
Initial pH Conditions Shape the Microbial Community Structure of Sewage Sludge in Batch Fermentations for the Improvement of Volatile Fatty Acid Production
 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Setup
2.2. sCOD and VFA Analysis
2.3. Prokaryotic Microbiota Structure Analysis
3. Results and Discussion
3.1. sCOD and pH Values in Batch Fermentations
3.2. Sequencing Output and Analysis
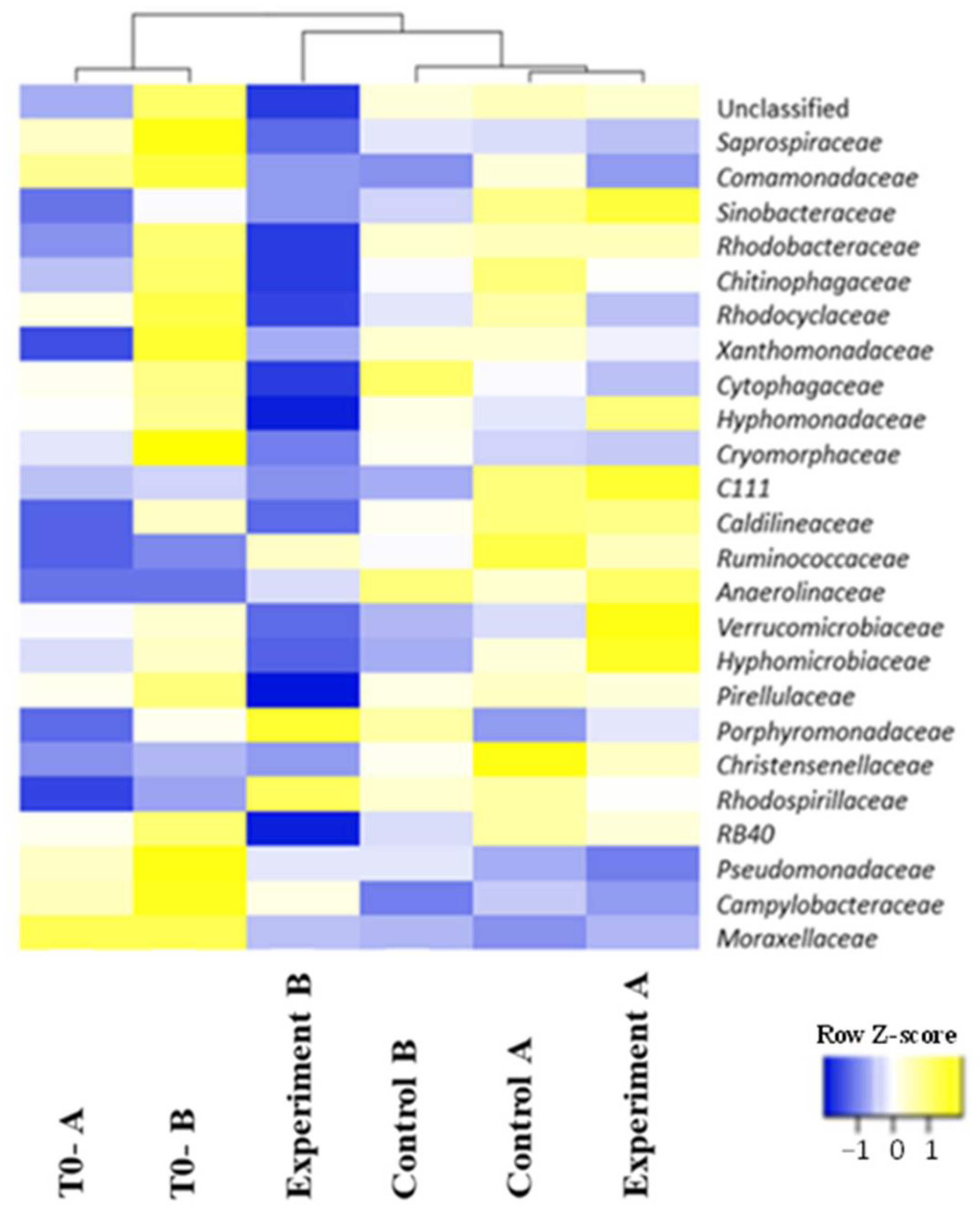
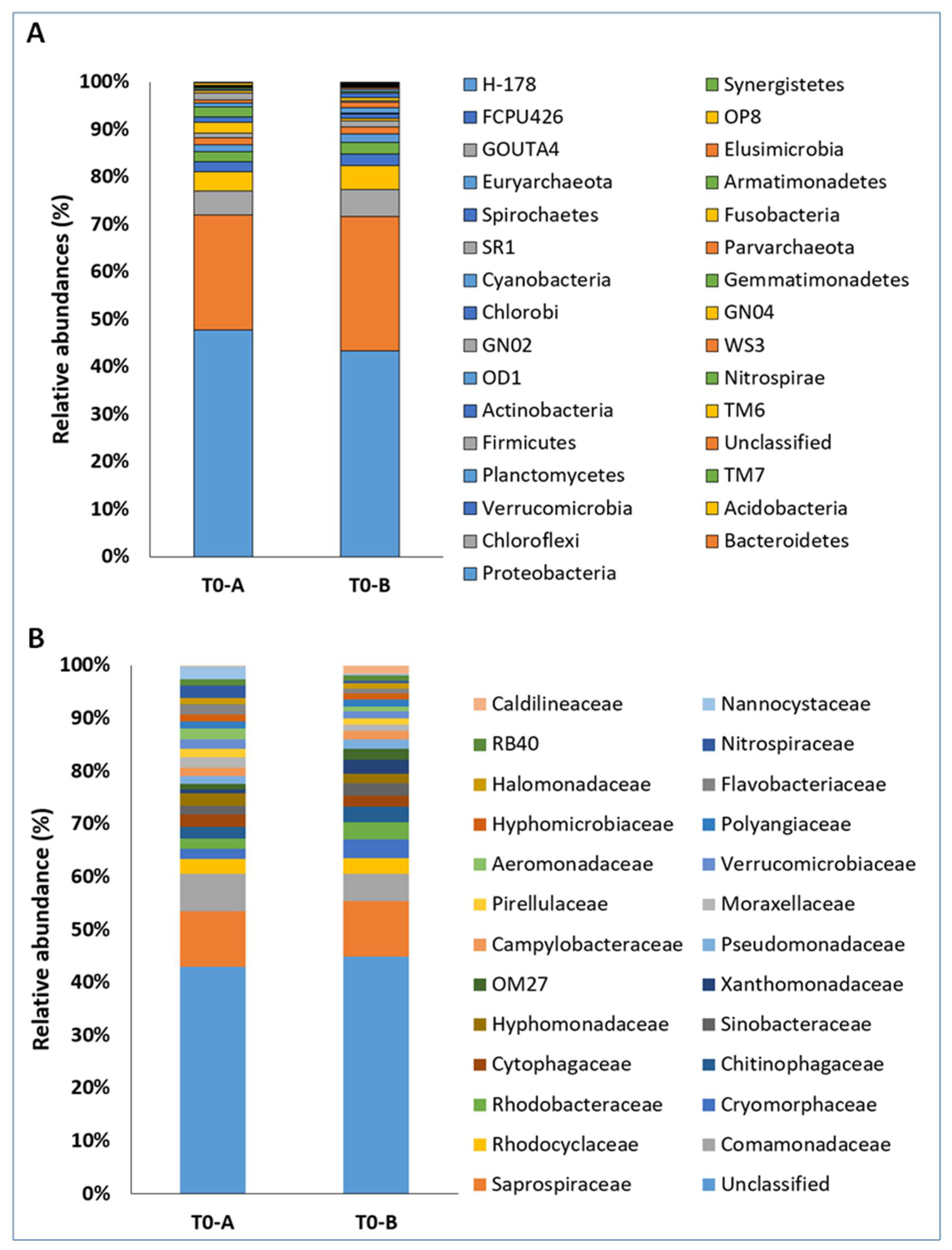
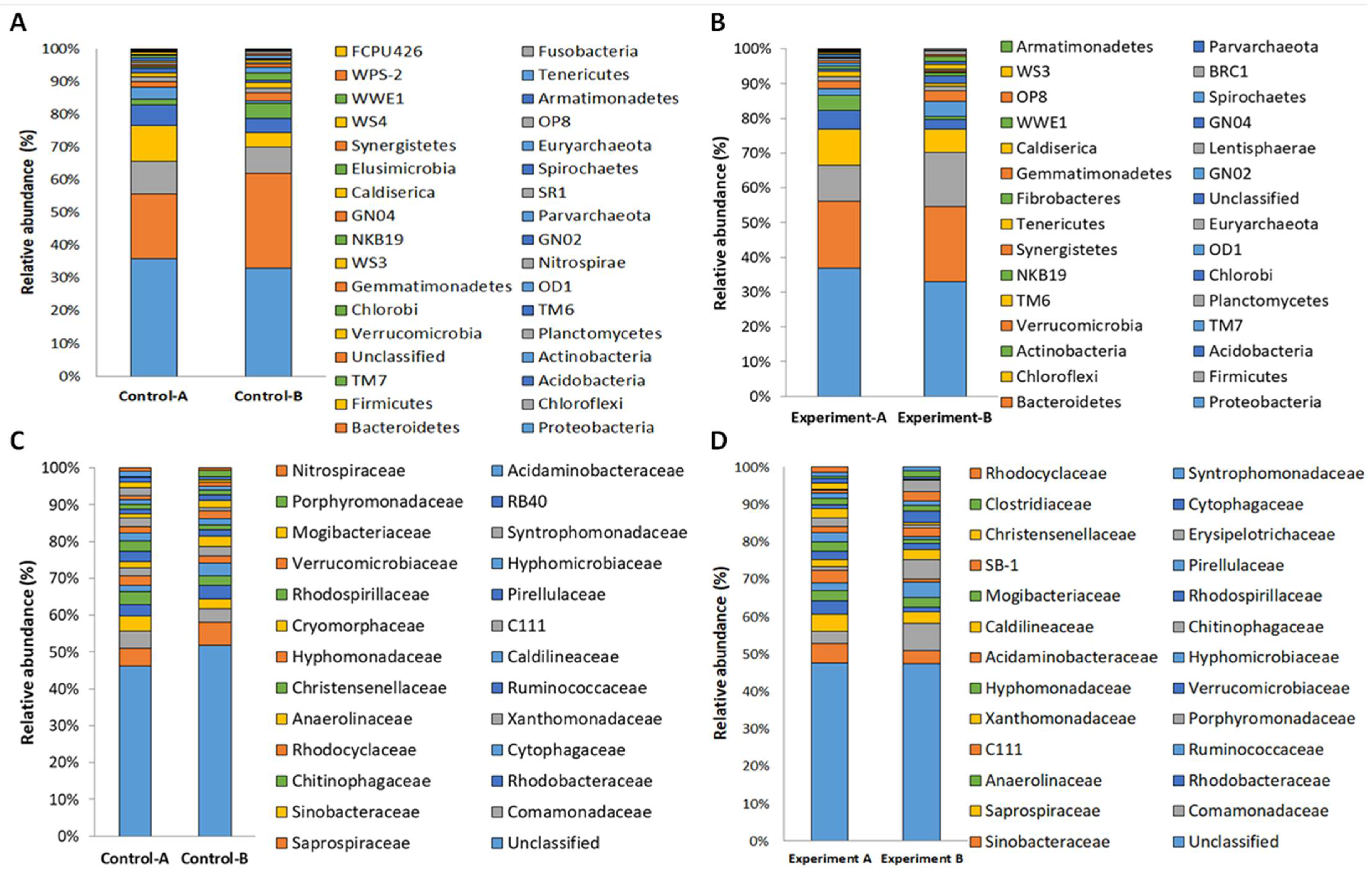
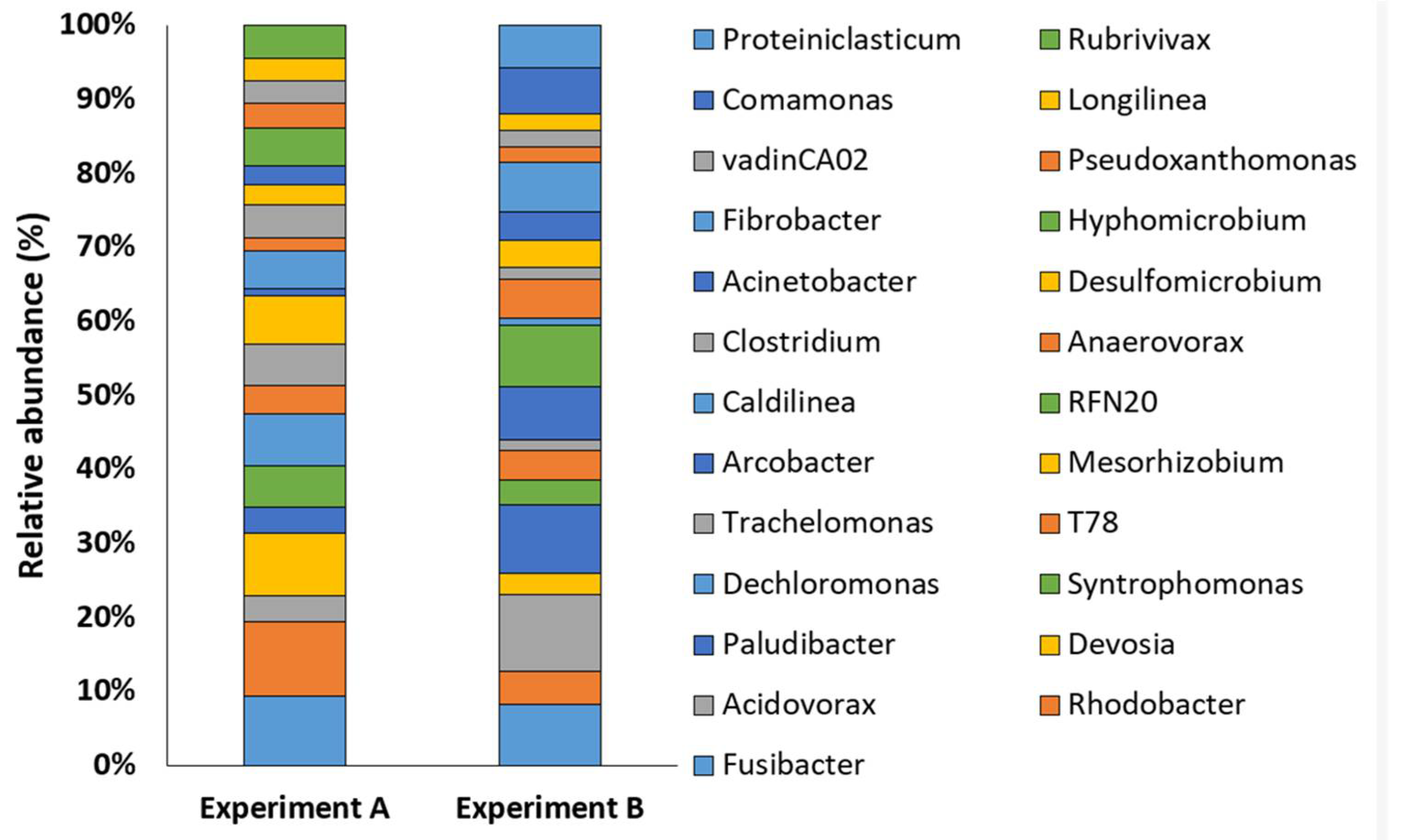
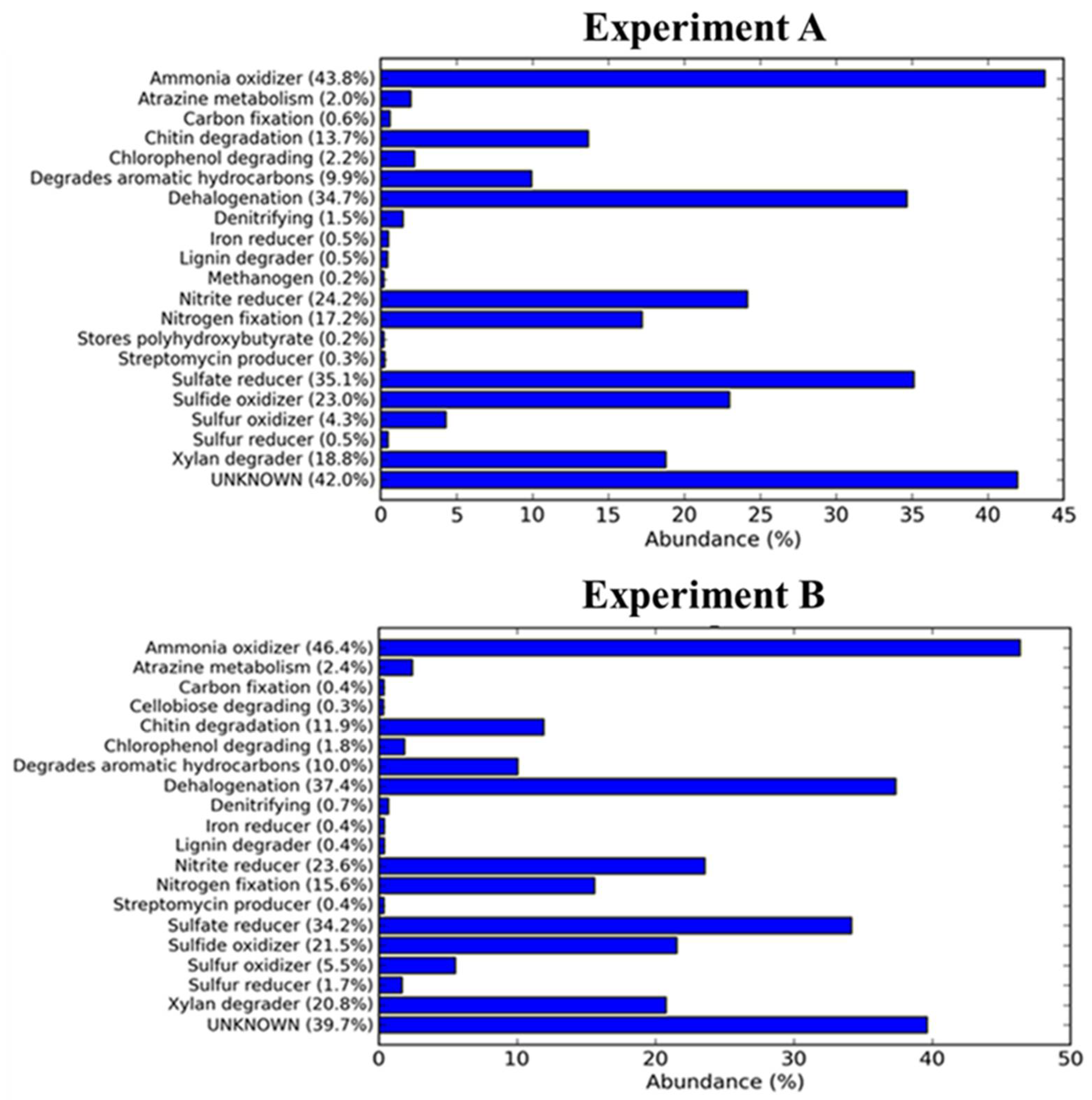
3.3. Structure of Prokaryotic Communities of Sewage Sludge
3.4. Metabolism and Oxygen Requirement Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Choudri, B.S.; Al-Awadhi, T.; Charabi, Y.; Al-Nasiri, N. Wastewater Treatment, Reuse, and Disposal-Associated Effects on Environment and Health. Water Environ. Res. 2020, 92, 1595–1602. [Google Scholar] [CrossRef] [PubMed]
- Newhart, K.B.; Holloway, R.W.; Hering, A.S.; Cath, T.Y. Data-Driven Performance Analyses of Wastewater Treatment Plants: A Review. Water Res. 2019, 157, 498–513. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.; Wang, X.; Liu, R.; Li, S.; van Loosdrecht, M.C.M.; Jiang, H. Environmental Impacts of Resource Recovery from Wastewater Treatment Plants. Water Res. 2019, 160, 268–277. [Google Scholar] [CrossRef]
- Mininni, G.; Mauro, E.; Piccioli, B.; Colarullo, G.; Brandolini, F.; Giacomelli, P. Production and Characteristics of Sewage Sludge in Italy. Water Sci. Technol. 2019, 79, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Mannina, G.; Badalucco, L.; Barbara, L.; Cosenza, A.; Di Trapani, D.; Gallo, G.; Laudicina, V.A.; Marino, G.; Muscarella, S.M.; Presti, D.; et al. Enhancing a Transition to a Circular Economy in the Water Sector: The EU Project WIDER UPTAKE. Water 2021, 13, 946. [Google Scholar] [CrossRef]
- Hejase, H.J.; Hejase, A.J.; Tabsh, H.; Chalak, H.C.; Wamitu, S.N.; Pavel, S. Advances in Business and Management; Scientific Research Publishing, Inc.: Wuhan, China, 2018. [Google Scholar]
- Tarrahi, R.; Fathi, Z.; Seydibeyoğlu, M.Ö.; Doustkhah, E.; Khataee, A. Polyhydroxyalkanoates (PHA): From Production to Nanoarchitecture. Int. J. Biol. Macromol. 2020, 146, 596–619. [Google Scholar] [CrossRef]
- Sarkar, O.; Butti, S.K.; Mohan, S.V. Acidogenic Biorefinery: Food Waste Valorization to Biogas and Platform Chemicals. In Waste Biorefinery; Elsevier: Amsterdam, The Netherlands, 2018; pp. 203–218. [Google Scholar]
- Sarkar, O.; Rova, U.; Christakopoulos, P.; Matsakas, L. Influence of Initial Uncontrolled PH on Acidogenic Fermentation of Brewery Spent Grains to Biohydrogen and Volatile Fatty Acids Production: Optimization and Scale-Up. Bioresour. Technol. 2021, 319, 124233. [Google Scholar] [CrossRef]
- Shahid, S.; Razzaq, S.; Farooq, R. Polyhydroxyalkanoates: Next Generation Natural Biomolecules and a Solution for the World’s Future Economy. Int. J. Biol. Macromol. 2021, 166, 297–321. [Google Scholar] [CrossRef]
- Zheng, Y.; Guo, L.; Liu, Y.; She, Z.; Gao, M.; Jin, C.; Zhao, Y. Effects of Chemical Oxygen Demand Concentration, PH and Operation Cycle on Polyhydroxyalkanoates Synthesis with Waste Sludge. Environ. Technol. 2021, 42, 1922–1929. [Google Scholar] [CrossRef]
- Zhang, L.; Loh, K.-C.; Dai, Y.; Tong, Y.W. Acidogenic Fermentation of Food Waste for Production of Volatile Fatty Acids: Bacterial Community Analysis and Semi-Continuous Operation. Waste Manag. 2020, 109, 75–84. [Google Scholar] [CrossRef]
- Chen, Z.; Rao, Y.; Usman, M.; Chen, H.; Białowiec, A.; Zhang, S.; Luo, G. Anaerobic Fermentation of Hydrothermal Liquefaction Wastewater of Dewatered Sewage Sludge for Volatile Fatty Acids Production with Focuses on the Degradation of Organic Components and Microbial Community Compositions. Sci. Total Environ. 2021, 777, 146077. [Google Scholar] [CrossRef] [PubMed]
- Miguel, M.A.; Lee, S.S.; Mamuad, L.L.; Choi, Y.J.; Jeong, C.D.; Son, A.; Cho, K.K.; Kim, E.T.; Kim, S.B.; Lee, S.S. Enhancing Butyrate Production, Ruminal Fermentation and Microbial Population through Supplementation with Clostridium Saccharobutylicum. J. Microbiol Biotechnol. 2019, 29, 1083–1095. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Tsui, T.-H.; Loh, K.-C.; Dai, Y.; Tong, Y.W. Effects of Plastics on Reactor Performance and Microbial Communities during Acidogenic Fermentation of Food Waste for Production of Volatile Fatty Acids. Bioresour. Technol. 2021, 337, 125481. [Google Scholar] [CrossRef] [PubMed]
- Fra-Vázquez, A.; Pedrouso, A.; Del Rio, A.V.; Mosquera-Corral, A. Volatile Fatty Acid Production from Saline Cooked Mussel Processing Wastewater at Low PH. Sci. Total Environ. 2020, 732, 139337. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Aguirre, J.; Aymerich, E.; de Goñi, J.G.-M.; Esteban-Gutiérrez, M. Selective VFA Production Potential from Organic Waste Streams: Assessing Temperature and PH Influence. Bioresour. Technol. 2017, 244, 1081–1088. [Google Scholar] [CrossRef] [PubMed]
- Slezak, R.; Grzelak, J.; Krzystek, L.; Ledakowicz, S. Influence of Initial PH on the Production of Volatile Fatty Acids and Hydrogen during Dark Fermentation of Kitchen Waste. Environ. Technol. 2021, 42, 4269–4278. [Google Scholar] [CrossRef]
- Presti, D.; Cosenza, A.; Capri, F.C.; Gallo, G.; Alduina, R.; Mannina, G. Influence of Volatile Solids and PH for the Production of Volatile Fatty Acids: Batch Fermentation Tests Using Sewage Sludge. Bioresour. Technol. 2021, 342, 125853. [Google Scholar] [CrossRef]
- Muhorakeye, A.; Cayetano, R.D.; Kumar, A.N.; Park, J.; Pandey, A.K.; Kim, S.-H. Valorization of Pretreated Waste Activated Sludge to Organic Acids and Biopolymer. Chemosphere 2022, 303, 135078. [Google Scholar] [CrossRef]
- Mannina, G.; Badalucco, L.; Barbara, L.; Cosenza, A.; Di Trapani, D.; Laudicina, V.A.; Muscarella, S.M.; Presti, D. Roadmapping the Transition to Water Resource Recovery Facilities: The Two Demonstration Case Studies of Corleone and Marineo (Italy). Water 2022, 14, 156. [Google Scholar] [CrossRef]
- Arndt, D.; Xia, J.; Liu, Y.; Zhou, Y.; Guo, A.C.; Cruz, J.A.; Sinelnikov, I.; Budwill, K.; Nesbø, C.L.; Wishart, D.S. METAGENassist: A Comprehensive Web Server for Comparative Metagenomics. Nucleic Acids Res. 2012, 40, W88–W95. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, C.; Zhang, X.; Feng, L.; Li, Y.; Zhou, Q. Waste Activated Sludge Hydrolysis and Acidification: A Comparison between Sodium Hydroxide and Steel Slag Addition. J. Environ. Sci. 2016, 48, 200–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atasoy, M.; Eyice, Ö.; Cetecioglu, Z. Volatile Fatty Acid Production from Semi-Synthetic Milk Processing Wastewater under Alkali PH: The Pearls and Pitfalls of Microbial Culture. Bioresour. Technol. 2020, 297, 122415. [Google Scholar] [CrossRef]
- Wang, Z.; Yan, M.; Chen, X.; Li, D.; Qin, L.; Li, Z.; Yao, J.; Liang, X. Mixed Culture of Saccharomyces Cerevisiae and Acetobacter Pasteurianus for Acetic Acid Production. Biochem. Eng. J. 2013, 79, 41–45. [Google Scholar] [CrossRef]
- Guo, J.; Ni, B.-J.; Han, X.; Chen, X.; Bond, P.; Peng, Y.; Yuan, Z. Data on Metagenomic Profiles of Activated Sludge from a Full-Scale Wastewater Treatment Plant. Data Br. 2017, 15, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Chen, Y.; Feng, L. Polycyclic Aromatic Hydrocarbon Affects Acetic Acid Production during Anaerobic Fermentation of Waste Activated Sludge by Altering Activity and Viability of Acetogen. Environ. Sci. Technol. 2016, 50, 6921–6929. [Google Scholar] [CrossRef]
- Zheng, X.; Su, Y.; Li, X.; Xiao, N.; Wang, D.; Chen, Y. Pyrosequencing Reveals the Key Microorganisms Involved in Sludge Alkaline Fermentation for Efficient Short-Chain Fatty Acids Production. Environ. Sci. Technol. 2013, 47, 4262–4268. [Google Scholar] [CrossRef] [PubMed]
- Riviere, D.; Desvignes, V.; Pelletier, E.; Chaussonnerie, S.; Guermazi, S.; Weissenbach, J.; Li, T.; Camacho, P.; Sghir, A. Towards the Definition of a Core of Microorganisms Involved in Anaerobic Digestion of Sludge. ISME J. 2009, 3, 700–714. [Google Scholar] [CrossRef] [Green Version]
- Vu, D.H.; Wainaina, S.; Taherzadeh, M.J.; Åkesson, D.; Ferreira, J.A. Production of Polyhydroxyalkanoates (PHAs) by Bacillus Megaterium Using Food Waste Acidogenic Fermentation-Derived Volatile Fatty Acids. Bioengineered 2021, 12, 2480–2498. [Google Scholar] [CrossRef]
- Johnston, J.; LaPara, T.; Behrens, S. Composition and Dynamics of the Activated Sludge Microbiome during Seasonal Nitrification Failure. Sci. Rep. 2019, 9, 4565. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Chu, L. Biological Nitrate Removal from Water and Wastewater by Solid-Phase Denitrification Process. Biotechnol. Adv. 2016, 34, 1103–1112. [Google Scholar] [CrossRef]
- Yang, X.; Wen, L.; Liu, X.; Chen, S.; Wang, Y.; Wan, C. Bio-Augmentative Volatile Fatty Acid Production from Waste Activated Sludge Hydrolyzed at PH 12. RSC Adv. 2015, 5, 50033–50039. [Google Scholar] [CrossRef]
- Zakaria, M.R.; Tabatabaei, M.; Ghazali, F.M.; Abd-Aziz, S.; Shirai, Y.; Hassan, M.A. Polyhydroxyalkanoate Production from Anaerobically Treated Palm Oil Mill Effluent by New Bacterial Strain Comamonas Sp. EB172. World J. Microbiol. Biotechnol. 2010, 26, 767–774. [Google Scholar] [CrossRef]
- Pereira, J.; Queirós, D.; Lemos, P.C.; Rossetti, S.; Serafim, L.S. Enrichment of a Mixed Microbial Culture of PHA-Storing Microorganisms by Using Fermented Hardwood Spent Sulfite Liquor. New Biotechnol. 2020, 56, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Ueki, A.; Akasaka, H.; Suzuki, D.; Ueki, K. Paludibacter Propionicigenes Gen. Nov., Sp. Nov., a Novel Strictly Anaerobic, Gram-Negative, Propionate-Producing Bacterium Isolated from Plant Residue in Irrigated Rice-Field Soil in Japan. Int. J. Syst. Evol. Microbiol. 2006, 56, 39–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jie, W.; Peng, Y.; Ren, N.; Li, B. Volatile Fatty Acids (VFAs) Accumulation and Microbial Community Structure of Excess Sludge (ES) at Different pHs. Bioresour. Technol. 2014, 152, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, R.; Toyama, K.; Miyanaga, K.; Tanji, Y. Identification of Crude-Oil Components and Microorganisms That Cause Souring under Anaerobic Conditions. Appl. Microbiol. Biotechnol. 2014, 98, 1853–1861. [Google Scholar] [CrossRef] [PubMed]
- Xin, X.; She, Y.; Hong, J. Insights into Microbial Interaction Profiles Contributing to Volatile Fatty Acids Production via Acidogenic Fermentation of Waste Activated Sludge Assisted by Calcium Oxide Pretreatment. Bioresour. Technol. 2021, 320, 124287. [Google Scholar] [CrossRef] [PubMed]
- Ramin, M.; Alimon, A.R.; Panandam, J.M.; Sijam, K.; Javanmard, A.; Abdullah, N. Digestion of Rice Straw and Oil Palm Fronds by Microflora from Rumen and Termite Bacteria, in Vitro. Pak. J. Biol. Sci. 2008, 11, 583–588. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Yang, H.; Wang, X.; Cao, W.; Guo, L. Strong PH Dependence of Hydrogen Production from Glucose by Rhodobacter Sphaeroides. Int. J. Hydrogen Energy 2020, 45, 9451–9458. [Google Scholar] [CrossRef]
- Goud, R.K.; Arunasri, K.; Yeruva, D.K.; Krishna, K.V.; Dahiya, S.; Mohan, S.V. Impact of Selectively Enriched Microbial Communities on Long-Term Fermentative Biohydrogen Production. Bioresour. Technol. 2017, 242, 253–264. [Google Scholar] [CrossRef]
- Wang, Q.; Liang, Y.; Zhao, P.; Li, Q.X.; Guo, S.; Chen, C. Potential and Optimization of Two-Phase Anaerobic Digestion of Oil Refinery Waste Activated Sludge and Microbial Community Study. Sci. Rep. 2016, 6, 38245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh Saharan, B.; Grewal, A.; Kumar, P. Biotechnological Production of Polyhydroxyalkanoates: A Review on Trends and Latest Developments. Chin. J. Biol. 2014, 2014, 802984. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Lee, C. Response of a Continuous Anaerobic Digester to Temperature Transitions: A Critical Range for Restructuring the Microbial Community Structure and Function. Water Res. 2016, 89, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Luo, K.; Pang, Y.; Yang, Q.; Wang, D.; Li, X.; Lei, M.; Huang, Q. A Critical Review of Volatile Fatty Acids Produced from Waste Activated Sludge: Enhanced Strategies and Its Applications. Environ. Sci. Pollut. Res. 2019, 26, 13984–13998. [Google Scholar] [CrossRef]
- Schellenberger, S.; Drake, H.L.; Kolb, S. Functionally Redundant Cellobiose-Degrading Soil Bacteria Respond Differentially to Oxygen. Appl. Environ. Microbiol. 2011, 77, 6043–6048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiken, C.J.; Breuer, G.; Klaversma, E.; Santiago, T.; Van Loosdrecht, M.C.M. Sieving Wastewater–Cellulose Recovery, Economic and Energy Evaluation. Water Res. 2013, 47, 43–48. [Google Scholar] [CrossRef]
- Odriozola, M.; Abraham, E.; Lousada-Ferreira, M.; Spanjers, H.; Van Lier, J.B. Identification of the Methanogenesis Inhibition Mechanism Using Comparative Analysis of Mathematical Models. Front. Bioeng. Biotechnol. 2019, 7, 93. [Google Scholar] [CrossRef] [Green Version]
- Samanides, C.G.; Koutsokeras, L.; Constantinides, G.; Vyrides, I. Methanogenesis Inhibition in Anaerobic Granular Sludge for the Generation of Volatile Fatty Acids from CO2 and Zero Valent Iron. Front. Energy Res. 2020, 8, 37. [Google Scholar] [CrossRef]
- Khatami, K.; Atasoy, M.; Ludtke, M.; Baresel, C.; Eyice, Ö.; Cetecioglu, Z. Bioconversion of Food Waste to Volatile Fatty Acids: Impact of Microbial Community, PH and Retention Time. Chemosphere 2021, 275, 129981. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | Sample | Sampling Date | Sample Details |
|---|---|---|---|
| Condition A (pH 8) | T0-A | 01/06/2021 | Sewage sludge sample collected from Marineo WWTP and used as inoculum for Control A and Experiment A batch fermentations |
| Control A | 16/06/2021 | Sewage sludge sample collected after batch fermentation without pH adjustment | |
| Experiment A | 16/06/2021 | Sewage sludge sample collected after batch fermentation with initial pH adjustment at 8 | |
| Condition B (pH 10) | T0-B | 25/06/2021 | Sewage sludge sample collected from Marineo WWTP and used as inoculum for control B and experiment B batch fermentations |
| Control B | 9/07/2021 | Sewage sludge sample collected after fermentation without pH adjustment | |
| Experiment B | 9/07/2021 | Sewage sludge sample collected after fermentation with initial pH adjustment at 10 |
| Batch Fermentation Condition A—Initial pH 8 | |||||
|---|---|---|---|---|---|
| Control A | Experiment A | ||||
| Day | sCOD mg/L | pH | Day | sCOD mg/L | pH |
| 0 | 790 ± 16 | 7.2 ± 0.3 | 0 | 790 ± 16 | 8 ± 0.8 |
| 1 | 930 ± 18 | 6.8 ± 0.2 | 1 | 1186 ± 8 | 6.8 ± 0.5 |
| 2 | 1329 ± 25 | 6.6 ± 0.1 | 2 | 1377 ± 10 | 6.7 ± 0.3 |
| 3 | 1374 ± 26 | 6.6 ± 0.1 | 3 | 1587 ± 15 | 6.6 ± 0.2 |
| 4 | 1468.5 ± 25 | 6.6 ± 0.1 | 4 | 1705.5 ± 15 | 6.6 ± 0.1 |
| 5 | 1563 ± 34 | 6.6 ± 0.1 | 5 | 1824 ± 22 | 6.6 ± 0.1 |
| 8 | 1359 ± 14 | 6.6 ± 0.1 | 8 | 1540.5 ± 27 | 6.8 ± 0.1 |
| 9 | 1155 ± 17 | 6.6 ± 0.1 | 9 | 1257 ± 25 | 6.8 ± 0.1 |
| 10 | 1117.5 ± 19 | 6.7 ± 0.1 | 10 | 1188 ± 23 | 6.7 ± 0.1 |
| 11 | 1080 ± 20 | 6.7 ± 0.1 | 11 | 1119 ± 18 | 6.7 ± 0.1 |
| 12 | 1035 ± 18 | 6.7 ± 0.1 | 12 | 1089 ± 20 | 6.8 ± 0.1 |
| 15 | 825 ± 15 | 6.7 ± 0.1 | 15 | 813 ± 15 | 6.8 ± 0.1 |
| Batch Fermentation Condition B—Starting pH 10 | |||||
|---|---|---|---|---|---|
| Control B | Experiment B | ||||
| Day | sCOD mg/L | pH | Day | sCOD mg/L | pH |
| 0 | 237 ± 6 | 7.2 ± 0.3 | 0 | 237 ± 6 | 10 ± 2 |
| 1 | 556 ± 15 | 6.8 ± 0.2 | 1 | 2234 ± 31 | 7.4 ± 1 |
| 2 | 809 ± 14 | 6.6 ± 0.1 | 2 | 2413 ± 28 | 6.7 ± 0.4 |
| 3 | 1428 ± 20 | 6.6 ± 0.1 | 3 | 2568 ± 24 | 6.6 ± 0.1 |
| 6 | 1756 ± 26 | 6.6 ± 0.1 | 6 | 2733 ± 36 | 6.7 ± 0.1 |
| 7 | 1661 ± 30 | 6.6 ± 0.1 | 7 | 2916 ± 34 | 6.8 ± 0.1 |
| 8 | 1444 ± 24 | 6.6 ± 0.1 | 8 | 2838 ± 32 | 6.8 ± 0.1 |
| 9 | 1180 ± 19 | 6.6 ± 0.1 | 9 | 2610 ± 29 | 6.8 ± 0.1 |
| 10 | 1131 ± 17 | 6.7 ± 0.1 | 10 | 2418 ± 27 | 6.8 ± 0.1 |
| 13 | 1023 ± 21 | 6.7 ± 0.1 | 13 | 2070 ± 34 | 6.8 ± 0.1 |
| 14 | 1014 ± 23 | 6.7 ± 0.1 | 14 | 2025 ± 36 | 6.8 ± 0.1 |
| Batch Fermentation | Peak Day of sCOD | VFA mg COD/L | VFA/sCOD |
|---|---|---|---|
| Control A | 5 | 1303.54 ± 54.07 | 0.834 |
| Experiment A | 5 | 1391.71 ± 27.66 | 0.763 |
| Control B | 6 | 1411.82 ± 28.02 | 0.8 |
| Experiment B | 7 | 2020 ± 37.04 | 0.69 |
| Sample | Input | Filtered | Percentage of Input Passed Filter (%) | Denoised | Merged | Percentage of Input Merged (%) | Nonchimeric | Percentage of Input Nonchimeric (%) |
|---|---|---|---|---|---|---|---|---|
| T0-A | 74,926 | 61,752 | 82.42 | 56,341 | 45,171 | 60.29 | 28,642 | 38.23 |
| Control A | 107,349 | 88,438 | 82.38 | 82,197 | 66,368 | 61.82 | 40,084 | 37.34 |
| Experiment A | 101,116 | 81,999 | 81.09 | 76,364 | 63,186 | 62.49 | 37,502 | 37.09 |
| T0-B | 117,728 | 96,462 | 81.94 | 89,387 | 75,002 | 63.71 | 46,400 | 39.41 |
| Control B | 83,492 | 67,136 | 80.41 | 61,455 | 51,157 | 61.27 | 32,491 | 38.92 |
| Experiment B | 61,013 | 51,049 | 83.67 | 46,841 | 40,321 | 66.09 | 18,048 | 29.58 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Leto, Y.; Capri, F.C.; Mineo, A.; Cosenza, A.; Gallo, G.; Alduina, R.; Mannina, G. Initial pH Conditions Shape the Microbial Community Structure of Sewage Sludge in Batch Fermentations for the Improvement of Volatile Fatty Acid Production. Microorganisms 2022, 10, 2073. https://doi.org/10.3390/microorganisms10102073
Di Leto Y, Capri FC, Mineo A, Cosenza A, Gallo G, Alduina R, Mannina G. Initial pH Conditions Shape the Microbial Community Structure of Sewage Sludge in Batch Fermentations for the Improvement of Volatile Fatty Acid Production. Microorganisms. 2022; 10(10):2073. https://doi.org/10.3390/microorganisms10102073
Chicago/Turabian StyleDi Leto, Ylenia, Fanny Claire Capri, Antonio Mineo, Alida Cosenza, Giuseppe Gallo, Rosa Alduina, and Giorgio Mannina. 2022. "Initial pH Conditions Shape the Microbial Community Structure of Sewage Sludge in Batch Fermentations for the Improvement of Volatile Fatty Acid Production" Microorganisms 10, no. 10: 2073. https://doi.org/10.3390/microorganisms10102073