
Gut Microbiota-Derived Short-Chain Fatty Acids: Impact on Cancer Treatment Response and Toxicities
 , , and
, , and
Abstract
:1. Introduction
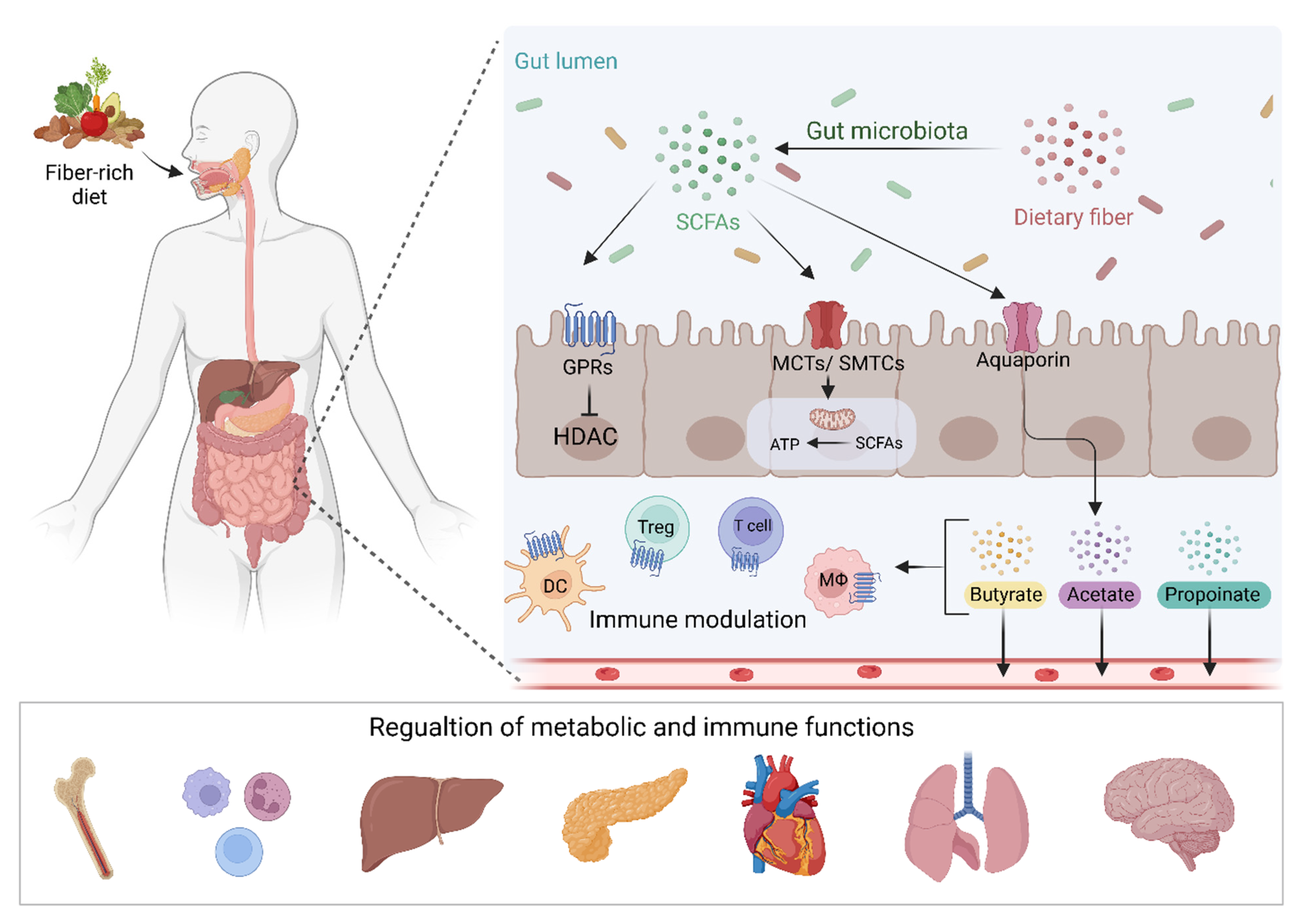
2. Short-Chain Fatty Acids
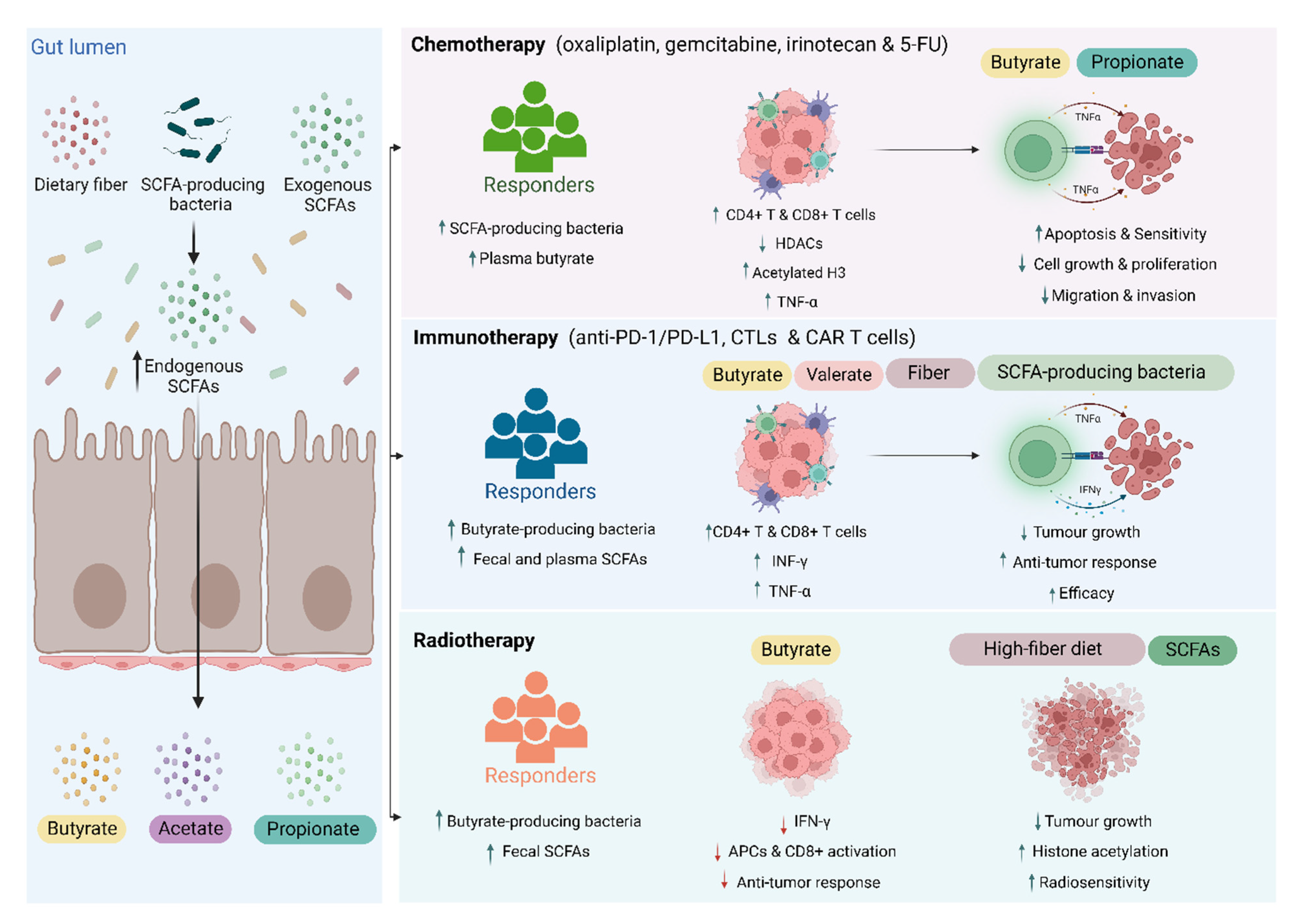
3. SCFAs and Cancer Treatment Response
3.1. Chemotherapy

{kind=link}
{kind=link}
{kind=link}
| Ref. | Subjects/Model | Treatment | Key Findings |
|---|---|---|---|
| Geng et al. [44] | HCT116 human colorectal cancer cell line | 5-FU +/− butyrate | Butyrate enhanced 5-FU-induced apoptosis on colorectal cancer cells Butyrate improved cell sensitivity to 5-FU by augmenting 5-FU-induced inhibition of DNA synthesis. |
| Encarnação et al. [45] | WiDr, C2BBe1, and LS1034 colorectal cancer cells WiDr colorectal mouse model | Irinotecan +/− butyrate | In vitro, butyrate reduced the IC50 of irinotecan by enhancing cancer cell apoptosis and reducing proliferation. Butyrate significantly decreased the expression of chemoresistant-related protein. In vivo, butyrate delayed tumor growth following irinotecan treatment. |
| Panebianco et al. [46] | BxPC-3 and PANC-1 pancreatic cancer cell line BxPC-3 pancreatic cancer mouse model | Gemcitabine +/− butyrate | In vitro, butyrate administration enhanced gemcitabine-induced inhibition of cancer cell growth and apoptosis. In vivo, butyrate did not affect tumor volume but suppressed stromatogenesis by reducing the density of stroma collagen bundles, and expression of myofibroblasts, vascular architecture, and M2-polarized macrophage markers in tumors. |
| Li et al. [47] | HGC-27 and SGC-7901 gastric cancer cell lines SGC-7901 gastric cancer mouse model | Cisplatin +/− butyrate | Butyrate synergized cisplatin-induced tumor cell apoptosis by increasing mitochondrial ROS levels and mitochondrial membrane potential. Suppressed cell migration and invasion in vitro by reducing the levels of MMP-2, -9 proteins. Cisplatin augmented cisplatin-induced suppression of tumor growth by increasing the levels of apoptosis makers. |
| Kobayashi et al. [48] | HepG2, HuH-7, JHH-4 hepatocellular carcinoma cell lines HepG2 human hepatocellular carcinoma mouse model | Cisplatin +/− propionate | In vitro, combined therapy inhibited proliferation and enhanced apoptosis through the GPR41 signaling pathway. Enhanced levels of activated DNA fragmentation markers (cleaved caspase-3). Enhanced the expression of TNF-α by downregulating the expression of HDACs and enhancing histone H3 acetylation. In vivo, combined therapy suppressed tumor growth and enhanced histone H3 acetylation and mRNA expression of TNF-α. |
| He et al. [43] | MC38 colon cancer/ EG7 lymphoma mouse model Patients with gastrointestinal cancer (n= 21) | Oxaliplatin +/− butyrate | Butyrate augmented oxaliplatin efficacy by enhancing the anti-tumor activity of CD8+ T cells in an IL-12 signaling pathway-dependent manner. Butyrate prompted cytotoxic CD8+ T cell anti-tumor responses in vitro and in vivo through the IL-12 signaling pathway. Clinically, responders to oxaliplatin had higher concentrations of serum butyrate compared to non-responders. |
| Zhang et al. [49] | MC38 colon cancer mouse model | Anti-PD-1 +/− butyrate | Pre- immunotherapy butyrate supplement improved anti-PD-1 efficacy in mice humanized with gut microbiota from CRC patients Butyrate increased infiltration of tumor-killing CD4+ and CD8+ cells in the tumor. |
| Luu et al. [10] | B16-OVA melanoma/ PancOVA pancreatic cancer mouse model | CTLs and CAR T cells +/− valerate / butyrate | Transferring valerate or butyrate-treated cytotoxic T cells and chimeric antigen receptor T cells into tumor-bearing mice increased the production of CD25, IFN-γ, and TNF-α and enhances the anti-tumor activity. |
| Jing et al. [50] | MC38 colon adenocarcinoma mouse model | Anti-PD-L1 +/− fiber-rich powder or SCFAs | Administrating fiber-rich powder improved anti-PD-L1 efficacy by increasing the production of acetate, propionate, butyrate, and valerianate. Oral butyrate supplements did not affect anti-PD-L1 efficiency or total leukocytes and CD8+ T cell proportion. Oral SCFAs did not increase cecum or colon SCFA levels In vitro, culturing lymphocytes with SCFAs increased the proportion of CD8+ T cells. |
| Han et al. [51] | CT26 colon cancer mouse model | Anti-PD-1 +/− Inulin (dietary fiber) or SCFAs | Inulin improved anti-tumor response and delayed tumor growth by increasing systemic tumor-specific CD8+ cells count. Inulin increased SCFA-producing Lactobacillus, Akkermansia, and Roseburia, and fecal SCFAs. Negative correlation between the tumor size and the fecal propionate and butyrate. Inulin increased systemic tumor-specific CD8+ cells, splenic IFN-γ+ CD8+ T cells, intratumoral CD8+, CD4+, activated dendritic cells and decreased PD-1 positive CD8+ cells. Oral administration of free SCFAs did not improve the anti-tumor response. Antibiotic treatment and GPR43 knockdown abrogated anti-tumor activity. In vitro, SCFAs enhanced the memory response of IFN-γ+ CD8+ T cells and upregulated T-cell factor 1. |
| Spencer et al. [52] | Patients with metastatic melanoma (n = 128) B2905 and HMel melanoma mouse model | Anti-PD-1 +/− fiber-rich diet | High fiber dietary intake improved progression-free survival in patients. Fiber-rich diet delayed tumor growth in mice. Fiber-rich diet had no impact on tumor response in germ-free mice. Higher propionate levels were observed in mice receiving a fiber-rich diet. Fiber-rich diet increased the number of tumor-infiltrating CD4+ cells and IFNγ+ cytotoxic T cells. |
| Coutzac et al. [12] | Patients with metastatic melanoma (n = 85) CT26 and MC38 Colon/ MCA101OVA fibrosarcoma mouse model | Ipilimumab +/− systemic butyrate | Higher serum levels of propionate and butyrate were associated with poor clinical outcome and high serum SCFAs was positively correlated to the proportion of Tregs. Butyrate administration reduced treatment efficacy by suppressing dendritic cell maturation and decreased T-cell expansion and functions in mice. |
| Tomita et al. [53,54] | Patients with advanced non–small cell lung cancer received antibiotics or PPI (n = 118) | Immune checkpoint blockade +/− C. butyricum | C. butyricum was associated with longer progression-free and overall survival, particularly in patients who received PPI with/without antibiotics. C. butyricum administration increased beneficial microbiota and reduced oral-related pathobionts in the gut. |
| Then et al. [55] | RT112 bladder carcinoma mouse model RT112 bladder cell line | Radiation+ fiber-containing diet Cells irradiation +/− SCFAs | Soluble high-fiber diet delayed tumor growth following irradiation Soluble high-fiber diet increased acetate-producing Bacteroides acidifaciens, which was associated with better radiation response and long survival. In vitro, SCFAs increased histone acetylation and reduced cell proliferation while butyrate only significantly enhanced radiosensitivity. |
| Yang et al. [56] | MC38 colon/ B16F1 melanoma mouse model | Local radiation +/− Systemic or intratumoral butyrate | Both systemic and intratumoral butyrate impaired anti-tumor response in MC38 and B16F1 models. Intratumoral butyrate did not directly protect tumor cells from radiation but inhibited radiation-induced anti-tumor immune responses. Intratumoral butyrate inhibited type I IFN expression in dendritic cells and hence suppressed dendritic cell functions and activation of CD8+ T cell immune responses. |
| Uribe-Herranz et al. [57] | B16OVA melanoma/ TC-1 lung cancer mouse model | Local irradiation +/− butyrate | Depletion of butyrate-producing taxa and reducing tumor butyrate levels by vancomycin improved anti-tumor activity. Butyrate administration reduces antigen-presenting cells activation and functions. |
3.2. Immunotherapy
3.3. Radiotherapy
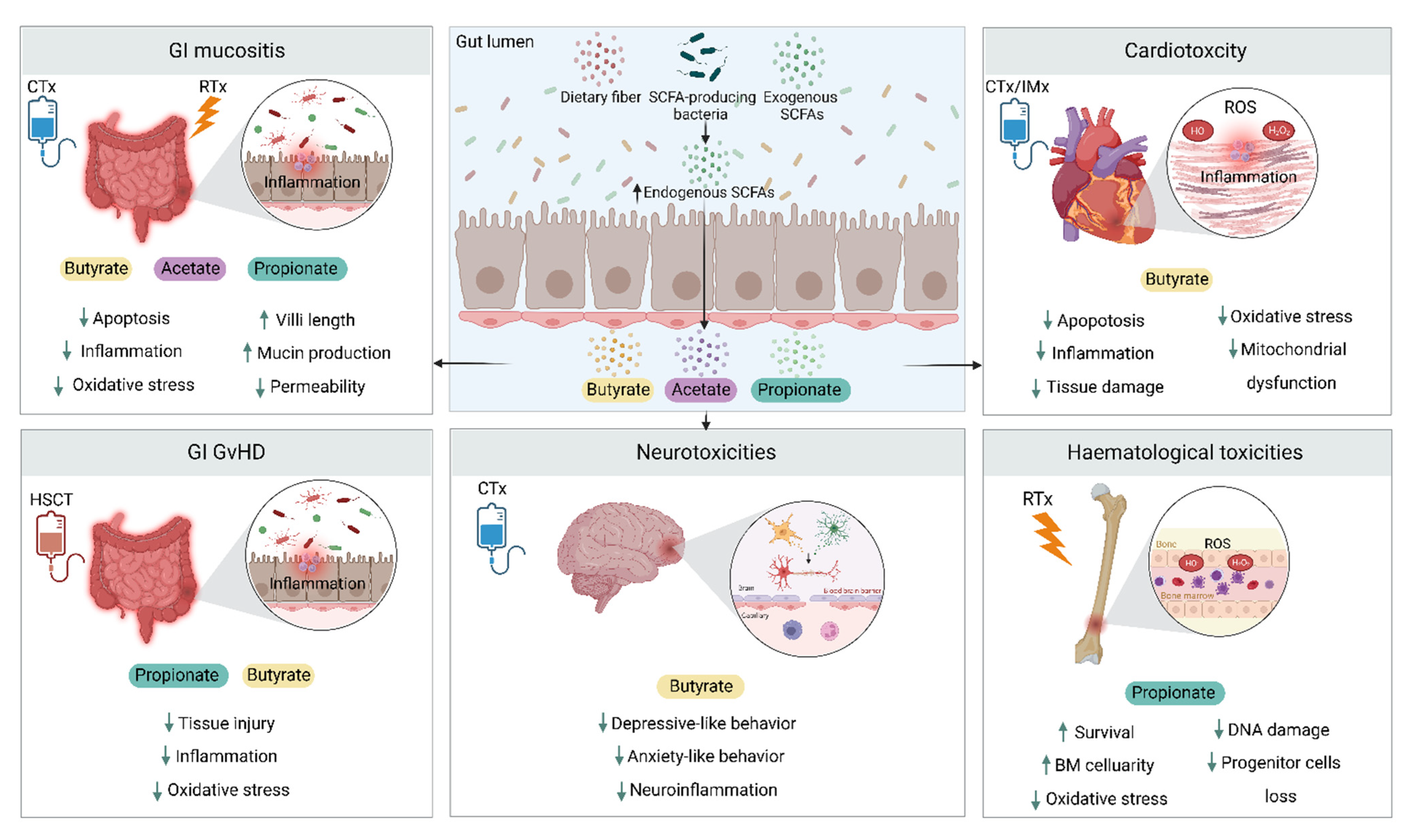
4. SCFAs and Cancer Treatment Toxicities
| Ref. | Subjects/Model | Treatment | Toxicity | Key Findings |
|---|---|---|---|---|
| da Silva Ferreira et al. [68] | 3D intestinal organoids | Methotrexate +/− SCFAs | GI toxicity | Butyrate and propionate reduced the reduction in metabolic activity caused by methotrexate. |
| Ferreira et al. [69] | Mice | 5-FU +/− SCFAs/ butyrate | GI toxicity | SCFA and Butyrate reduced or prevented the 5-FU-induced reduction in body weight and intestinal length. SCFA and Butyrate reduced tissue damage and mucosal ulceration in the small intestine. SCFA and Butyrate had no effect on inflammatory infiltrates but prevented 5-FU induced increase in EPO enzyme activity (a marker for eosinophil infiltration). Butyrate decreased intestinal permeability and ZO-1 expression associated with 5-FU. |
| Yue et al. [70] | THP-1 cells and Caco-2 cells Mice | 5-FU +/− SCFAs/ L. rhamnoides | GI toxicity | SCFAs (in vitro): Suppressed ROS production Reduced expression of NLRP3 and proinflammatory cytokines Reduce autophagy markers. Lactobacillus rhamnoides (in vivo): Increased fecal SCFAs Increased serum IL-1β, IL-6 and IgA Decreased splenic NLRP3 and IL-17 Increased intestinal ZO-1 and occludin |
| Wang et al. [71] | Colon carcinoma-bearing mice | 5-FU +/− Carboxymethylated pachyman (modified polysaccharide) | GI toxicity | Acetate, propionate, and butyrate reduced while isobutyrate and isovalerate increased following 5-FU. Carboxymethylated pachyman restored normal levels of SCFAs Intervention reduced 5-FU-induced intestinal tissue injury, apoptosis, and inflammation. |
| Panebianco et al. [46] | Pancreatic adenocarcinoma-bearing mice | Gemcitabine +/− butyrate | GI toxicity | Butyrate attenuated toxicity by protecting villi structure, increasing mucin production, and enrichment of anti-inflammatory SCFA-producing microbiota. |
| Guo et al. [72] | Mice | TBI (8 – 8.2 Gy) +/− SCFAs | Haemopoietic + GI toxicity | Radiation-resistant mice had higher concentrations of fecal total SCFA and propionate. Propionate enhanced the survival rate. Propionate increased bone marrow cellularity and splenic pulp recovery and reduced the radiation-induced loss of hematopoietic progenitor cells. Propionate increased crypt length and mucus thickness. SCFAs attenuated DNA damage and reactive oxygen species production in hematopoietic and gastrointestinal tissues |
| Huang et al. [73] | Mice | Doxorubicin +/− sodium butyrate | Cardiotoxicity | Doxorubicin reduced levels of fecal and serum butyrate Butyrate increased arginase-1 and CD206 levels and decreased cardiomyocyte apoptosis and myocardial enzymes. Butyrate promotes the polarization of colonic anti-inflammatory M2 macrophages. |
| Russo et al. [74] | Mice Cardiomyocytes/ endothelial cells | Doxorubicin +/−butyrate derivative (phenylalanine-butyramide (FBA)) | Cardiotoxicity | FBA: Reduced Doxorubicin-induced left ventricle dilatation and volume Prevented fibrosis, apoptosis, and reduction in cardiomyocyte size Reduced expression of cardiac dysfunction and remodeling markers Reduced oxidative stress markers and prevented mitochondrial dysfunction Prevented cell damage and apoptosis. |
| Chen et al. [75] | Melanoma-bearing mice | PD-1/PD-L1 inhibitor +/− P. loescheii/ butyrate | Cardiotoxicity | Lower fecal butyrate in cardiotoxicity model SCFA-producing bacteria (P. loescheii) or butyrate reduced myocardial apoptosis and serum myocardial enzymes. P. loescheii and butyrate downregulated proinflammatory factors in the colonic and cardiac tissues. |
| Mathewson et al. [76] | Mice | BMT +/− butyrate / butyrate-producing Clostridia strains | GI GVHD | BMT reduced intestinal butyrate and reduced histone acetylation in IECs. Butyrate restored histone acetylation, reduced apoptosis, and enhanced tight junction integrity. Butyrate + Clostridia strains attenuated GI GVHD severity. |
| Cristiano et al. [77] | Mice | Paclitaxel +/− sodium butyrate | GI + behavioral dysfunctions | Paclitaxel reduced intestinal barrier integrity, caused microbial dysbiosis, and decreased fecal butyrate. Oral butyrate attenuated disruption of intestinal barrier integrity and microbial dysbiosis. Oral butyrate attenuated depressive and anxiety-like behavior and neuroinflammation. |
4.1. Gastrointestinal Toxicity
4.2. Cardiotoxicity
4.3. Hematological Toxicities
4.4. Graft Versus Host Disease
4.5. Psychoneurological Toxicities
5. Targeting SCFA to Improve Cancer Treatment Outcomes
6. Current Limitations and Future Considerations
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gopalakrishnan, V.; Spencer, C.N.; Nezi, L.; Reuben, A.; Andrews, M.C.; Karpinets, T.V.; Prieto, P.A.; Vicente, D.; Hoffman, K.; Wei, S.C.; et al. Gut microbiome modulates response to anti-PD-1 immunotherapy in melanoma patients. Science 2018, 359, 97–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, J.L.; Wilson, I.D.; Teare, J.; Marchesi, J.R.; Nicholson, J.K.; Kinross, J.M. Gut microbiota modulation of chemotherapy efficacy and toxicity. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 356–365. [Google Scholar] [CrossRef] [PubMed]
- Chaput, N.; Lepage, P.; Coutzac, C.; Soularue, E.; Le Roux, K.; Monot, C.; Boselli, L.; Routier, E.; Cassard, L.; Collins, M.; et al. Baseline gut microbiota predicts clinical response and colitis in metastatic melanoma patients treated with ipilimumab. Ann. Oncol. 2017, 28, 1368–1379. [Google Scholar] [CrossRef] [PubMed]
- Sims, T.T.; El Alam, M.B.; Karpinets, T.V.; Dorta-Estremera, S.; Hegde, V.L.; Nookala, S.; Yoshida-Court, K.; Wu, X.; Biegert, G.W.G.; Delgado Medrano, A.Y.; et al. Gut microbiome diversity is an independent predictor of survival in cervical cancer patients receiving chemoradiation. Commun. Biol. 2021, 4, 237. [Google Scholar] [CrossRef] [PubMed]
- Takasuna, K.; Hagiwara, T.; Hirohashi, M.; Kato, M.; Nomura, M.; Nagai, E.; Yokoi, T.; Kamataki, T. Involvement of beta-glucuronidase in intestinal microflora in the intestinal toxicity of the antitumor camptothecin derivative irinotecan hydrochloride (CPT-11) in rats. Cancer Res. 1996, 56, 3752–3757. [Google Scholar] [PubMed]
- Rong, Y.; Dong, Z.; Hong, Z.; Jin, Y.; Zhang, W.; Zhang, B.; Mao, W.; Kong, H.; Wang, C.; Yang, B.; et al. Reactivity toward Bifidobacterium longum and Enterococcus hirae demonstrate robust CD8+ T cell response and better prognosis in HBV-related hepatocellular carcinoma. Exp. Cell Res. 2017, 358, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Atarashi, K.; Tanoue, T.; Oshima, K.; Suda, W.; Nagano, Y.; Nishikawa, H.; Fukuda, S.; Saito, T.; Narushima, S.; Hase, K.; et al. Treg induction by a rationally selected mixture of Clostridia strains from the human microbiota. Nature 2013, 500, 232–236. [Google Scholar] [CrossRef]
- Deleu, S.; Machiels, K.; Raes, J.; Verbeke, K.; Vermeire, S. Short chain fatty acids and its producing organisms: An overlooked therapy for IBD? EBioMedicine 2021, 66, 103293. [Google Scholar] [CrossRef]
- Gomes, S.D.; Oliveira, C.S.; Azevedo-Silva, J.; Casanova, M.R.; Barreto, J.; Pereira, H.; Chaves, S.R.; Rodrigues, L.R.; Casal, M.; Côrte-Real, M.; et al. The Role of Diet Related Short-Chain Fatty Acids in Colorectal Cancer Metabolism and Survival: Prevention and Therapeutic Implications. Curr. Med. Chem. 2020, 27, 4087–4108. [Google Scholar] [CrossRef]
- Luu, M.; Riester, Z.; Baldrich, A.; Reichardt, N.; Yuille, S.; Busetti, A.; Klein, M.; Wempe, A.; Leister, H.; Raifer, H.; et al. Microbial short-chain fatty acids modulate CD8(+) T cell responses and improve adoptive immunotherapy for cancer. Nat. Commun. 2021, 12, 4077. [Google Scholar] [CrossRef]
- Nomura, M.; Nagatomo, R.; Doi, K.; Shimizu, J.; Baba, K.; Saito, T.; Matsumoto, S.; Inoue, K.; Muto, M. Association of Short-Chain Fatty Acids in the Gut Microbiome With Clinical Response to Treatment With Nivolumab or Pembrolizumab in Patients With Solid Cancer Tumors. JAMA Netw. Open 2020, 3, e202895. [Google Scholar] [CrossRef] [Green Version]
- Coutzac, C.; Jouniaux, J.M.; Paci, A.; Schmidt, J.; Mallardo, D.; Seck, A.; Asvatourian, V.; Cassard, L.; Saulnier, P.; Lacroix, L.; et al. Systemic short chain fatty acids limit antitumor effect of CTLA-4 blockade in hosts with cancer. Nat. Commun. 2020, 11, 2168. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, R.; Afaghi, A.; Babakhani, S.; Sohrabi, M.R.; Hosseini-Fard, S.R.; Babolhavaeji, K.; Khani Ali Akbari, S.; Yousefimashouf, R.; Karampoor, S. Role of microbiota-derived short-chain fatty acids in cancer development and prevention. Biomed. Pharmacother. 2021, 139, 111619. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.A.; Luong, M.K.; Shaw, H.; Nathan, P.; Bataille, V.; Spector, T.D. The gut microbiome: What the oncologist ought to know. Br. J. Cancer 2021, 125, 1197–1209. [Google Scholar] [CrossRef]
- Surana, N.K.; Kasper, D.L. Moving beyond microbiome-wide associations to causal microbe identification. Nature 2017, 552, 244–247. [Google Scholar] [CrossRef] [PubMed]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [Green Version]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.; Macfarlane, G.T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boets, E.; Gomand, S.V.; Deroover, L.; Preston, T.; Vermeulen, K.; De Preter, V.; Hamer, H.M.; Van den Mooter, G.; De Vuyst, L.; Courtin, C.M.; et al. Systemic availability and metabolism of colonic-derived short-chain fatty acids in healthy subjects: A stable isotope study. J. Physiol. 2017, 595, 541–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids From Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNeil, N.I.; Cummings, J.H.; James, W.P. Short chain fatty acid absorption by the human large intestine. Gut 1978, 19, 819–822. [Google Scholar] [CrossRef] [PubMed]
- Trend, S.; Leffler, J.; Jones, A.P.; Cha, L.; Gorman, S.; Brown, D.A.; Breit, S.N.; Kermode, A.G.; French, M.A.; Ward, N.C.; et al. Associations of serum short-chain fatty acids with circulating immune cells and serum biomarkers in patients with multiple sclerosis. Sci. Rep. 2021, 11, 5244. [Google Scholar] [CrossRef]
- Morrison, D.J.; Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pryde, S.E.; Duncan, S.H.; Hold, G.L.; Stewart, C.S.; Flint, H.J. The microbiology of butyrate formation in the human colon. FEMS Microbiol. Lett. 2002, 217, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Louis, P.; Flint, H.J. Formation of propionate and butyrate by the human colonic microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schönfeld, P.; Wojtczak, L. Short- and medium-chain fatty acids in energy metabolism: The cellular perspective. J. Lipid Res. 2016, 57, 943–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bloemen, J.G.; Venema, K.; van de Poll, M.C.; Olde Damink, S.W.; Buurman, W.A.; Dejong, C.H. Short chain fatty acids exchange across the gut and liver in humans measured at surgery. Clin. Nutr. 2009, 28, 657–661. [Google Scholar] [CrossRef] [PubMed]
- Donohoe, D.R.; Garge, N.; Zhang, X.; Sun, W.; O’Connell, T.M.; Bunger, M.K.; Bultman, S.J. The microbiome and butyrate regulate energy metabolism and autophagy in the mammalian colon. Cell Metab. 2011, 13, 517–526. [Google Scholar] [CrossRef] [Green Version]
- Kelly, C.J.; Zheng, L.; Campbell, E.L.; Saeedi, B.; Scholz, C.C.; Bayless, A.J.; Wilson, K.E.; Glover, L.E.; Kominsky, D.J.; Magnuson, A.; et al. Crosstalk between Microbiota-Derived Short-Chain Fatty Acids and Intestinal Epithelial HIF Augments Tissue Barrier Function. Cell Host Microbe 2015, 17, 662–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, M.; Wu, W.; Liu, Z.; Cong, Y. Microbiota metabolite short chain fatty acids, GPCR, and inflammatory bowel diseases. J. Gastroenterol. 2017, 52, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Le Poul, E.; Loison, C.; Struyf, S.; Springael, J.Y.; Lannoy, V.; Decobecq, M.E.; Brezillon, S.; Dupriez, V.; Vassart, G.; Van Damme, J.; et al. Functional characterization of human receptors for short chain fatty acids and their role in polymorphonuclear cell activation. J. Biol. Chem. 2003, 278, 25481–25489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Hee, B.; Wells, J.M. Microbial Regulation of Host Physiology by Short-chain Fatty Acids. Trends Microbiol. 2021, 29, 700–712. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, M.A.; Singh, N.; Martin, P.M.; Thangaraju, M.; Ganapathy, V.; Waller, J.L.; Shi, H.; Robertson, K.D.; Munn, D.H.; Liu, K. Butyrate suppresses colonic inflammation through HDAC1-dependent Fas upregulation and Fas-mediated apoptosis of T cells. Am. J. Physiol.-Gastrointest. Liver Physiol. 2012, 302, G1405–G1415. [Google Scholar] [CrossRef] [PubMed]
- Corrêa-Oliveira, R.; Fachi, J.L.; Vieira, A.; Sato, F.T.; Vinolo, M.A.R. Regulation of immune cell function by short-chain fatty acids. Clin. Transl. Immunol. 2016, 5, e73. [Google Scholar] [CrossRef]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef]
- Park, J.; Kim, M.; Kang, S.G.; Jannasch, A.H.; Cooper, B.; Patterson, J.; Kim, C.H. Short-chain fatty acids induce both effector and regulatory T cells by suppression of histone deacetylases and regulation of the mTOR–S6K pathway. Mucosal Immunol. 2015, 8, 80–93. [Google Scholar] [CrossRef] [Green Version]
- Bachem, A.; Makhlouf, C.; Binger, K.J.; de Souza, D.P.; Tull, D.; Hochheiser, K.; Whitney, P.G.; Fernandez-Ruiz, D.; Dähling, S.; Kastenmüller, W.; et al. Microbiota-Derived Short-Chain Fatty Acids Promote the Memory Potential of Antigen-Activated CD8(+) T Cells. Immunity 2019, 51, 285–297.e5. [Google Scholar] [CrossRef]
- Moffett, J.R.; Puthillathu, N.; Vengilote, R.; Jaworski, D.M.; Namboodiri, A.M. Acetate Revisited: A Key Biomolecule at the Nexus of Metabolism, Epigenetics, and Oncogenesis–Part 2: Acetate and ACSS2 in Health and Disease. Front. Physiol. 2020, 11, 580171. [Google Scholar] [CrossRef]
- Hosseini, E.; Grootaert, C.; Verstraete, W.; Van de Wiele, T. Propionate as a health-promoting microbial metabolite in the human gut. Nutr. Rev. 2011, 69, 245–258. [Google Scholar] [CrossRef]
- Rangan, P.; Mondino, A. Microbial short-chain fatty acids: A strategy to tune adoptive T cell therapy. J. Immunotherapy Cancer 2022, 10, e004147. [Google Scholar] [CrossRef]
- Li, Y.; Seto, E. HDACs and HDAC Inhibitors in Cancer Development and Therapy. Cold Spring Harb. Perspect. Med. 2016, 6, a026831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Dong, B.; Wu, W.; Wang, J.; Jin, H.; Chen, K.; Huang, K.; Huang, S.; Yao, Y. Metagenomic Analyses Reveal Distinct Gut Microbiota Signature for Predicting the Neoadjuvant Chemotherapy Responsiveness in Breast Cancer Patients. Front. Oncol. 2022, 12, 865121. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Fu, L.; Li, Y.; Wang, W.; Gong, M.; Zhang, J.; Dong, X.; Huang, J.; Wang, Q.; Mackay, C.R.; et al. Gut microbial metabolites facilitate anticancer therapy efficacy by modulating cytotoxic CD8(+) T cell immunity. Cell Metab. 2021, 33, 988–1000.e7. [Google Scholar] [CrossRef] [PubMed]
- Geng, H.W.; Yin, F.Y.; Zhang, Z.F.; Gong, X.; Yang, Y. Butyrate Suppresses Glucose Metabolism of Colorectal Cancer Cells via GPR109a-AKT Signaling Pathway and Enhances Chemotherapy. Front. Mol. Biosci. 2021, 8, 634874. [Google Scholar] [CrossRef] [PubMed]
- Encarnação, J.C.; Pires, A.S.; Amaral, R.A.; Gonçalves, T.J.; Laranjo, M.; Casalta-Lopes, J.E.; Gonçalves, A.C.; Sarmento-Ribeiro, A.B.; Abrantes, A.M.; Botelho, M.F. Butyrate, a dietary fiber derivative that improves irinotecan effect in colon cancer cells. J. Nutr. Biochem. 2018, 56, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Panebianco, C.; Villani, A.; Pisati, F.; Orsenigo, F.; Ulaszewska, M.; Latiano, T.P.; Potenza, A.; Andolfo, A.; Terracciano, F.; Tripodo, C.; et al. Butyrate, a postbiotic of intestinal bacteria, affects pancreatic cancer and gemcitabine response in in vitro and in vivo models. Biomed. Pharmacother. 2022, 151, 113163. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; He, P.; Liu, Y.; Qi, M.; Dong, W. Combining Sodium Butyrate With Cisplatin Increases the Apoptosis of Gastric Cancer In Vivo and In Vitro via the Mitochondrial Apoptosis Pathway. Front. Pharmacol. 2021, 12, 708093. [Google Scholar] [CrossRef]
- Kobayashi, M.; Mikami, D.; Uwada, J.; Yazawa, T.; Kamiyama, K.; Kimura, H.; Taniguchi, T.; Iwano, M. A short-chain fatty acid, propionate, enhances the cytotoxic effect of cisplatin by modulating GPR41 signaling pathways in HepG2 cells. Oncotarget 2018, 9, 31342–31354. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.L.; Mao, Y.Q.; Zhang, Z.Y.; Li, Z.M.; Kong, C.Y.; Chen, H.L.; Cai, P.R.; Han, B.; Ye, T.; Wang, L.S. Pectin supplement significantly enhanced the anti-PD-1 efficacy in tumor-bearing mice humanized with gut microbiota from patients with colorectal cancer. Theranostics 2021, 11, 4155–4170. [Google Scholar] [CrossRef]
- Jing, N.; Wang, L.; Zhuang, H.; Jiang, G.; Liu, Z. Ultrafine Jujube Powder Enhances the Infiltration of Immune Cells during Anti-PD-L1 Treatment against Murine Colon Adenocarcinoma. Cancers 2021, 13, 3987. [Google Scholar] [CrossRef]
- Han, K.; Nam, J.; Xu, J.; Sun, X.; Huang, X.; Animasahun, O.; Achreja, A.; Jeon, J.H.; Pursley, B.; Kamada, N.; et al. Generation of systemic antitumour immunity via the in situ modulation of the gut microbiome by an orally administered inulin gel. Nat. Biomed. Eng. 2021, 5, 1377–1388. [Google Scholar] [CrossRef] [PubMed]
- Spencer, C.N.; McQuade, J.L.; Gopalakrishnan, V.; McCulloch, J.A.; Vetizou, M.; Cogdill, A.P.; Khan, M.A.W.; Zhang, X.; White, M.G.; Peterson, C.B.; et al. Dietary fiber and probiotics influence the gut microbiome and melanoma immunotherapy response. Science 2021, 374, 1632–1640. [Google Scholar] [CrossRef] [PubMed]
- Tomita, Y.; Ikeda, T.; Sakata, S.; Saruwatari, K.; Sato, R.; Iyama, S.; Jodai, T.; Akaike, K.; Ishizuka, S.; Saeki, S.; et al. Association of Probiotic Clostridium butyricum Therapy with Survival and Response to Immune Checkpoint Blockade in Patients with Lung Cancer. Cancer Immunol. Res. 2020, 8, 1236–1242. [Google Scholar] [CrossRef] [PubMed]
- Tomita, Y.; Goto, Y.; Sakata, S.; Imamura, K.; Minemura, A.; Oka, K.; Hayashi, A.; Jodai, T.; Akaike, K.; Anai, M.; et al. Clostridium butyricum therapy restores the decreased efficacy of immune checkpoint blockade in lung cancer patients receiving proton pump inhibitors. Oncoimmunology 2022, 11, 2081010. [Google Scholar] [CrossRef] [PubMed]
- Then, C.K.; Paillas, S.; Wang, X.; Hampson, A.; Kiltie, A.E. Association of Bacteroides acidifaciens relative abundance with high-fibre diet-associated radiosensitisation. BMC Biol. 2020, 18, 102. [Google Scholar] [CrossRef]
- Yang, K.; Hou, Y.; Zhang, Y.; Liang, H.; Sharma, A.; Zheng, W.; Wang, L.; Torres, R.; Tatebe, K.; Chmura, S.J.; et al. Suppression of local type I interferon by gut microbiota–derived butyrate impairs antitumor effects of ionizing radiation. J Exp. Med. 2021, 218, e20201915. [Google Scholar] [CrossRef]
- Uribe-Herranz, M.; Rafail, S.; Beghi, S.; Gil-de-Gómez, L.; Verginadis, I.; Bittinger, K.; Pustylnikov, S.; Pierini, S.; Perales-Linares, R.; Blair, I.A.; et al. Gut microbiota modulate dendritic cell antigen presentation and radiotherapy-induced antitumor immune response. J. Clin. Investig. 2020, 130, 466–479. [Google Scholar] [CrossRef] [Green Version]
- Kitazono, M.; Shinchi, H.; Ishigami, S.; Ueno, S.; Natsugoe, S. Effects of a histone deacetylase inhibitor, sodium butyrate, on 53-kDa protein expression and sensitivity to anticancer drugs of pancreatic cancer cells. Curr. Ther. Res. Clin. Exp. 2010, 71, 162–172. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Jiang, W.; Xiao, C.; Yang, W.; Qin, Q.; Mao, A.; Tan, Q.; Lian, B.; Wei, C. Sodium Butyrate Combined with Docetaxel for the Treatment of Lung Adenocarcinoma A549 Cells by Targeting Gli1. OncoTargets Ther. 2020, 13, 8861–8875. [Google Scholar] [CrossRef]
- Frankel, A.E.; Coughlin, L.A.; Kim, J.; Froehlich, T.W.; Xie, Y.; Frenkel, E.P.; Koh, A.Y. Metagenomic Shotgun Sequencing and Unbiased Metabolomic Profiling Identify Specific Human Gut Microbiota and Metabolites Associated with Immune Checkpoint Therapy Efficacy in Melanoma Patients. Neoplasia 2017, 19, 848–855. [Google Scholar] [CrossRef]
- Botticelli, A.; Vernocchi, P.; Marini, F.; Quagliariello, A.; Cerbelli, B.; Reddel, S.; Del Chierico, F.; Di Pietro, F.; Giusti, R.; Tomassini, A.; et al. Gut metabolomics profiling of non-small cell lung cancer (NSCLC) patients under immunotherapy treatment. J. Transl. Med. 2020, 18, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Lou, H.; Peng, Y.; Chen, S.; Fan, L.; Li, X. Elevated levels of circulating short-chain fatty acids and bile acids in type 2 diabetes are linked to gut barrier disruption and disordered gut microbiota. Diabetes Res. Clin. Pract. 2020, 169, 108418. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Alcoholado, L.; Laborda-Illanes, A.; Otero, A.; Ordóñez, R.; González-González, A.; Plaza-Andrades, I.; Ramos-Molina, B.; Gómez-Millán, J.; Queipo-Ortuño, M.I. Relationships of Gut Microbiota Composition, Short-Chain Fatty Acids and Polyamines with the Pathological Response to Neoadjuvant Radiochemotherapy in Colorectal Cancer Patients. Int. J. Mol. Sci. 2021, 22, 9549. [Google Scholar] [CrossRef] [PubMed]
- Yi, Y.; Shen, L.; Shi, W.; Xia, F.; Zhang, H.; Wang, Y.; Zhang, J.; Wang, Y.; Sun, X.; Zhang, Z.; et al. Gut Microbiome Components Predict Response to Neoadjuvant Chemoradiotherapy in Patients with Locally Advanced Rectal Cancer: A Prospective, Longitudinal Study. Clin. Cancer Res. 2021, 27, 1329–1340. [Google Scholar] [CrossRef]
- Sonis, S.T. Regimen-related gastrointestinal toxicities in cancer patients. Curr Opin Support Palliat Care 2010, 4, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Choksey, A.; Timm, K.N. Cancer Therapy-Induced Cardiotoxicity-A Metabolic Perspective on Pathogenesis, Diagnosis and Therapy. Int. J. Mol. Sci. 2021, 23, 441. [Google Scholar] [CrossRef]
- Huang, W.; Guo, H.L.; Deng, X.; Zhu, T.T.; Xiong, J.F.; Xu, Y.H.; Xu, Y. Short-Chain Fatty Acids Inhibit Oxidative Stress and Inflammation in Mesangial Cells Induced by High Glucose and Lipopolysaccharide. Exp. Clin. Endocrinol. Diabetes 2017, 125, 98–105. [Google Scholar] [CrossRef]
- da Silva Ferreira, A.R.; van der Aa, S.A.J.; Wehkamp, T.; Wardill, H.R.; Ten Klooster, J.P.; Garssen, J.; Harthoorn, L.F.; Hartog, A.; Harmsen, H.J.M.; Tissing, W.J.E.; et al. Development of a self-limiting model of methotrexate-induced mucositis reinforces butyrate as a potential therapy. Sci. Rep. 2021, 11, 22911. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, T.M.; Leonel, A.J.; Melo, M.A.; Santos, R.R.; Cara, D.C.; Cardoso, V.N.; Correia, M.I.; Alvarez-Leite, J.I. Oral supplementation of butyrate reduces mucositis and intestinal permeability associated with 5-Fluorouracil administration. Lipids 2012, 47, 669–678. [Google Scholar] [CrossRef] [PubMed]
- Yue, X.; Wen, S.; Long-kun, D.; Man, Y.; Chang, S.; Min, Z.; Shuang-yu, L.; Xin, Q.; Jie, M.; Liang, W. Three important short-chain fatty acids (SCFAs) attenuate the inflammatory response induced by 5-FU and maintain the integrity of intestinal mucosal tight junction. BMC Immunol. 2022, 23, 19. [Google Scholar] [CrossRef]
- Wang, C.; Yang, S.; Gao, L.; Wang, L.; Cao, L. Carboxymethyl pachyman (CMP) reduces intestinal mucositis and regulates the intestinal microflora in 5-fluorouracil-treated CT26 tumour-bearing mice. Food Funct. 2018, 9, 2695–2704. [Google Scholar] [CrossRef]
- Guo, H.; Chou, W.C.; Lai, Y.; Liang, K.; Tam, J.W.; Brickey, W.J.; Chen, L.; Montgomery, N.D.; Li, X.; Bohannon, L.M.; et al. Multi-omics analyses of radiation survivors identify radioprotective microbes and metabolites. Science 2020, 370, eaay9097. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Liu, Y.; Tang, H.; Qiu, M.; Li, C.; Duan, C.; Wang, C.; Yang, J.; Zhou, X. Glabridin Prevents Doxorubicin-Induced Cardiotoxicity Through Gut Microbiota Modulation and Colonic Macrophage Polarization in Mice. Front. Pharmacol. 2019, 10, 107. [Google Scholar] [CrossRef] [PubMed]
- Russo, M.; Guida, F.; Paparo, L.; Trinchese, G.; Aitoro, R.; Avagliano, C.; Fiordelisi, A.; Napolitano, F.; Mercurio, V.; Sala, V.; et al. The novel butyrate derivative phenylalanine-butyramide protects from doxorubicin-induced cardiotoxicity. Eur. J. Heart Fail. 2019, 21, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liu, Y.; Wang, Y.; Chen, X.; Wang, C.; Chen, X.; Yuan, X.; Liu, L.; Yang, J.; Zhou, X. Prevotellaceae produces butyrate to alleviate PD-1/PD-L1 inhibitor-related cardiotoxicity via PPARα-CYP4X1 axis in colonic macrophages. J. Exp. Clin. Cancer Res. 2022, 41, 1. [Google Scholar] [CrossRef] [PubMed]
- Mathewson, N.D.; Jenq, R.; Mathew, A.V.; Koenigsknecht, M.; Hanash, A.; Toubai, T.; Oravecz-Wilson, K.; Wu, S.-R.; Sun, Y.; Rossi, C.; et al. Gut microbiome–derived metabolites modulate intestinal epithelial cell damage and mitigate graft-versus-host disease. Nat. Immunol. 2016, 17, 505–513. [Google Scholar] [CrossRef]
- Cristiano, C.; Cuozzo, M.; Coretti, L.; Liguori, F.M.; Cimmino, F.; Turco, L.; Avagliano, C.; Aviello, G.; Mollica, M.P.; Lembo, F.; et al. Oral sodium butyrate supplementation ameliorates paclitaxel-induced behavioral and intestinal dysfunction. Biomed. Pharmacother. 2022, 153, 113528. [Google Scholar] [CrossRef]
- Rubenstein, E.B.; Peterson, D.E.; Schubert, M.; Keefe, D.; McGuire, D.; Epstein, J.; Elting, L.S.; Fox, P.C.; Cooksley, C.; Sonis, S.T. Clinical practice guidelines for the prevention and treatment of cancer therapy-induced oral and gastrointestinal mucositis. Cancer 2004, 100, 2026–2046. [Google Scholar] [CrossRef] [PubMed]
- Bowen, J.; Al-Dasooqi, N.; Bossi, P.; Wardill, H.; Van Sebille, Y.; Al-Azri, A.; Bateman, E.; Correa, M.E.; Raber-Durlacher, J.; Kandwal, A.; et al. The pathogenesis of mucositis: Updated perspectives and emerging targets. Support Care Cancer 2019, 27, 4023–4033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zhang, Y.; Wei, K.; He, J.; Ding, N.; Hua, J.; Zhou, T.; Niu, F.; Zhou, G.; Shi, T.; et al. Review: Effect of Gut Microbiota and Its Metabolite SCFAs on Radiation-Induced Intestinal Injury. Front. Cell Infect. Microbiol. 2021, 11, 577236. [Google Scholar] [CrossRef]
- Ramos, M.G.; Bambirra, E.A.; Cara, D.C.; Vieira, E.C.; Alvarez-Leite, J.I. Oral administration of short-chain fatty acids reduces the intestinal mucositis caused by treatment with Ara-C in mice fed commercial or elemental diets. Nutr. Cancer 1997, 28, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Gill, P.A.; Bogatyrev, A.; van Zelm, M.C.; Gibson, P.R.; Muir, J.G. Delivery of Acetate to the Peripheral Blood after Consumption of Foods High in Short-Chain Fatty Acids. Mol. Nutr. Food Res. 2021, 65, 2000953. [Google Scholar] [CrossRef]
- Gallotti, B.; Galvao, I.; Leles, G.; Quintanilha, M.F.; Souza, R.O.; Miranda, V.C.; Rocha, V.M.; Trindade, L.M.; Jesus, L.C.L.; Mendes, V.; et al. Effects of dietary fibre intake in chemotherapy-induced mucositis in murine model. Br. J. Nutr. 2021, 126, 853–864. [Google Scholar] [CrossRef]
- Tian, T.; Zhao, Y.; Yang, Y.; Wang, T.; Jin, S.; Guo, J.; Liu, Z. The protective role of short-chain fatty acids acting as signal molecules in chemotherapy- or radiation-induced intestinal inflammation. Am. J. Cancer Res. 2020, 10, 3508–3531. [Google Scholar] [PubMed]
- Ferreira, M.R.; Sands, C.J.; Li, J.V.; Andreyev, J.N.; Chekmeneva, E.; Gulliford, S.; Marchesi, J.; Lewis, M.R.; Dearnaley, D.P. Impact of Pelvic Radiation Therapy for Prostate Cancer on Global Metabolic Profiles and Microbiota-Driven Gastrointestinal Late Side Effects: A Longitudinal Observational Study. Int. J. Radiat. Oncol. Biol. Phys. 2021, 111, 1204–1213. [Google Scholar] [CrossRef] [PubMed]
- Tabaja, L.; Sidani, S.M. Management of Radiation Proctitis. Dig. Dis. Sci. 2018, 63, 2180–2188. [Google Scholar] [CrossRef] [PubMed]
- Vernia, P.; Fracasso, P.L.; Casale, V.; Villotti, G.; Marcheggiano, A.; Stigliano, V.; Pinnaro, P.; Bagnardi, V.; Caprilli, R. Topical butyrate for acute radiation proctitis: Randomised, crossover trial. Lancet 2000, 356, 1232–1235. [Google Scholar] [CrossRef]
- Hille, A.; Herrmann, M.K.; Kertesz, T.; Christiansen, H.; Hermann, R.M.; Pradier, O.; Schmidberger, H.; Hess, C.F. Sodium butyrate enemas in the treatment of acute radiation-induced proctitis in patients with prostate cancer and the impact on late proctitis. A prospective evaluation. Strahlenther. Onkol. 2008, 184, 686–692. [Google Scholar] [CrossRef] [PubMed]
- Maggio, A.; Magli, A.; Rancati, T.; Fiorino, C.; Valvo, F.; Fellin, G.; Ricardi, U.; Munoz, F.; Cosentino, D.; Cazzaniga, L.F.; et al. Daily sodium butyrate enema for the prevention of radiation proctitis in prostate cancer patients undergoing radical radiation therapy: Results of a multicenter randomized placebo-controlled dose-finding phase 2 study. Int. J. Radiat. Oncol. Biol. Phys. 2014, 89, 518–524. [Google Scholar] [CrossRef] [PubMed]
- Bowen, J.M.; Gibson, R.J.; Coller, J.K.; Blijlevens, N.; Bossi, P.; Al-Dasooqi, N.; Bateman, E.H.; Chiang, K.; de Mooij, C.; Mayo, B.; et al. Systematic review of agents for the management of cancer treatment-related gastrointestinal mucositis and clinical practice guidelines. Support Care Cancer 2019, 27, 4011–4022. [Google Scholar] [CrossRef]
- Sasidharan, B.K.; Ramadass, B.; Viswanathan, P.N.; Samuel, P.; Gowri, M.; Pugazhendhi, S.; Ramakrishna, B.S. A phase 2 randomized controlled trial of oral resistant starch supplements in the prevention of acute radiation proctitis in patients treated for cervical cancer. J. Cancer Res. Ther. 2019, 15, 1383–1391. [Google Scholar] [CrossRef]
- Jain, D.; Aronow, W. Cardiotoxicity of cancer chemotherapy in clinical practice. Hosp. Pract. (1995) 2019, 47, 6–15. [Google Scholar] [CrossRef]
- Florescu, M.; Cinteza, M.; Vinereanu, D. Chemotherapy-induced Cardiotoxicity. Maedica 2013, 8, 59–67. [Google Scholar] [PubMed]
- Rahouma, M.; Karim, N.A.; Baudo, M.; Yahia, M.; Kamel, M.; Eldessouki, I.; Abouarab, A.; Saad, I.; Elmously, A.; Gray, K.D.; et al. Cardiotoxicity with immune system targeting drugs: A meta-analysis of anti-PD/PD-L1 immunotherapy randomized clinical trials. Immunotherapy 2019, 11, 725–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ichikawa, Y.; Ghanefar, M.; Bayeva, M.; Wu, R.; Khechaduri, A.; Naga Prasad, S.V.; Mutharasan, R.K.; Naik, T.J.; Ardehali, H. Cardiotoxicity of doxorubicin is mediated through mitochondrial iron accumulation. J. Clin. Investig. 2014, 124, 617–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simůnek, T.; Stérba, M.; Popelová, O.; Adamcová, M.; Hrdina, R.; Gersl, V. Anthracycline-induced cardiotoxicity: Overview of studies examining the roles of oxidative stress and free cellular iron. Pharmacol. Rep. 2009, 61, 154–171. [Google Scholar] [CrossRef]
- Tay, W.T.; Fang, Y.H.; Beh, S.T.; Liu, Y.W.; Hsu, L.W.; Yen, C.J.; Liu, P.Y. Programmed Cell Death-1: Programmed Cell Death-Ligand 1 Interaction Protects Human Cardiomyocytes Against T-Cell Mediated Inflammation and Apoptosis Response In Vitro. Int. J. Mol. Sci. 2020, 21, 2399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitzthum, L.K.; Heide, E.S.; Park, H.; Williamson, C.W.; Sheridan, P.; Huynh-Le, M.P.; Sirak, I.; Wei, L.; Tarnawski, R.; Mahantshetty, U.; et al. Comparison of Hematologic Toxicity and Bone Marrow Compensatory Response in Head and Neck vs Cervical Cancer Patients Undergoing Chemoradiotherapy. Front. Oncol. 2020, 10, 1179. [Google Scholar] [PubMed]
- Pearlman, R.; Hanna, R.; Burmeister, J.; Abrams, J.; Dominello, M. Adverse Effects of Total Body Irradiation: A Two-Decade, Single Institution Analysis. Adv. Radiat. Oncol. 2021, 6, 100723. [Google Scholar] [CrossRef] [PubMed]
- Thiruvengadam, M.; Subramanian, U.; Venkidasamy, B.; Thirupathi, P.; Samynathan, R.; Shariati, M.A.; Rebezov, M.; Chung, I.M.; Rengasamy, K.R.R. Emerging role of nutritional short-chain fatty acids (SCFAs) against cancer via modulation of hematopoiesis. Crit. Rev. Food. Sci. Nutr. 2021, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Motoori, M.; Sugimura, K.; Tanaka, K.; Shiraishi, O.; Kimura, Y.; Miyata, H.; Yamasaki, M.; Makino, T.; Miyazaki, Y.; Iwama, M.; et al. Comparison of synbiotics combined with enteral nutrition and prophylactic antibiotics as supportive care in patients with esophageal cancer undergoing neoadjuvant chemotherapy: A multicenter randomized study. Clin. Nutr. 2022, 41, 1112–1121. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, V.; Kolli, S.S.; Strowd, L.C. Review of Graft-Versus-Host Disease. Dermatol Clin 2019, 37, 569–582. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, J.L.; Levine, J.E.; Reddy, P.; Holler, E. Graft-versus-host disease. Lancet 2009, 373, 1550–1561. [Google Scholar] [CrossRef]
- Payen, M.; Nicolis, I.; Robin, M.; Michonneau, D.; Delannoye, J.; Mayeur, C.; Kapel, N.; Berçot, B.; Butel, M.J.; Le Goff, J.; et al. Functional and phylogenetic alterations in gut microbiome are linked to graft-versus-host disease severity. Blood Adv. 2020, 4, 1824–1832. [Google Scholar] [CrossRef]
- Biagi, E.; Zama, D.; Nastasi, C.; Consolandi, C.; Fiori, J.; Rampelli, S.; Turroni, S.; Centanni, M.; Severgnini, M.; Peano, C.; et al. Gut microbiota trajectory in pediatric patients undergoing hematopoietic SCT. Bone Marrow Transplant. 2015, 50, 992–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romick-Rosendale, L.E.; Haslam, D.B.; Lane, A.; Denson, L.; Lake, K.; Wilkey, A.; Watanabe, M.; Bauer, S.; Litts, B.; Luebbering, N.; et al. Antibiotic Exposure and Reduced Short Chain Fatty Acid Production after Hematopoietic Stem Cell Transplant. Biol. Blood Marrow Transplant. 2018, 24, 2418–2424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markey, K.A.; Schluter, J.; Gomes, A.L.C.; Littmann, E.R.; Pickard, A.J.; Taylor, B.P.; Giardina, P.A.; Weber, D.; Dai, A.; Docampo, M.D.; et al. The microbe-derived short-chain fatty acids butyrate and propionate are associated with protection from chronic GVHD. Blood 2020, 136, 130–136. [Google Scholar] [CrossRef]
- Galloway-Peña, J.R.; Peterson, C.B.; Malik, F.; Sahasrabhojane, P.V.; Shah, D.P.; Brumlow, C.E.; Carlin, L.G.; Chemaly, R.F.; Im, J.S.; Rondon, G.; et al. Fecal Microbiome, Metabolites, and Stem Cell Transplant Outcomes: A Single-Center Pilot Study. Open Forum Infect. Dis. 2019, 6, ofz173. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara, H.; Docampo, M.D.; Riwes, M.; Peltier, D.; Toubai, T.; Henig, I.; Wu, S.J.; Kim, S.; Taylor, A.; Brabbs, S.; et al. Microbial metabolite sensor GPR43 controls severity of experimental GVHD. Nat. Commun. 2018, 9, 3674. [Google Scholar] [CrossRef] [Green Version]
- Apple, A.C.; Schroeder, M.P.; Ryals, A.J.; Wagner, L.I.; Cella, D.; Shih, P.A.; Reilly, J.; Penedo, F.J.; Voss, J.L.; Wang, L. Hippocampal functional connectivity is related to self-reported cognitive concerns in breast cancer patients undergoing adjuvant therapy. Neuroimage Clin. 2018, 20, 110–118. [Google Scholar] [CrossRef]
- Apple, A.C.; Ryals, A.J.; Alpert, K.I.; Wagner, L.I.; Shih, P.A.; Dokucu, M.; Cella, D.; Penedo, F.J.; Voss, J.L.; Wang, L. Subtle hippocampal deformities in breast cancer survivors with reduced episodic memory and self-reported cognitive concerns. Neuroimage Clin. 2017, 14, 685–691. [Google Scholar] [CrossRef] [PubMed]
- Kesler, S.R.; Kent, J.S.; O’Hara, R. Prefrontal cortex and executive function impairments in primary breast cancer. Arch. Neurol. 2011, 68, 1447–1453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, R.; Dos Santos, M.R.; das Chagas Valota, I.A.; Sousa, C.S.; Costa Calache, A.L.S. Factors associated with sleep quality during chemotherapy: An integrative review. Nurs. Open 2020, 7, 1274–1284. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wen, Y.; Bedi, C.; Humphris, G. The relationship between cancer patient’s fear of recurrence and chemotherapy: A systematic review and meta-analysis. J. Psychosom. Res. 2017, 98, 55–63. [Google Scholar] [CrossRef] [Green Version]
- Okubo, R.; Kinoshita, T.; Katsumata, N.; Uezono, Y.; Xiao, J.; Matsuoka, Y.J. Impact of chemotherapy on the association between fear of cancer recurrence and the gut microbiota in breast cancer survivors. Brain Behav. Immun. 2020, 85, 186–191. [Google Scholar] [CrossRef]
- Paulsen, J.A.; Ptacek, T.S.; Carter, S.J.; Liu, N.; Kumar, R.; Hyndman, L.; Lefkowitz, E.J.; Morrow, C.D.; Rogers, L.Q. Gut microbiota composition associated with alterations in cardiorespiratory fitness and psychosocial outcomes among breast cancer survivors. Support Care Cancer 2017, 25, 1563–1570. [Google Scholar] [CrossRef] [Green Version]
- Subramaniam, C.B.; Bowen, J.M.; Gladman, M.A.; Lustberg, M.B.; Mayo, S.J.; Wardill, H.R. The microbiota-gut-brain axis: An emerging therapeutic target in chemotherapy-induced cognitive impairment. Neurosci. Biobehav. Rev. 2020, 116, 470–479. [Google Scholar] [CrossRef]
- Santos, J.C.; Pyter, L.M. Neuroimmunology of Behavioral Comorbidities Associated With Cancer and Cancer Treatments. Front. Immunol. 2018, 9, 1195. [Google Scholar] [CrossRef] [Green Version]
- Müller, B.; Rasmusson, A.J.; Just, D.; Jayarathna, S.; Moazzami, A.; Novicic, Z.K.; Cunningham, J.L. Fecal Short-Chain Fatty Acid Ratios as Related to Gastrointestinal and Depressive Symptoms in Young Adults. Psychosom. Med. 2021, 83, 693–699. [Google Scholar] [CrossRef]
- Wu, L.; Han, Y.; Zheng, Z.; Peng, G.; Liu, P.; Yue, S.; Zhu, S.; Chen, J.; Lv, H.; Shao, L.; et al. Altered Gut Microbial Metabolites in Amnestic Mild Cognitive Impairment and Alzheimer’s Disease: Signals in Host-Microbe Interplay. Nutrients 2021, 13, 228. [Google Scholar] [CrossRef]
- Lee, H.J.; Son, Y.; Lee, M.; Moon, C.; Kim, S.H.; Shin, I.S.; Yang, M.; Bae, S.; Kim, J.S. Sodium butyrate prevents radiation-induced cognitive impairment by restoring pCREB/BDNF expression. Neural. Regen. Res. 2019, 14, 1530–1535. [Google Scholar] [PubMed]
- Topuz, R.D.; Gunduz, O.; Tastekin, E.; Karadag, C.H. Effects of hippocampal histone acetylation and HDAC inhibition on spatial learning and memory in the Morris water maze in rats. Fundam. Clin. Pharmacol. 2020, 34, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Ji, M.; Dong, L.; Jia, M.; Liu, W.; Zhang, M.; Ju, L.; Yang, J.; Xie, Z.; Yang, J. Epigenetic enhancement of brain-derived neurotrophic factor signaling pathway improves cognitive impairments induced by isoflurane exposure in aged rats. Mol. Neurobiol. 2014, 50, 937–944. [Google Scholar] [CrossRef]
- Fernando, W.; Martins, I.J.; Morici, M.; Bharadwaj, P.; Rainey-Smith, S.R.; Lim, W.L.F.; Martins, R.N. Sodium Butyrate Reduces Brain Amyloid-β Levels and Improves Cognitive Memory Performance in an Alzheimer’s Disease Transgenic Mouse Model at an Early Disease Stage. J. Alzheimers Dis. 2020, 74, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Unger, M.M.; Spiegel, J.; Dillmann, K.U.; Grundmann, D.; Philippeit, H.; Bürmann, J.; Faßbender, K.; Schwiertz, A.; Schäfer, K.H. Short chain fatty acids and gut microbiota differ between patients with Parkinson’s disease and age-matched controls. Parkinsonism Relat. Disord. 2016, 32, 66–72. [Google Scholar] [CrossRef]
- Dizman, N.; Meza, L.; Bergerot, P.; Alcantara, M.; Dorff, T.; Lyou, Y.; Frankel, P.; Cui, Y.; Mira, V.; Llamas, M.; et al. Nivolumab plus ipilimumab with or without live bacterial supplementation in metastatic renal cell carcinoma: A randomized phase 1 trial. Nat. Med. 2022, 28, 704–712. [Google Scholar] [CrossRef]
- Mattace Raso, G.; Simeoli, R.; Russo, R.; Iacono, A.; Santoro, A.; Paciello, O.; Ferrante, M.C.; Canani, R.B.; Calignano, A.; Meli, R. Effects of sodium butyrate and its synthetic amide derivative on liver inflammation and glucose tolerance in an animal model of steatosis induced by high fat diet. PLoS ONE 2013, 8, e68626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wardill, H.R.; Secombe, K.R.; Bryant, R.V.; Hazenberg, M.D.; Costello, S.P. Adjunctive fecal microbiota transplantation in supportive oncology: Emerging indications and considerations in immunocompromised patients. EBioMedicine 2019, 44, 730–740. [Google Scholar] [CrossRef] [Green Version]
- Lu, K.; Dong, S.; Wu, X.; Jin, R.; Chen, H. Probiotics in Cancer. Front. Oncol. 2021, 11, 638148. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Anselmo, A.C.; Huang, L. Nanotechnology intervention of the microbiome for cancer therapy. Nat. Nanotechnol. 2019, 14, 1093–1103. [Google Scholar] [CrossRef]
- Singh, N.; Gurav, A.; Sivaprakasam, S.; Brady, E.; Padia, R.; Shi, H.; Thangaraju, M.; Prasad, P.D.; Manicassamy, S.; Munn, D.H.; et al. Activation of Gpr109a, receptor for niacin and the commensal metabolite butyrate, suppresses colonic inflammation and carcinogenesis. Immunity 2014, 40, 128–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wardill, H.R.; Chan, R.J.; Chan, A.; Keefe, D.; Costello, S.P.; Hart, N.H. Dual contribution of the gut microbiome to immunotherapy efficacy and toxicity: Supportive care implications and recommendations. Support Care Cancer 2022, 30, 6369–6373. [Google Scholar] [CrossRef] [PubMed]
- Hersi, F.; Elgendy, S.M.; Al Shamma, S.A.; Altell, R.T.; Sadiek, O.; Omar, H.A. Cancer immunotherapy resistance: The impact of microbiome-derived short-chain fatty acids and other emerging metabolites. Life Sci. 2022, 300, 120573. [Google Scholar] [CrossRef] [PubMed]
- Luu, M.; Visekruna, A. Microbial metabolites: Novel therapeutic tools for boosting cancer therapies. Trends Cell Biol. 2021, 31, 873–875. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wang, B.; Dong, J.; Li, Y.; Zhang, S.; Zeng, X.; Xiao, H.; Fan, S.; Cui, M. Gut Microbiota-Derived l-Histidine/Imidazole Propionate Axis Fights against the Radiation-Induced Cardiopulmonary Injury. Int. J. Mol. Sci. 2021, 22, 11436. [Google Scholar] [CrossRef] [PubMed]
- Gill, P.A.; van Zelm, M.C.; Muir, J.G.; Gibson, P.R. Review article: Short chain fatty acids as potential therapeutic agents in human gastrointestinal and inflammatory disorders. Aliment. Pharmacol. Ther. 2018, 48, 15–34. [Google Scholar] [CrossRef] [Green Version]
- Rahat-Rozenbloom, S.; Fernandes, J.; Cheng, J.; Gloor, G.B.; Wolever, T.M. The acute effects of inulin and resistant starch on postprandial serum short-chain fatty acids and second-meal glycemic response in lean and overweight humans. Eur. J. Clin. Nutr. 2017, 71, 227–233. [Google Scholar] [CrossRef] [Green Version]
- Hamaker, B.R.; Tuncil, Y.E. A perspective on the complexity of dietary fiber structures and their potential effect on the gut microbiota. J. Mol. Biol. 2014, 426, 3838–3850. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Qadami, G.H.; Secombe, K.R.; Subramaniam, C.B.; Wardill, H.R.; Bowen, J.M. Gut Microbiota-Derived Short-Chain Fatty Acids: Impact on Cancer Treatment Response and Toxicities. Microorganisms 2022, 10, 2048. https://doi.org/10.3390/microorganisms10102048
Al-Qadami GH, Secombe KR, Subramaniam CB, Wardill HR, Bowen JM. Gut Microbiota-Derived Short-Chain Fatty Acids: Impact on Cancer Treatment Response and Toxicities. Microorganisms. 2022; 10(10):2048. https://doi.org/10.3390/microorganisms10102048
Chicago/Turabian StyleAl-Qadami, Ghanyah H., Kate R. Secombe, Courtney B. Subramaniam, Hannah R. Wardill, and Joanne M. Bowen. 2022. "Gut Microbiota-Derived Short-Chain Fatty Acids: Impact on Cancer Treatment Response and Toxicities" Microorganisms 10, no. 10: 2048. https://doi.org/10.3390/microorganisms10102048