
Electrochemical Enrichment and Isolation of Electrogenic Bacteria from 0.22 µm Filtrate

Abstract
:1. Introduction
2. Materials and Methods
2.1. Medium and Electrolyte Composition
2.2. Electrochemical Enrichment and Isolation of 0.22 μm Filter–Passable Bacteria
2.3. DNA Extraction and 16S rRNA Gene Phylogenetic Analysis
2.4. Comparison of the Current Production of the Isolated Strain with Related Species and Differential Pulse Voltammetry (DPV) Analysis
2.5. Evaluation of Cell Size under Various Pure Culture Conditions Using Scanning Electron Microscopy (SEM) and Filtration
2.6. Whole-Genome Sequence of Isolated Strain and Genetic Characterization
3. Results
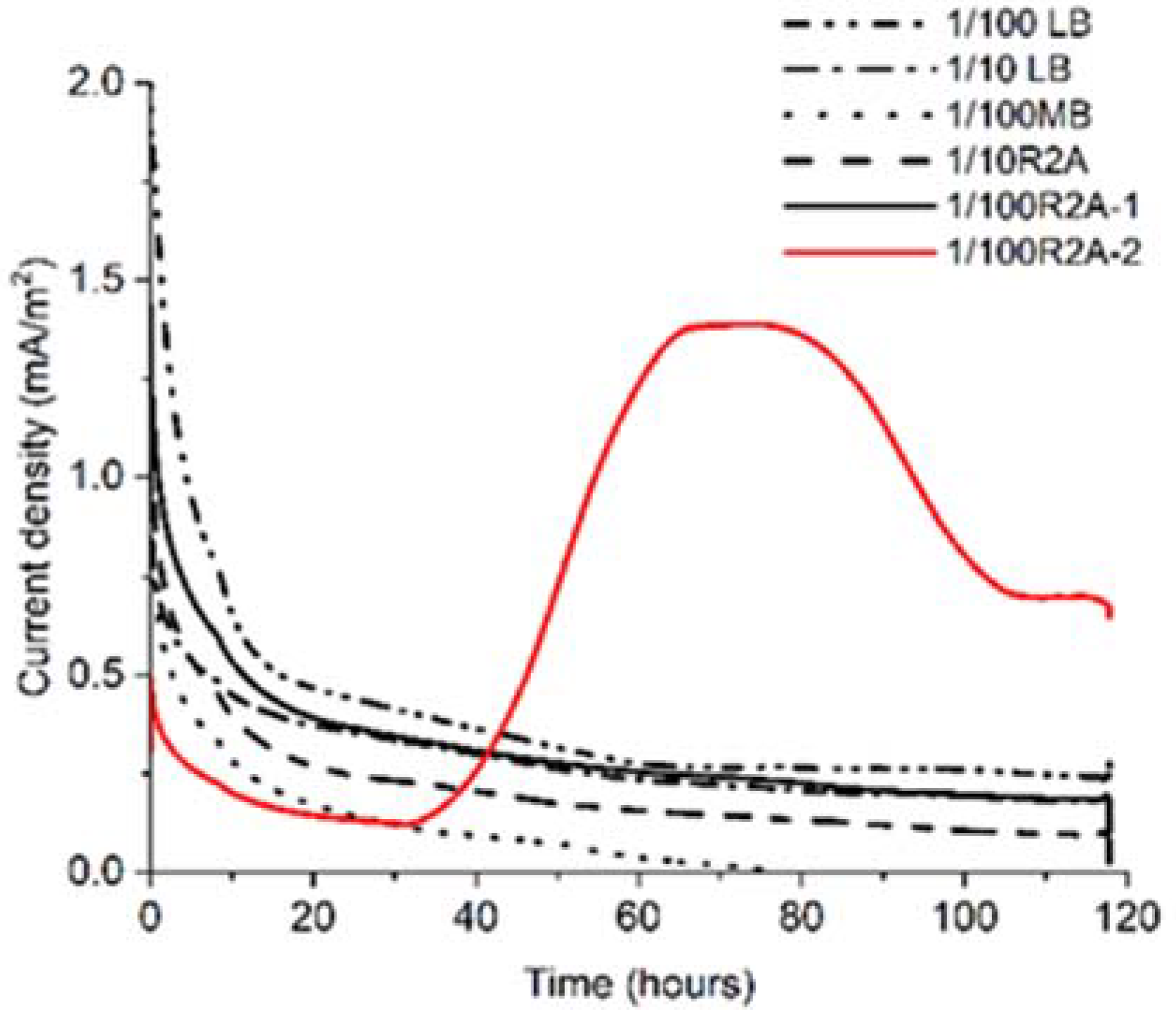
3.1. Electrochemical Enrichment and Isolation of 0.22 μm Filter–Passable Bacteria
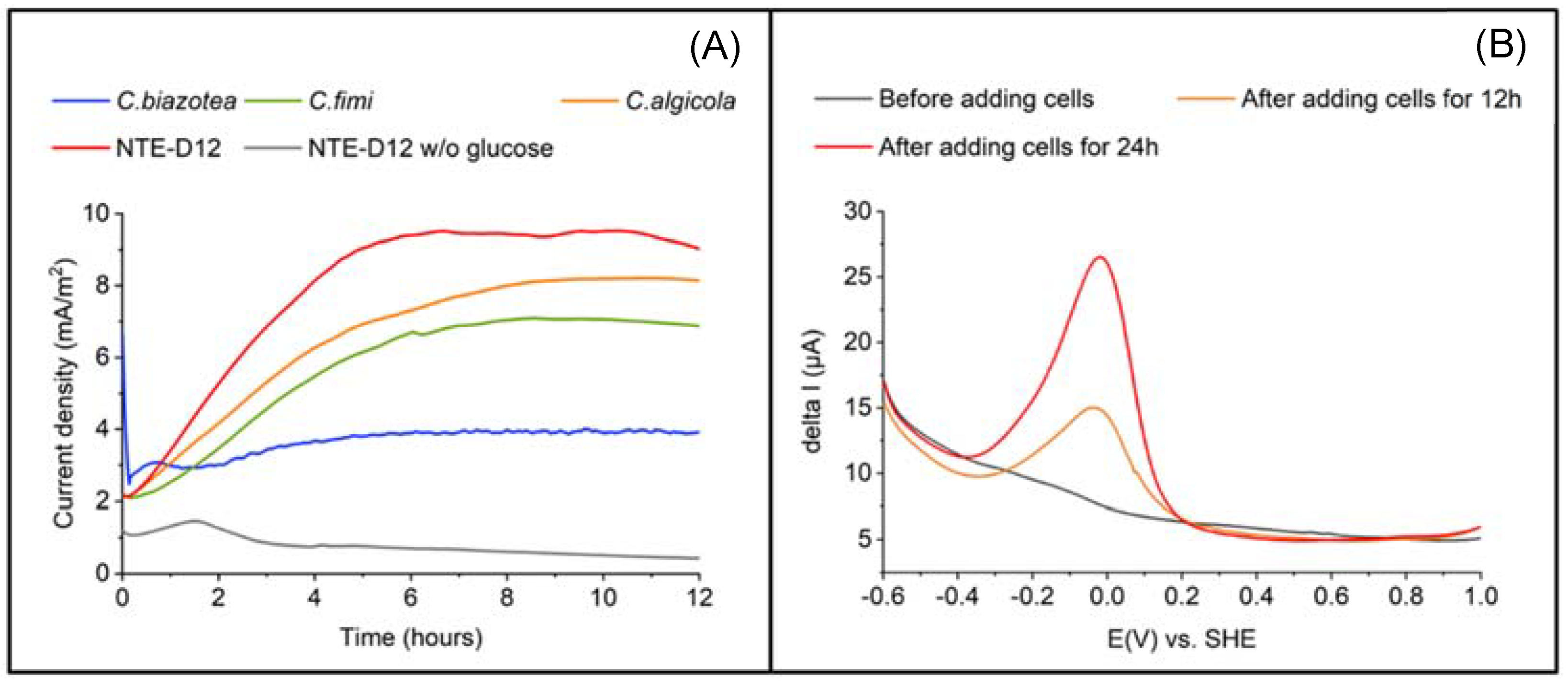
3.2. Electrochemical Characterization of the Isolated Strain
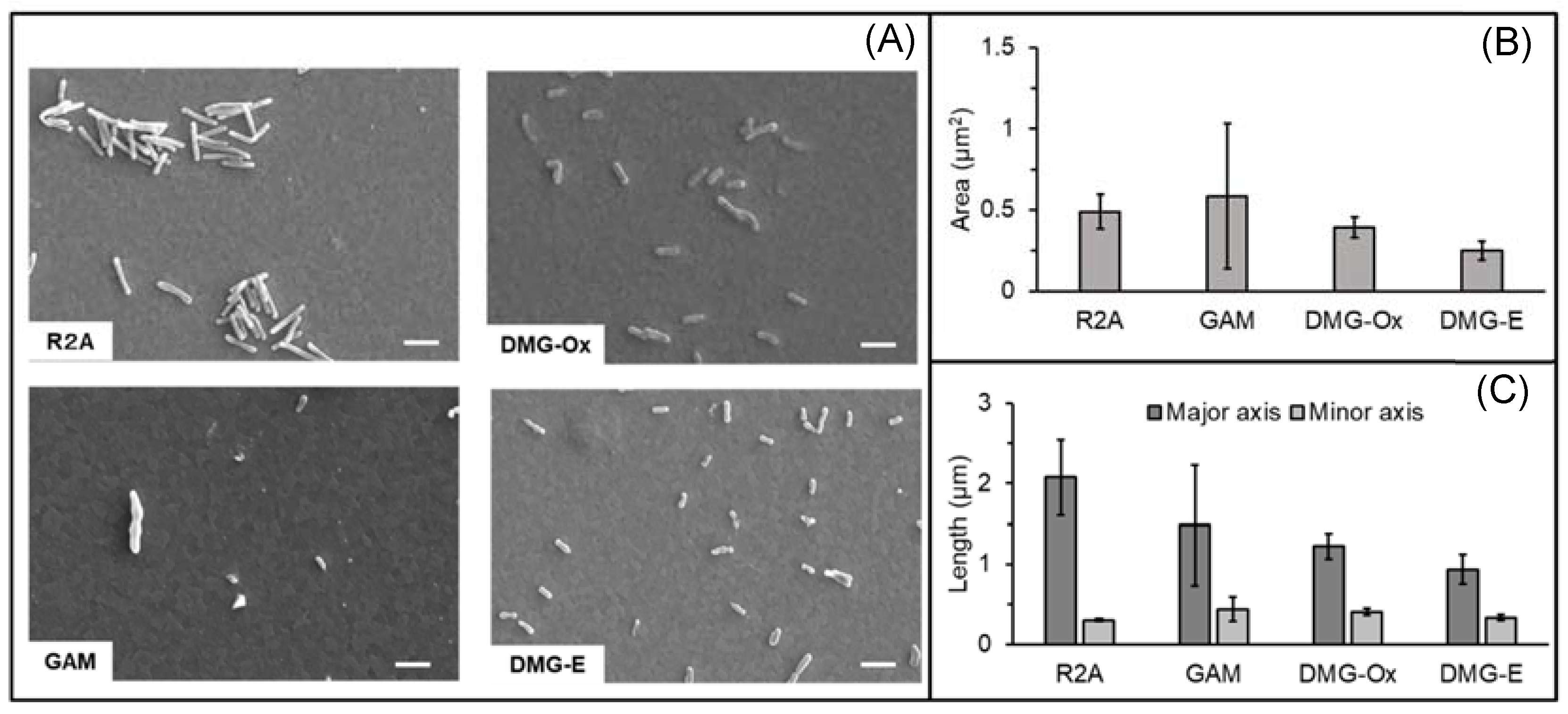
3.3. Evaluation of Cell Size in Various Culture Conditions
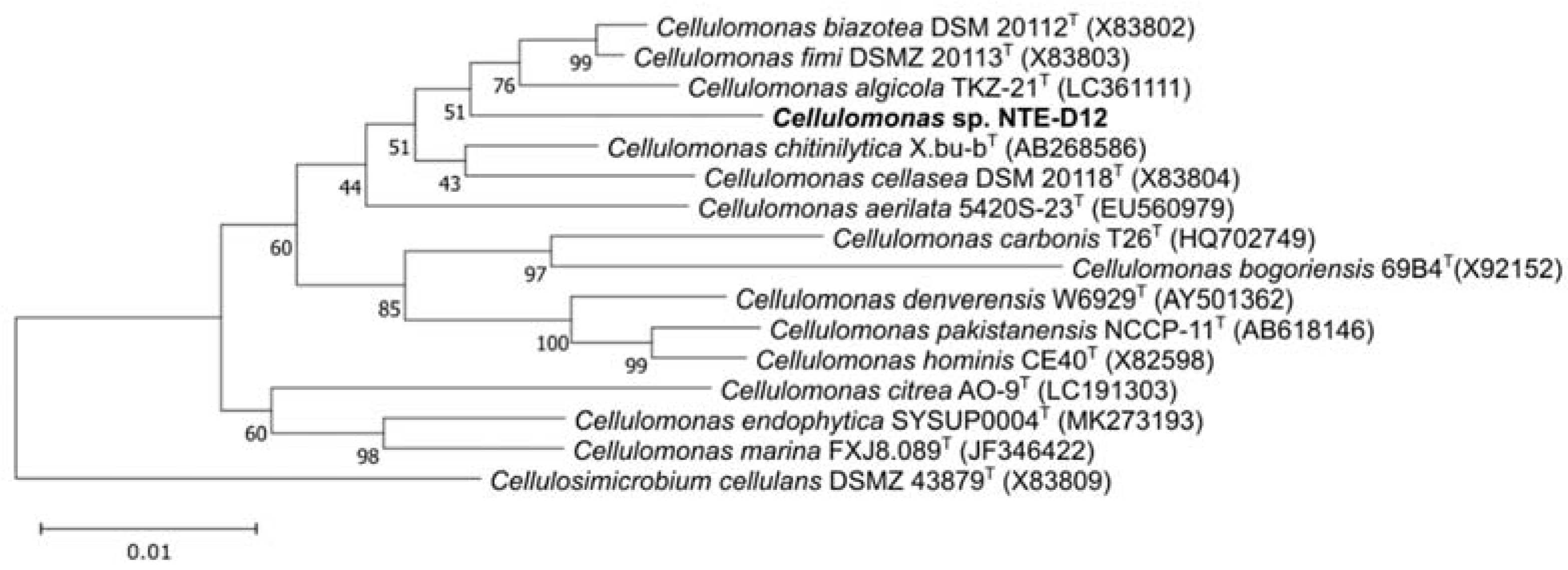
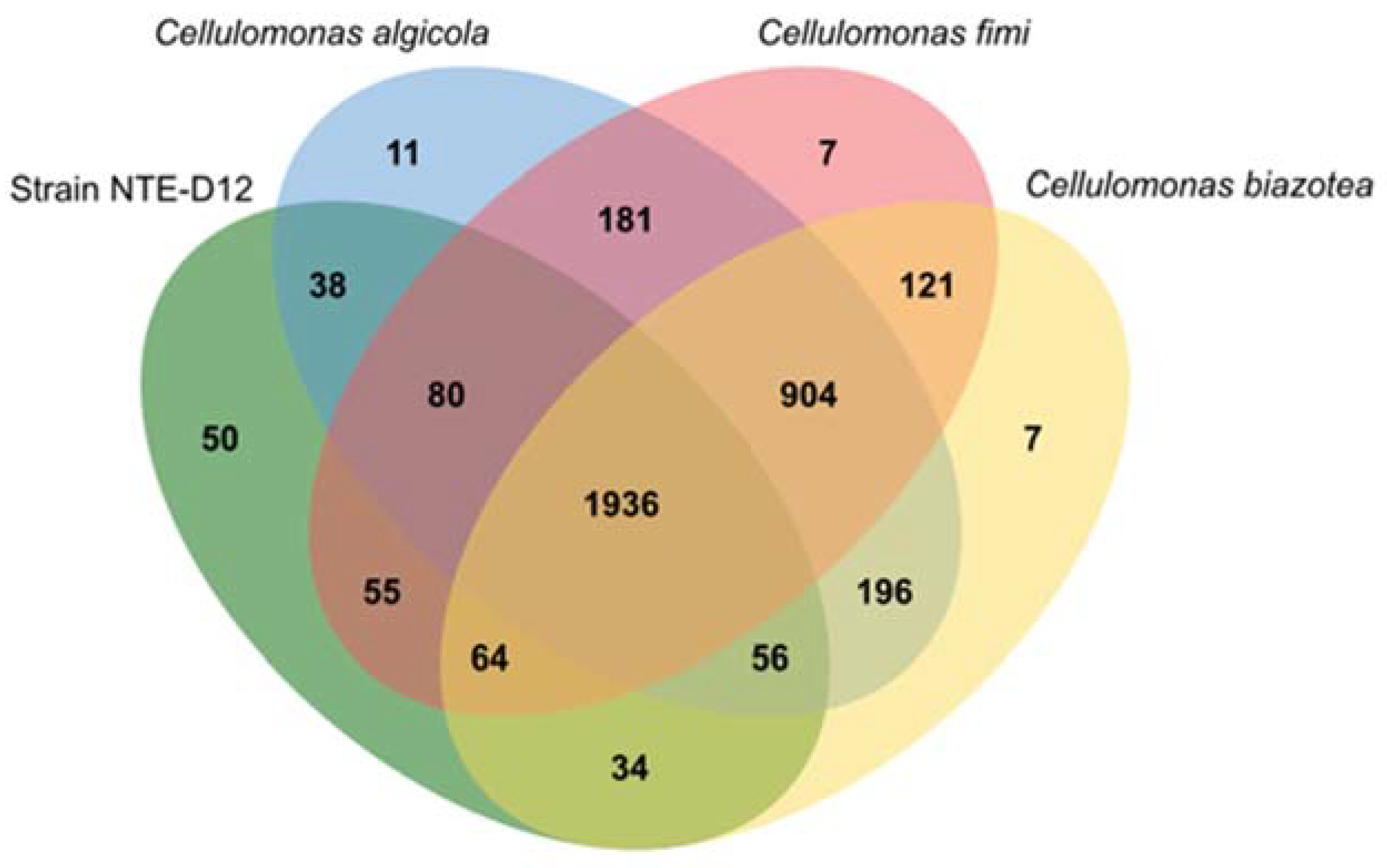
3.4. Genomic Features of the Isolated Strain and Comparison with Related Species
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- MacDonell, M.T.; Hood, M.A. Isolation and Characterization of Ultramicrobacteria from a Gulf Coast Estuary. Appl. Environ. Microbiol. 1982, 43, 566–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naganuma, T.; Miyoshi, T.; Kimura, H. Phylotype diversity of deep-sea hydrothermal vent prokaryotes trapped by 0.2- and 0.1-μm-pore-size filters. Extremophiles 2007, 11, 637–646. [Google Scholar]
- Nakai, R.; Abe, T.; Takeyama, H.; Naganuma, T. Metagenomic Analysis of 0.2-μm-Passable Microorganisms in Deep-Sea Hydrothermal Fluid. Mar. Biotechnol. 2011, 13, 900–908. [Google Scholar]
- Brown, C.T.; Hug, L.A.; Thomas, B.C.; Sharon, I.; Castelle, C.J.; Singh, A.; Wilkins, M.J.; Wrighton, K.C.; Williams, K.H.; Banfield, J.F. Unusual biology across a group comprising more than 15% of domain Bacteria. Nature 2015, 523, 208–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hug, L.A.; Baker, B.; Anantharaman, K.; Brown, C.T.; Probst, A.; Castelle, C.; Butterfield, C.N.; Hernsdorf, A.W.; Amano, Y.; Ise, K.; et al. A new view of the tree of life. Nat. Microbiol. 2016, 1, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duda, V.I.; Suzina, N.E.; Polivtseva, V.N.; Boronin, A.M. Ultramicrobacteria: Formation of the concept and contribution of ultramicrobacteria to biology. Microbiology 2012, 81, 379–390. [Google Scholar] [CrossRef]
- Suzuki, S.; Horinouchi, S.; Beppu, T. Growth of a Tryptophanase-producing Thermophile, Symbiobacterium thermophilum gen. nov., sp. nov., Is Dependent on Co-culture with a Bacillus Sp. Microbiology 1988, 134, 2353–2362. [Google Scholar]
- Morris, J.J.; Lenski, R.E.; Zinser, E.R. The black queen hypothesis: Evolution of dependencies through adaptive gene loss. mBio 2012, 3. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, K.; Manefield, M.; Lee, M.; Kouzuma, A. Electron shuttles in biotechnology. Curr. Opin. Biotechnol. 2009, 20, 633–641. [Google Scholar] [CrossRef]
- Summers, Z.M.; Fogarty, H.E.; Leang, C.; Franks, A.E.; Malvankar, N.S.; Lovley, D.R. Direct exchange of electrons within aggregates of an evolved syntrophic coculture of anaerobic bacteria. Science 2010, 330, 1413–1415. [Google Scholar] [CrossRef] [Green Version]
- Kato, S.; Hashimoto, K.; Watanabe, K. Microbial interspecies electron transfer via electric currents through conductive minerals. Proc. Natl. Acad. Sci. USA 2012, 109, 10042–10046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovley, D.R. Syntrophy Goes Electric: Direct Interspecies Electron Transfer. Annu. Rev. Microbiol. 2017, 71, 643–664. [Google Scholar] [CrossRef]
- Luef, B.; Frischkorn, K.R.; Wrighton, K.C.; Holman, H.-Y.N.; Birarda, G.; Thomas, B.C.; Singh, A.; Williams, K.H.; Siegerist, C.E.; Tringe, S.G.; et al. Diverse uncultivated ultra-small bacterial cells in groundwater. Nat. Commun. 2015, 6, 6372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garber, A.I.; Nealson, K.H.; Okamoto, A.; McAllister, S.M.; Chan, C.S.; Barco, R.A.; Merino, N. FeGenie: A Comprehensive Tool for the Identification of Iron Genes and Iron Gene Neighborhoods in Genome and Metagenome Assemblies. Front. Microbiol. 2020, 11, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conley, B.E.; Intile, P.J.; Bond, D.R.; Gralnick, J.A. Divergent Nrf family proteins and MtrCAB homologs facilitate extracellular electron transfer in Aeromonas hydrophila. Appl. Environ. Microbiol. 2018, 84, e02134-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conley, B.E.; Weinstock, M.T.; Bond, D.R.; Gralnick, J.A. A hybrid extracellular electron transfer pathway enhances the survival of vibrio natriegens. Appl. Environ. Microbiol. 2020, 86, e01253-20. [Google Scholar] [CrossRef]
- Rowe, A.R.; Yoshimura, M.; LaRowe, D.E.; Bird, L.J.; Amend, J.P.; Hashimoto, K.; Nealson, K.H.; Okamoto, A. In situ electrochemical enrichment and isolation of a magnetite-reducing bacterium from a high pH serpentinizing spring. Environ. Microbiol. 2017, 19, 2272–2285. [Google Scholar] [CrossRef] [Green Version]
- Naradasu, D.; Miran, W.; Sakamoto, M.; Okamoto, A. Isolation and characterization of human gut bacteria capable of extracellular electron transport by electrochemical techniques. Front. Microbiol. 2019, 10, 3267. [Google Scholar] [CrossRef]
- Okamoto, A.; Nakamura, R.; Hashimoto, K. In-vivo identification of direct electron transfer from Shewanella oneidensis MR-1 to electrodes via outer-membrane OmcA-MtrCAB protein complexes. Electrochim. Acta 2011, 56, 5526–5531. [Google Scholar] [CrossRef]
- Miran, W.; Long, X.; Huang, W.; Okamoto, A. Current Production Capability of Drug-Resistant Pathogen Enables Its Rapid Label-Free Detection Applicable to Wastewater-Based Epidemiology. Microorganisms 2022, 10, 472. [Google Scholar] [CrossRef]
- Hiraishi, A.; Shin, Y.K.; Ueda, Y.; Sugiyama, J. Automated sequencing of PCR-amplified 16S rDNA on ‘Hydrolink’ gels. J. Microbiol. Methods 1994, 19, 145–154. [Google Scholar] [CrossRef]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Tokunou, Y.; Hashimoto, K.; Okamoto, A. Acceleration of Extracellular Electron Transfer by Alternative Redox-Active Molecules to Riboflavin for Outer-Membrane Cytochrome c of Shewanella oneidensis MR-1. J. Phys. Chem. C 2016, 120, 16168–16173. [Google Scholar] [CrossRef]
- Okamoto, A.; Hashimoto, K.; Nealson, K.H.; Nakamura, R. Rate enhancement of bacterial extracellular electron transport involves bound flavin semiquinones. Proc. Natl. Acad. Sci. USA 2013, 110, 7856. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [Green Version]
- Yu, N.Y.; Wagner, J.R.; Laird, M.R.; Melli, G.; Rey, S.; Lo, R.; Dao, P.; Sahinalp, S.C.; Ester, M.; Foster, L.J.; et al. PSORTb 3.0: Improved protein subcellular localization prediction with refined localization subcategories and predictive capabilities for all prokaryotes. Bioinformatics 2010, 26, 1608–1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Dong, Z.; Fang, L.; Luo, Y.; Wei, Z.; Guo, H.; Zhang, G.; Gu, Y.Q.; Coleman-Derr, D.; Xia, Q.; et al. OrthoVenn2: A web server for whole-genome comparison and annotation of orthologous clusters across multiple species. Nucleic Acids Res. 2019, 47, W52–W58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamura, H.; Hayashi, T.; Hamada, M.; Kohda, T.; Serisawa, Y.; Matsuyama-Serisawa, K.; Nakagawa, Y.; Otoguro, M.; Yanagida, F.; Tamura, T.; et al. Cellulomonas algicola sp. nov., an actinobacterium isolated from a freshwater alga. Int. J. Syst. Evol. Microbiol. 2019, 69, 2723–2728. [Google Scholar] [CrossRef] [PubMed]
- Light, S.H.; Su, L.; Rivera-Lugo, R.; Cornejo, J.A.; Louie, A.; Iavarone, A.T.; Ajo-Franklin, C.M.; Portnoy, D.A. A flavin-based extracellular electron transfer mechanism in diverse Gram-positive bacteria. Nature 2018, 562, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Fukuba, T.; Sato, M.; Elsaied, E.H.; Naganuma, T. Overlooked Microbial Agents in Aquaculture: Nanobacteria. In Microbial Approaches to Aquatic Nutrition within Environmentally Sound Aquaculture Production Systems; World Aquaculture Society: Becton Rouge, LA, USA, 2002; pp. 99–107. [Google Scholar]
- Ho, C.; Emran, M.Y.; Ihara, S.; Huang, W.; Wakai, S.; Li, W.P.; Okamoto, A. Osmium-Grafted Magnetic Nanobeads Increase Microbial Current Generation Via Culture-Free and Quick Enrichment of Electrogenic Bacteria. Available online: https://papers.ssrn.com/sol3/papers.cfm?abstract_id=4177599 (accessed on 1 August 2022).
- Nakai, R.; Shibuya, E.; Justel, A.; Rico, E.; Quesada, A.; Kobayashi, F.; Iwasaka, Y.; Shi, G.-Y.; Amano, Y.; Iwatsuki, T.; et al. Phylogeographic analysis of filterable bacteria with special reference to Rhizobiales strains that occur in cryospheric habitats. Antarct. Sci. 2013, 25, 219–228. [Google Scholar] [CrossRef]
- Kjelleberg, S.; Albertson, N.; Flärdh, K.; Holmquist, L.; Jouper-Jaan, Å.; Marouga, R.; Östling, J.; Svenblad, B.; Weichart, D. How do non-differentiating bacteria adapt to starvation? Antonie Van Leeuwenhoek 1993, 63, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Monier, J.M.; Lindow, S.E. Pseudomonas syringae Responds to the Environment on Leaves by Cell Size Reduction. Phytopathology 2003, 93, 1209–1216. [Google Scholar] [CrossRef] [Green Version]
- Eguchi, M.; Nishikawa, T.; Macdonald, K.; Cavicchioli, R.; Gottschal, J.C.; Kjelleberg, S. Responses to Stress and Nutrient Availability by the Marine Ultramicrobacterium Sphingomonas sp. Strain RB2256. Appl. Environ. Microbiol. 1996, 62, 1287–1294. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Richardson, D.J.; Wang, Z.; Kerisit, S.N.; Rosso, K.M.; Zachara, J.M.; Fredrickson, J.K. The roles of outer membrane cytochromes of Shewanella and Geobacter in extracellular electron transfer. Environ. Microbiol. Rep. 2009, 1, 220–227. [Google Scholar] [CrossRef]
- Deng, X.; Dohmae, N.; Nealson, K.H.; Hashimoto, K.; Okamoto, A. Multi-heme cytochromes provide a pathway for survival in energy-limited environments. Sci. Adv. 2018, 4, eaao5682. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cellulomonas sp. Strain NTE-D12 | C. algicola Strain NBRC112905T | C. fimi Strain NBRC15513T | C. biazotea Strain NBRC12680T | |
|---|---|---|---|---|
| GenBank accession No. | - | GCA_003851725.1 | GCA_000212695.1 | GCA_004306155.1 |
| Genome size (Mb) | 3.68 | 4.5 | 4.27 | 4.38 |
| CDS number | 3333 | 4047 | 3812 | 3911 |
| Iron-reduction gene | N.D. | N.D. | N.D. | N.D. |
| T4P-related gene | N.D. | 1 | 1 | 1 |
| Terminal oxidase-related gene | 5 | 6 | 6 | 4 |
| Ubiquinone biosynthesis gene | N.D. | 5 | 5 | 3 |
| Genes with heme-binding motif localized at | ||||
| Cytoplasmic membrane | 6 | 4 | 4 | 5 |
| Cell wall | 1 | N.D. | N.D. | N.D. |
| Flavoprotein localized at | ||||
| Cytoplasmic membrane | 2 | 3 | 1 | 2 |
| Cell wall | N.D. | N.D. | N.D. | N.D. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ihara, S.; Wakai, S.; Maehara, T.; Okamoto, A. Electrochemical Enrichment and Isolation of Electrogenic Bacteria from 0.22 µm Filtrate. Microorganisms 2022, 10, 2051. https://doi.org/10.3390/microorganisms10102051
Ihara S, Wakai S, Maehara T, Okamoto A. Electrochemical Enrichment and Isolation of Electrogenic Bacteria from 0.22 µm Filtrate. Microorganisms. 2022; 10(10):2051. https://doi.org/10.3390/microorganisms10102051
Chicago/Turabian StyleIhara, Sota, Satoshi Wakai, Tomoko Maehara, and Akihiro Okamoto. 2022. "Electrochemical Enrichment and Isolation of Electrogenic Bacteria from 0.22 µm Filtrate" Microorganisms 10, no. 10: 2051. https://doi.org/10.3390/microorganisms10102051