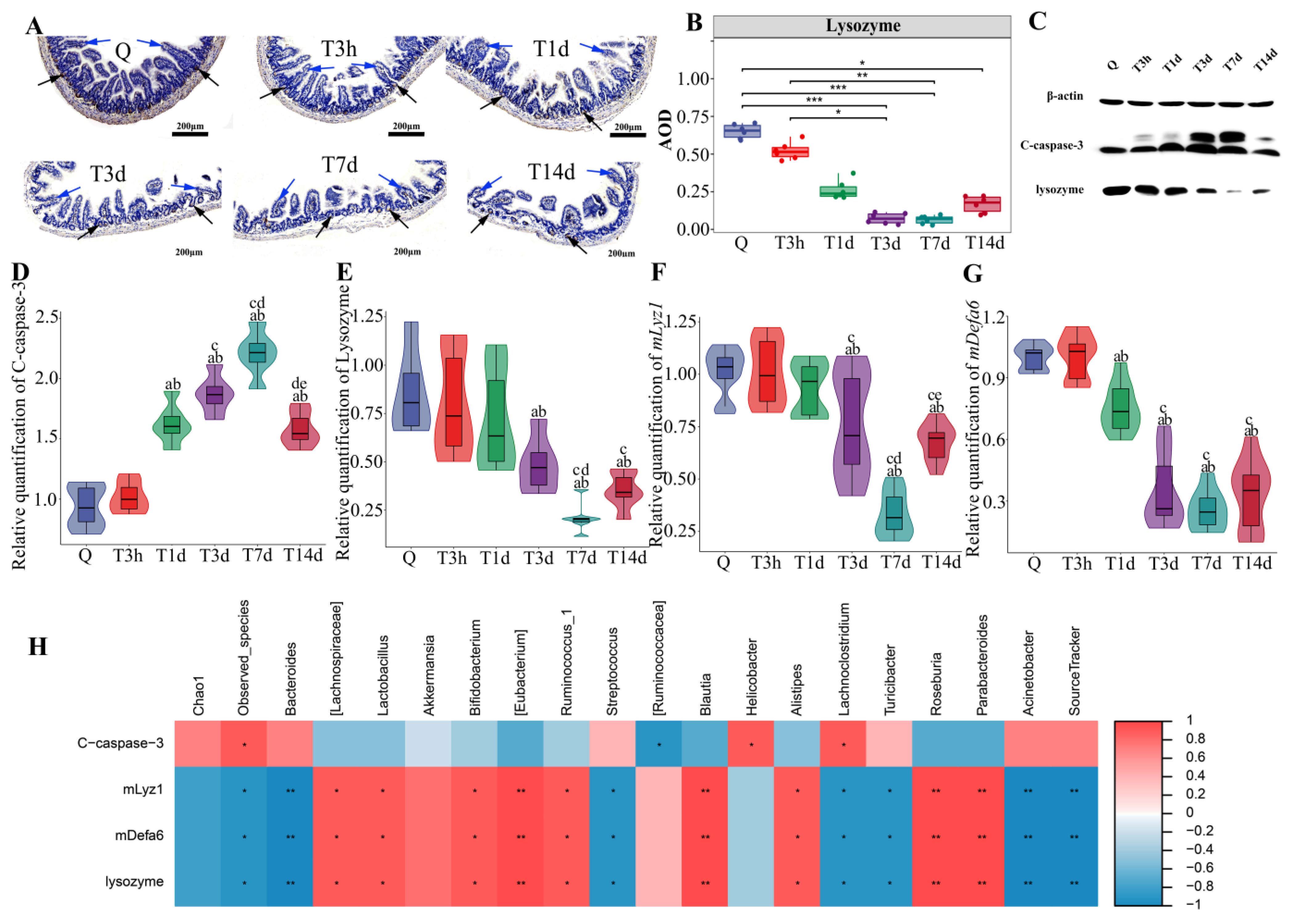
3.4. Paneth Cells Control of Gut Microbiota and Post-sTBI Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meyfroidt, G.; Bouzat, P.; Casaer, M.P.; Chesnut, R.; Hamada, S.R.; Helbok, R.; Hutchinson, P.; Maas, A.; Manley, G.; Menon, D.K.; et al. Management of moderate to severe traumatic brain injury: An update for the intensivist. Intensive Care Med. 2022, 48, 649–666. [Google Scholar] [CrossRef] [PubMed]
- Varela-Trinidad, G.U.; Dominguez-Diaz, C.; Solorzano-Castanedo, K.; Iniguez-Gutierrez, L.; Hernandez-Flores, T.J.; Fafutis-Morris, M. Probiotics: Protecting our health from the gut. Microorganisms 2022, 10, 1428. [Google Scholar] [CrossRef] [PubMed]
- Li, X.J.; You, X.Y.; Wang, C.Y.; Li, X.L.; Sheng, Y.Y.; Zhuang, P.W.; Zhang, Y.J. Bidirectional brain-gut-microbiota axis in increased intestinal permeability induced by central nervous system injury. CNS Neurosci. Ther. 2020, 26, 783–790. [Google Scholar] [CrossRef]
- Doran, S.J.; Henry, R.J.; Shirey, K.A.; Barrett, J.P.; Ritzel, R.M.; Lai, W.; Blanco, J.C.; Faden, A.I.; Vogel, S.N.; Loane, D.J. Early or late bacterial lung infection increases mortality after traumatic brain injury in male mice and chronically impairs monocyte innate immune function. Crit. Care. Med. 2020, 48, e418–e428. [Google Scholar] [CrossRef] [PubMed]
- Pittet, J.F.; Hu, P.J.; Honavar, J.; Brandon, A.P.; Evans, C.A.; Muthalaly, R.; Ding, Q.; Wagener, B.M. Estrogen alleviates sex-dependent differences in lung bacterial clearance and mortality secondary to bacterial pneumonia after traumatic brain injury. J. Neurotrauma 2021, 38, 989–999. [Google Scholar] [CrossRef] [PubMed]
- Panzer, A.R.; Lynch, S.V.; Langelier, C.; Christie, J.D.; McCauley, K.; Nelson, M.; Cheung, C.K.; Benowitz, N.L.; Cohen, M.J.; Calfee, C.S. Lung microbiota is related to smoking status and to development of acute respiratory distress syndrome in critically ill trauma patients. Am. J. Respir. Crit. Care. Med. 2018, 197, 621–631. [Google Scholar] [CrossRef]
- Dickson, R.P.; Singer, B.H.; Newstead, M.W.; Falkowski, N.R.; Erb-Downward, J.R.; Standiford, T.J.; Huffnagle, G.B. Enrichment of the lung microbiome with gut bacteria in sepsis and the acute respiratory distress syndrome. Nat. Microbiol. 2016, 1, 16113. [Google Scholar] [CrossRef] [Green Version]
- Stanley, D.; Mason, L.J.; Mackin, K.E.; Srikhanta, Y.N.; Lyras, D.; Prakash, M.D.; Nurgali, K.; Venegas, A.; Hill, M.D.; Moore, R.J.; et al. Translocation and dissemination of commensal bacteria in post-stroke infection. Nat. Med. 2016, 22, 1277–1284. [Google Scholar] [CrossRef]
- Wen, S.W.; Shim, R.; Ho, L.; Wanrooy, B.J.; Srikhanta, Y.N.; Prame, K.K.; Nicholls, A.J.; Shen, S.J.; Sepehrizadeh, T.; de-Veer, M.; et al. Advanced age promotes colonic dysfunction and gut-derived lung infection after stroke. Aging Cell 2019, 18, e12980. [Google Scholar] [CrossRef] [Green Version]
- Blanke, E.N.; Holmes, G.M.; Besecker, E.M. Altered physiology of gastrointestinal vagal afferents following neurotrauma. Neural. Regen. Res. 2021, 16, 254–263. [Google Scholar]
- Liu, Y.; Bao, Z.; Xu, X.; Chao, H.; Lin, C.; Li, Z.; Liu, Y.; Wang, X.; You, Y.; Liu, N.; et al. Extracellular signal-regulated kinase/nuclear factor-erythroid2-like2/heme oxygenase-1 pathway-mediated mitophagy alleviates traumatic brain injury-induced intestinal mucosa damage and epithelial barrier dysfunction. J. Neurotrauma 2017, 34, 2119–2131. [Google Scholar] [CrossRef] [PubMed]
- Chaaban, H.; Patel, M.M.; Burge, K.; Eckert, J.V.; Lupu, C.; Keshari, R.S.; Silasi, R.; Regmi, G.; Trammell, M.; Dyer, D.; et al. Early antibiotic exposure alters intestinal development and increases susceptibility to necrotizing enterocolitis: A mechanistic study. Microorganisms 2022, 10, 519. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.C.; Kern, J.T.; Jain, U.; Sonnek, N.M.; Xiong, S.; Simpson, K.F.; VanDussen, K.L.; Winkler, E.S.; Haritunians, T.; Malique, A.; et al. Western diet induces Paneth cell defects through microbiome alterations and farnesoid X receptor and type I interferon activation. Cell Host Microbe 2021, 29, 988–1001. [Google Scholar] [CrossRef]
- Nikolenko, V.N.; Oganesyan, M.V.; Sankova, M.V.; Bulygin, K.V.; Vovkogon, A.D.; Rizaeva, N.A.; Sinelnikov, M.Y. Paneth cells: Maintaining dynamic microbiome-host homeostasis, protecting against inflammation and cancer. Bioessays 2021, 43, e2000180. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Hanidziar, D. More of the Gut in the Lung: How two microbiomes meet in ARDS. Yale. J. Biol. Med. 2018, 91, 143–149. [Google Scholar] [PubMed]
- Kilkenny, C.; Browne, W.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Animal research: Reporting in vivo experiments: The ARRIVE guidelines. Br. J. Pharmacol. 2010, 160, 1577–1579. [Google Scholar] [CrossRef]
- Ma, X.; Aravind, A.; Pfister, B.J.; Chandra, N.; Haorah, J. Animal models of traumatic brain injury and assessment of injury severity. Mol. Neurobiol. 2019, 56, 5332–5345. [Google Scholar] [CrossRef]
- Daniel, S.; Phillippi, D.; Schneider, L.J.; Nguyen, K.N.; Mirpuri, J.; Lund, A.K. Exposure to diesel exhaust particles results in altered lung microbial profiles, associated with increased reactive oxygen species/reactive nitrogen species and inflammation, in C57Bl/6 wildtype mice on a high-fat diet. Part. Fibre. Toxicol. 2021, 18, 3. [Google Scholar] [CrossRef]
- Knights, D.; Kuczynski, J.; Charlson, E.S.; Zaneveld, J.; Mozer, M.C.; Collman, R.G.; Bushman, F.D.; Knight, R.; Kelley, S.T. Bayesian community-wide culture-independent microbial source tracking. Nat. Methods 2011, 8, 761–763. [Google Scholar] [CrossRef] [Green Version]
- Osadchiy, V.; Martin, C.R.; Mayer, E.A. The gut-brain axis and the microbiome: Mechanisms and clinical implications. Clin. Gastroenterol. Hepatol. 2019, 17, 322–332. [Google Scholar] [CrossRef]
- Siddiqui, R.; Akbar, N.; Ahmed, K.N. Gut microbiome and human health under the space environment. J. Appl. Microbiol. 2020, 130, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Tyler, P.T.; Grandhi, R. Gut microbiota and neurologic diseases and injuries. Adv. Exp. Med. Biol. 2020, 1238, 73–91. [Google Scholar]
- Weaver, J.L. The brain-gut axis: A prime therapeutic target in traumatic brain injury. Brain Res. 2021, 1753, 147225. [Google Scholar] [CrossRef] [PubMed]
- Busl, K.M. Nosocomial Infections in the neurointensive care unit. Neurol. Clin. 2017, 35, 785–807. [Google Scholar] [CrossRef] [PubMed]
- Jovanovic, B.; Milan, Z.; Markovic, D.L.; Djuric, O.; Radinovic, K.; Doklestic, K.; Velickovic, J.; Ivancevic, N.; Gregoric, P.; Pandurovic, M.; et al. Risk factors for ventilator-associated pneumonia in patients with severe traumatic brain injury in a Serbian trauma centre. Int. J. Infect. Dis. 2015, 38, 46–51. [Google Scholar] [CrossRef] [Green Version]
- Moffatt, M.F.; Cookson, W.O. The lung microbiome in health and disease. Clin. Med. 2017, 17, 525–529. [Google Scholar] [CrossRef]
- O’Dwyer, D.N.; Dickson, R.P.; Moore, B.B. The lung microbiome, immunity, and the pathogenesis of chronic lung disease. J. Immunol. 2016, 196, 4839–4847. [Google Scholar] [CrossRef] [Green Version]
- Lozupone, C.A.; Knight, R. Species divergence and the measurement of microbial diversity. Fems. Microbiol. Rev. 2008, 32, 557–578. [Google Scholar] [CrossRef]
- Dicker, A.J.; Huang, J.; Lonergan, M.; Keir, H.R.; Fong, C.J.; Tan, B.; Cassidy, A.J.; Finch, S.; Mullerova, H.; Miller, B.E.; et al. The sputum microbiome, airway inflammation, and mortality in chronic obstructive pulmonary disease. J. Allergy Clin. Immunol. 2021, 147, 158–167. [Google Scholar] [CrossRef]
- Oriano, M.; Gramegna, A.; Terranova, L.; Sotgiu, G.; Sulaiman, I.; Ruggiero, L.; Saderi, L.; Wu, B.; Chalmers, J.D.; Segal, L.N.; et al. Sputum neutrophil elastase associates with microbiota and Pseudomonas aeruginosa in bronchiectasis. Eur. Respir. J. 2020, 56, 2000769. [Google Scholar] [CrossRef]
- Zhang, R.; Chen, L.; Cao, L.; Li, K.J.; Huang, Y.; Luan, X.Q.; Li, G. Effects of smoking on the lower respiratory tract microbiome in mice. Respir. Res. 2018, 19, 253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greathouse, K.L.; White, J.R.; Vargas, A.J.; Bliskovsky, V.V.; Beck, J.A.; vo-Muhlinen, N.; Polley, E.C.; Bowman, E.D.; Khan, M.A.; Robles, A.I.; et al. Interaction between the microbiome and TP53 in human lung cancer. Genome. Biol. 2018, 19, 123. [Google Scholar] [CrossRef] [PubMed]
- Baek, M.G.; Woo, S.J.; Kim, N.E.; Baek, C.; Won, S.; Kim, Y.; Lee, J.J.; Yi, H.; Hong, J.Y. Respiratory microbiome profiles differ by recent hospitalization and nursing home residence in patients on mechanical ventilation. J. Transl. Med. 2020, 18, 464. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Vats, A.; Sharma, A.; Arora, A.; Kumar, A. The development of lower respiratory tract microbiome in mice. Microbiome 2017, 5, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daniel, S.; Pusadkar, V.; McDonald, J.; Mirpuri, J.; Azad, R.K.; Goven, A.; Lund, A.K. Traffic generated emissions alter the lung microbiota by promoting the expansion of Proteobacteria in C57Bl/6 mice placed on a high-fat diet. Ecotoxicol. Environ. Saf. 2021, 213, 112035. [Google Scholar] [CrossRef]
- Mortaz, E.; Adcock, I.M.; Folkerts, G.; Barnes, P.J.; Paul, V.A.; Garssen, J. Probiotics in the management of lung diseases. Mediators Inflamm. 2013, 2013, 751068. [Google Scholar] [CrossRef] [Green Version]
- Logan, N.A. Bacillus and relatives in foodborne illness. J. Appl. Microbiol. 2012, 112, 417–429. [Google Scholar] [CrossRef]
- Visca, P.; Seifert, H.; Towner, K.J. Acinetobacter infection—An emerging threat to human health. Iubmb. Life 2011, 63, 1048–1054. [Google Scholar] [CrossRef]
- Wong, D.; Nielsen, T.B.; Bonomo, R.A.; Pantapalangkoor, P.; Luna, B.; Spellberg, B. Clinical and pathophysiological overview of Acinetobacter infections: A century of challenges. Clin. Microbiol. Rev. 2017, 30, 409–447. [Google Scholar] [CrossRef] [Green Version]
- Nagpal, R.; Yadav, H. Bacterial translocation from the gut to the distant organs: An overview. Ann. Nutr. Metab. 2017, 71 (Suppl. S1), 11–16. [Google Scholar] [CrossRef]
- Stehle, J.J.; Leng, X.; Kitzman, D.W.; Nicklas, B.J.; Kritchevsky, S.B.; High, K.P. Lipopolysaccharide-binding protein, a surrogate marker of microbial translocation, is associated with physical function in healthy older adults. J. Gerontol. 2012, 67, 1212–1218. [Google Scholar] [CrossRef] [PubMed]
- Pieterman, E.J.; Princen, H.; Jarke, A.; Nilsson, R.; Cavallin, A.; Bergenholm, L.; Henricsson, M.; Gopaul, V.S.; Agrawal, R.; Nissen, S.E.; et al. Chronic oral administration of mineral oil compared with corn oil: Effects on gut permeability and plasma inflammatory and lipid biomarkers. Front. Pharmacol. 2021, 12, 681455. [Google Scholar] [CrossRef] [PubMed]
- Yeung, C.Y.; Chiang, C.J.; Cheng, M.L.; Chan, W.T.; Jiang, C.B.; Chang, S.W.; Liu, C.Y.; Chang, C.W.; Lee, H.C. Effects of vitamin D-deficient diet on intestinal epithelial integrity and zonulin expression in a C57bl/6 mouse model. Front. Med. 2021, 8, 649818. [Google Scholar] [CrossRef] [PubMed]
- Pan, P.; Song, Y.; Du, X.; Bai, L.; Hua, X.; Xiao, Y.; Yu, X. Intestinal barrier dysfunction following traumatic brain injury. Neurol. Sci. 2019, 40, 1105–1110. [Google Scholar] [CrossRef] [PubMed]
- Hanscom, M.; Loane, D.J.; Shea-Donohue, T. Brain-gut axis dysfunction in the pathogenesis of traumatic brain injury. J. Clin. Investig. 2021, 131, e143777. [Google Scholar] [CrossRef] [PubMed]
- Patterson, T.T.; Nicholson, S.; Wallace, D.; Hawryluk, G.; Grandhi, R. Complex feed-forward and feedback mechanisms underlie the relationship between traumatic brain injury and the gut-microbiota-brain axis. Shock 2019, 52, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Gao, Y.; Li, T.; Li, F.; Peng, R.; Wang, C.; Zhang, S.; Zhang, J.N. Recombinant human annexin A5 alleviated traumatic-brain-injury induced intestinal injury by regulating the Nrf2/HO-1/HMGB1 pathway. Molecules 2022, 27, 5755. [Google Scholar] [CrossRef]
- Ma, E.L.; Smith, A.D.; Desai, N.; Cheung, L.; Hanscom, M.; Stoica, B.A.; Loane, D.J.; Shea-Donohue, T.; Faden, A.I. Bidirectional brain-gut interactions and chronic pathological changes after traumatic brain injury in mice. Brain Behav. Immun. 2017, 66, 56–69. [Google Scholar] [CrossRef]
- Cray, P.; Sheahan, B.J.; Dekaney, C.M. Secretory sorcery: Paneth cell control of intestinal repair and homeostasis. Cell. Mol. Gastroenterol. Hepatol. 2021, 12, 1239–1250. [Google Scholar] [CrossRef] [PubMed]
- Delorme-Axford, E.; Klionsky, D.J. Secretory autophagy holds the key to lysozyme secretion during bacterial infection of the intestine. Autophagy 2018, 14, 365–367. [Google Scholar] [CrossRef]
- Barreto, E.B.L.; Rattes, I.C.; Da, C.A.; Gama, P. Paneth cells and their multiple functions. Cell Biol. Int. 2022, 46, 701–710. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, W.; Yuan, Q.; Li, Z.; Du, Z.; Wu, G.; Yu, J.; Hu, J. Translocation and Dissemination of Gut Bacteria after Severe Traumatic Brain Injury. Microorganisms 2022, 10, 2082. https://doi.org/10.3390/microorganisms10102082
Yang W, Yuan Q, Li Z, Du Z, Wu G, Yu J, Hu J. Translocation and Dissemination of Gut Bacteria after Severe Traumatic Brain Injury. Microorganisms. 2022; 10(10):2082. https://doi.org/10.3390/microorganisms10102082
Chicago/Turabian StyleYang, Weijian, Qiang Yuan, Zhiqi Li, Zhuoying Du, Gang Wu, Jian Yu, and Jin Hu. 2022. "Translocation and Dissemination of Gut Bacteria after Severe Traumatic Brain Injury" Microorganisms 10, no. 10: 2082. https://doi.org/10.3390/microorganisms10102082