
Enhanced Hemolytic Activity of Mesophilic Aeromonas salmonicida SRW-OG1 Is Brought about by Elevated Temperatures

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Transcriptome Sequencing
2.3. Read Processing and Mapping
2.4. Functional Classification and Enrichment Analysis
2.5. Quantitative Real-Time PCR (qPCR)
2.6. Sequence Alignment Analysis, Evolutionary Tree Construction, and Protein Structure Prediction
2.7. Knockdown of aerA and hlyA in A. salmonicida
2.8. Determination of Hemolytic Ability
2.9. Extraction of ECPs at Different Culture Temperatures
2.10. Artificial Infection and Sampling
2.11. Statistical Analysis
3. Results
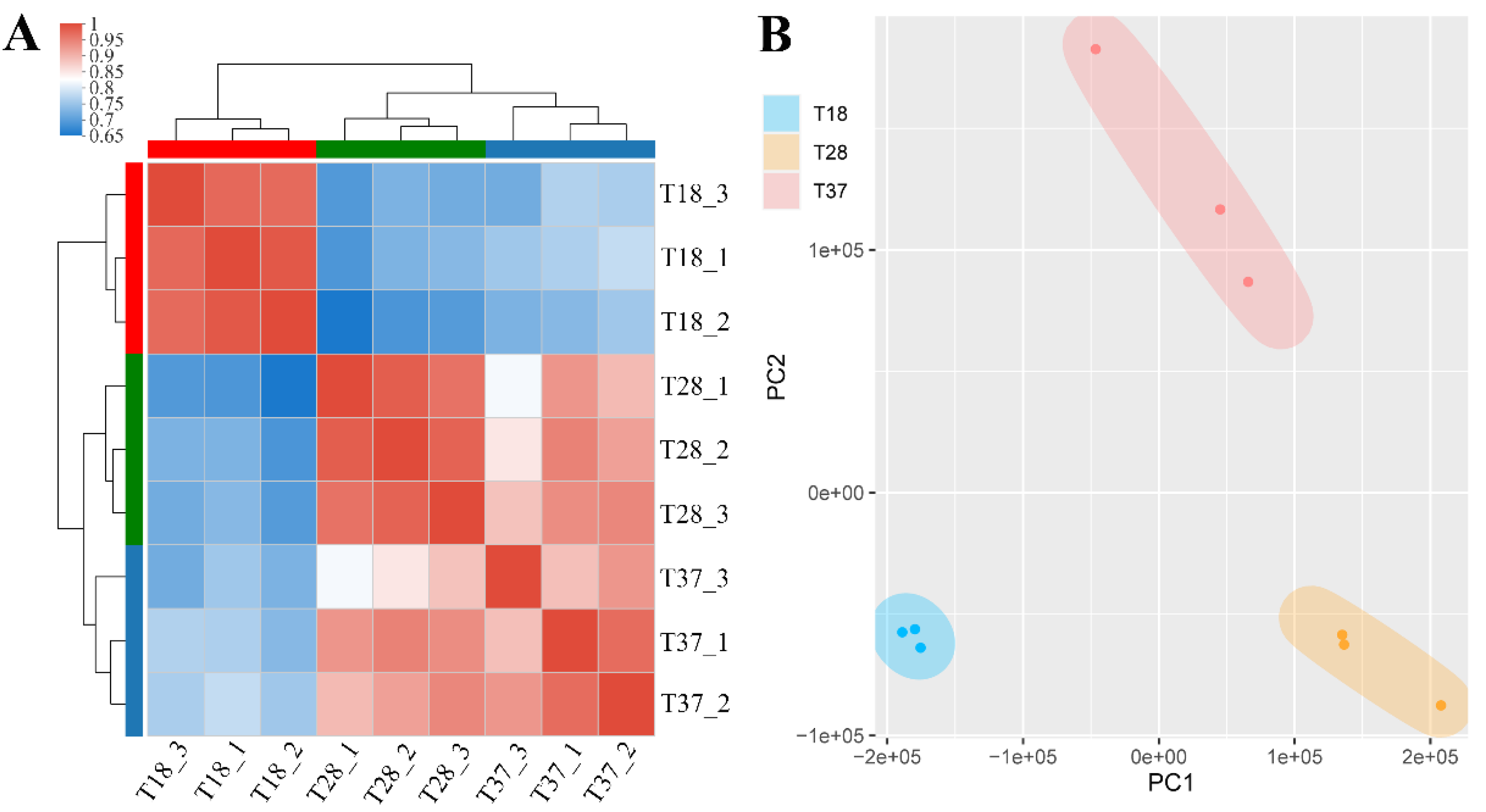
3.1. RNA and Data Quality
3.2. Functional Annotation of Genes Identified in All Samples
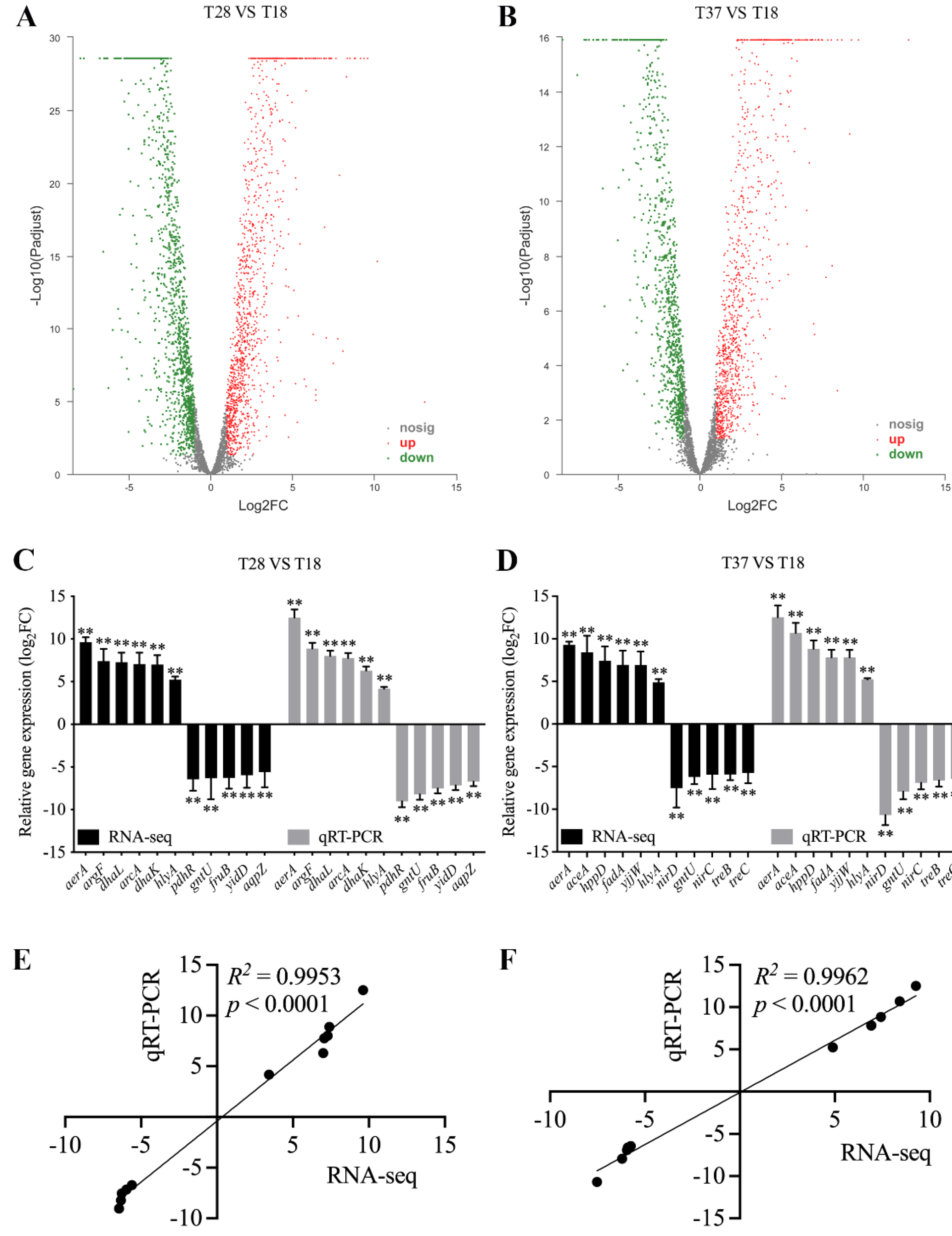
3.3. Validation of RNA-seq
3.4. Analysis of Differentially Expressed Genes
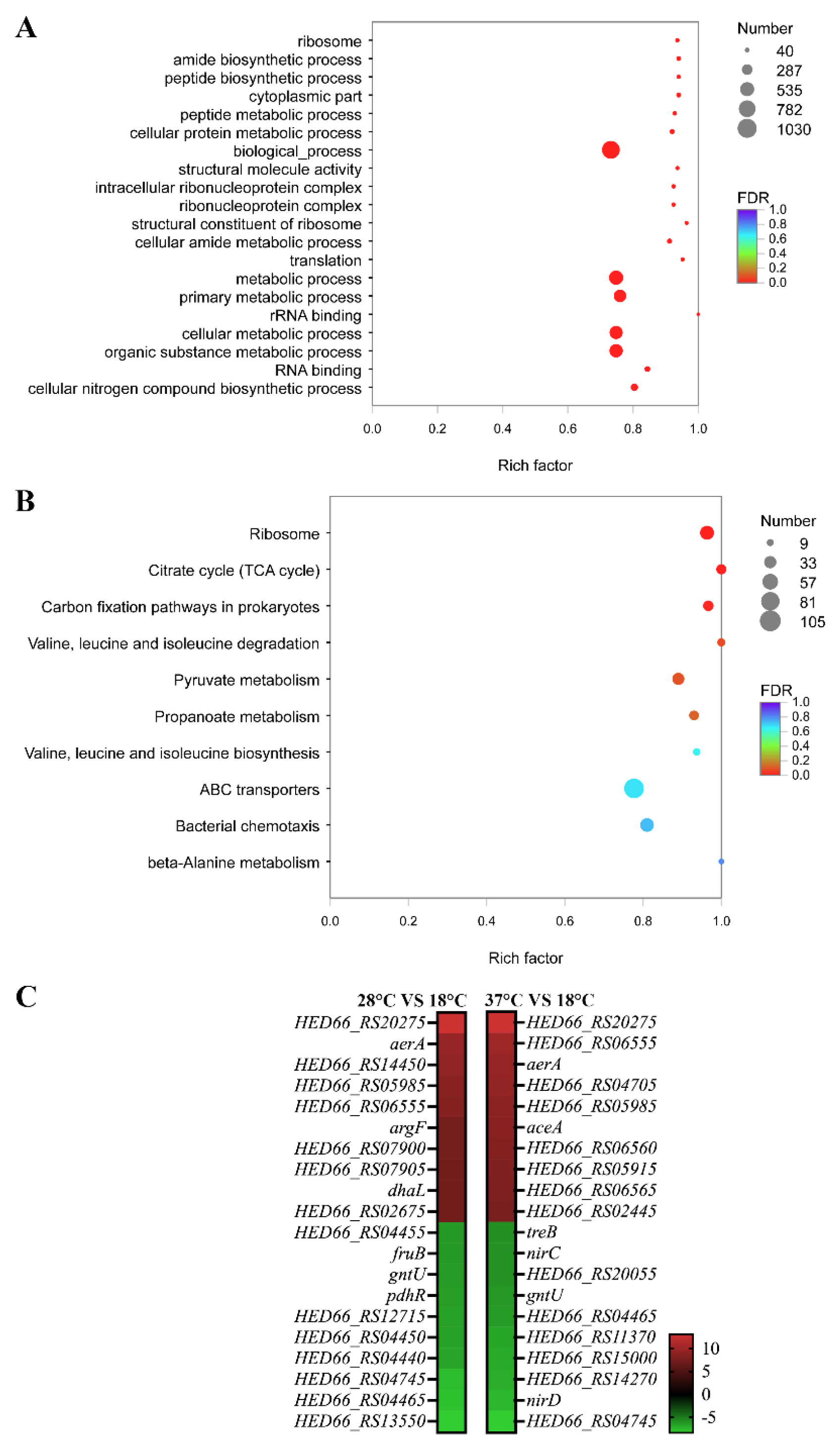
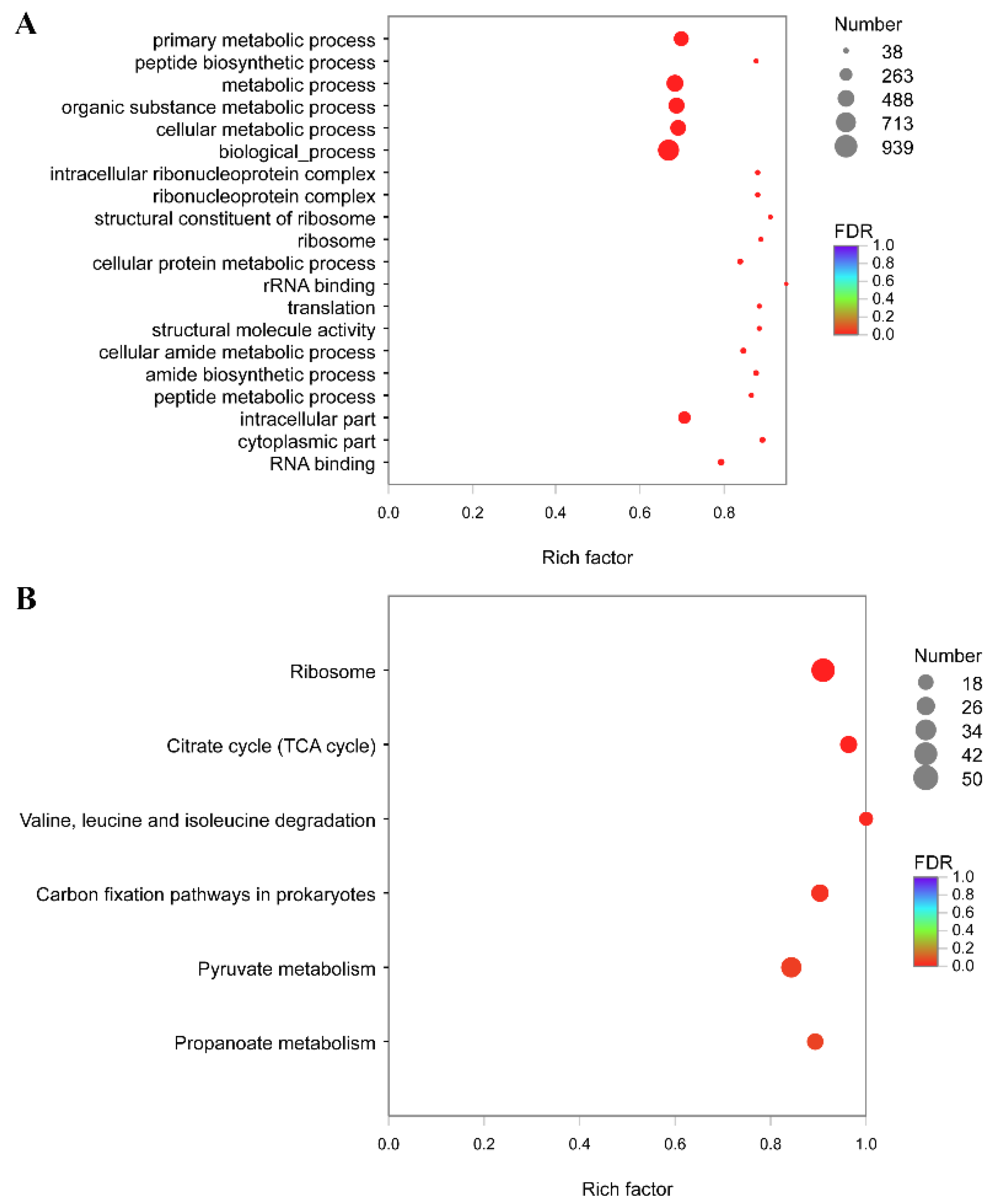
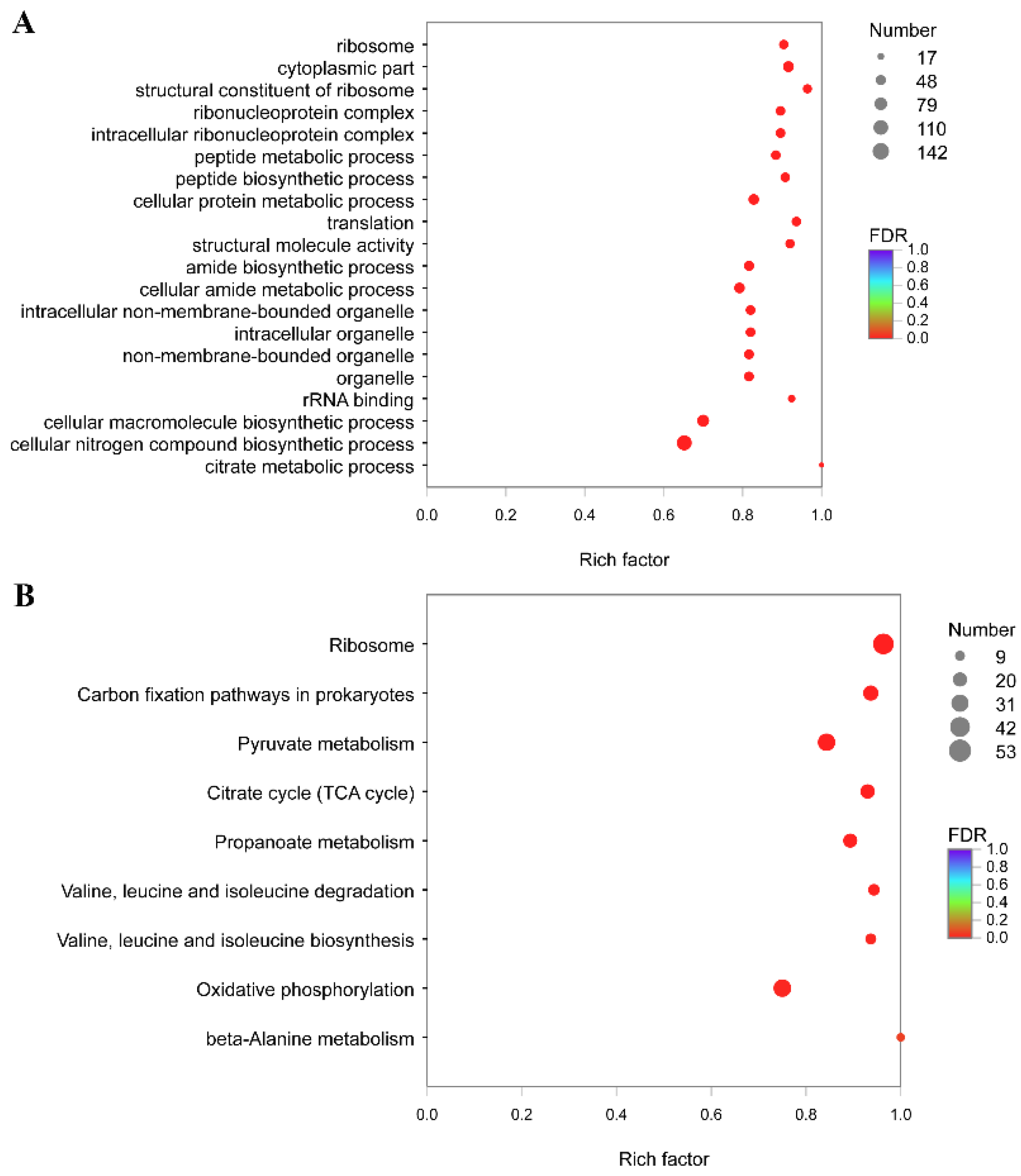
3.5. Functional Annotation and Enrichment Analysis of Genes Specifically Down-Regulated under 28 °C
3.6. Functional Annotation and Enrichment Analysis of Genes Specifically Up-Regulated under 28 °C
3.7. Functional Annotation and Enrichment Analysis of Genes Specifically Down-Regulated under 37 °C
3.8. Functional Annotation and Enrichment Analysis of Genes Specifically Up-Regulated under 37 °C
3.9. The Sequence Alignment Analysis of aerA and hlyA of A. salmonicida
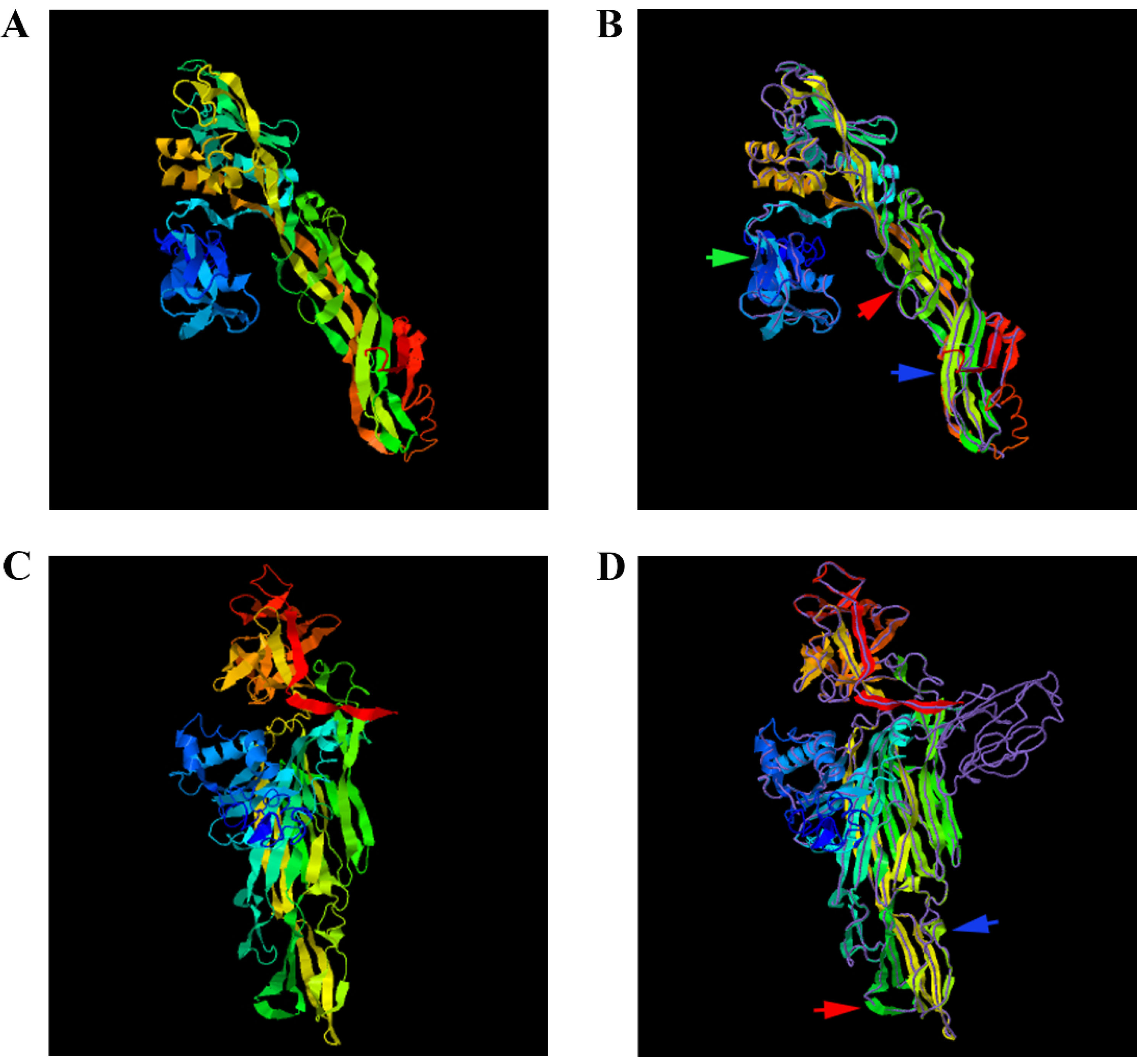
3.10. Protein Structure Prediction of Aerolysin and Hemolysin of A. salmonicida
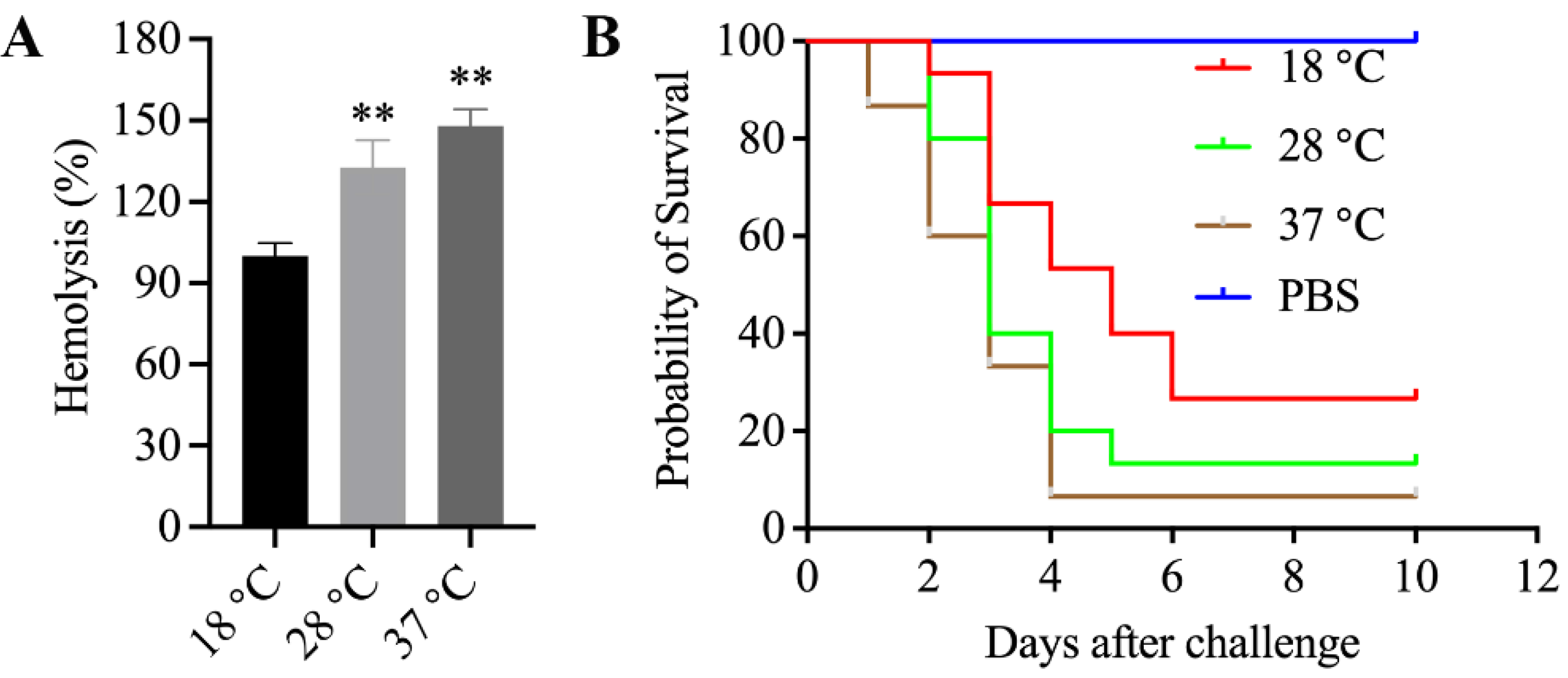
3.11. Enhanced Hemolytic Activity of Mesophilic A. salmonicida SRW-OG1 Was Induced by Elevated Temperature
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Findlay, C.; Tatner, M.F.J.F. The effect of reconstitution with cryopreserved thymocytes on thein vivoantibody response to sheep red blood cells, Aeromonas salmonicida, and DNP-KLH, in adult long term thymectomised rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 1996, 6, 371–381. [Google Scholar] [CrossRef]
- Hoover, G.J.; El-Mowafi, A. Plasma proteins of rainbow trout (Oncorhynchus mykiss) isolated by binding to lipopolysaccharide from Aeromonas salmonicida. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1998, 120, 559–569. [Google Scholar] [CrossRef]
- Long, M.; Zhao, J. Transcriptomic and proteomic analyses of splenic immune mechanisms of rainbow trout (Oncorhynchus mykiss) infected by Aeromonas salmonicida subsp. salmonicida. J. Proteom. 2015, 122, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Connors, E.; Soto-Dávila, M. Identification and validation of reliable Aeromonas salmonicida subspecies salmonicida reference genes for differential gene expression analyses. Infect. Genet. Evol. 2019, 73, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Coscelli, G.A.; Bermúdez, R. Vaccination against Aeromonas salmonicida in turbot (Scophthalmus maximus L.): Study of the efficacy, morphological changes and antigen distribution. Aquaculture 2015, 445, 22–32. [Google Scholar] [CrossRef]
- Nakayama, K.; Yamashita, R. Use of common carp (Cyprinus carpio) and Aeromonas salmonicida for detection of immunomodulatory effects of chemicals on fish. Mar. Pollut. Bull. 2017, 124, 710–713. [Google Scholar] [CrossRef] [PubMed]
- Braden, L.M.; Whyte, S.K. Vaccine-induced protection against furunculosis involves pre-emptive priming of humoral immunity in Arctic charr. Front. Immunol. 2019, 10, 120. [Google Scholar] [CrossRef] [PubMed]
- Dallaire-Dufresne, S.; Tanaka, K.H. Virulence, genomic features, and plasticity of Aeromonas salmonicida subsp. salmonicida, the causative agent of fish furunculosis. Vet. Microbiol. 2014, 169, 1–7. [Google Scholar]
- Huang, L.; Qi, W. The immune response of a warm water fish orange-spotted grouper (Epinephelus coioides) infected with a typical cold water bacterial pathogen Aeromonas salmonicida is AhR dependent. Dev. Comp. Immunol. 2020, 113, 103779. [Google Scholar] [CrossRef]
- Pavan, M.; Abbott, S. Aeromonas salmonicida subsp. pectinolytica subsp. nov., a new pectinase-positive subspecies isolated from a heavily polluted river. Int. J. Syst. Evol. Microbiol. 2000, 50, 1119–1124. [Google Scholar]
- Charette, S.J.J.M. Microbe profile: Aeromonas salmonicida: An opportunistic pathogen with multiple personalities. Microbiology 2021, 167, 001052. [Google Scholar] [CrossRef]
- Leduc, G.R.; Paquet, V.E. Characterization of bacteriophage T7-Ah reveals its lytic activity against a subset of both mesophilic and psychrophilic Aeromonas salmonicida strains. Arch. Virol. 2021, 166, 521–533. [Google Scholar] [CrossRef]
- Lee, H.J.; Hoel, S. Aeromonas spp. isolated from ready-to-eat seafood on the Norwegian market: Prevalence, putative virulence factors and antimicrobial resistance. J. Appl. Microbiol. 2021, 130, 1380–1393. [Google Scholar] [CrossRef]
- Umutoni, N.; Jakobsen, A.N. Occurrence, diversity and temperature-dependent growth kinetics of Aeromonas spp. in lettuce. Int. J. Food Microbiol. 2020, 335, 108852. [Google Scholar] [CrossRef]
- Vincent, A.T.; Bernatchez, A. A mesophilic Aeromonas salmonicida strain isolated from an unsuspected host, the migratory bird pied avocet. Microorganisms 2019, 7, 592. [Google Scholar] [CrossRef] [Green Version]
- Vincent, A.T.; Rouleau, F.D. Study of mesophilic Aeromonas salmonicida A527 strain sheds light on the species’ lifestyles and taxonomic dilemma. FEMS Microbiol. Lett. 2017, 364, fnx239. [Google Scholar] [CrossRef] [Green Version]
- Vincent, A.T.; Trudel, M.V. Increasing genomic diversity and evidence of constrained lifestyle evolution due to insertion sequences in Aeromonas salmonicida. BMC Genom. 2016, 17, 44. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Y.; Qi, W. Insights into mesophilic virulence, antibiotic resistant and human pathogenicity: A genomics study on the Aeromonas salmonicida SRW-OG1 newly isolated from the Asian fish Epinephelus coioides. Aquaculture 2021, 539, 736630. [Google Scholar] [CrossRef]
- Bi, Z.; Liu, Y. Contribution of AhyR to virulence of Aeromonas hydrophila J-1. Res. Vet. Sci. 2007, 83, 150–156. [Google Scholar] [CrossRef]
- Abrami, L.; Fivaz, M. Sensitivity of polarized epithelial cells to the pore-forming toxin aerolysin. Infect. Immun. 2003, 71, 739–746. [Google Scholar] [CrossRef] [Green Version]
- Iacovache, I.; Paumard, P. A rivet model for channel formation by aerolysin-like pore-forming toxins. EMBO J. 2006, 25, 457–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, J.; Liu, Y. Morin protects channel catfish from Aeromonas hydrophila infection by blocking aerolysin activity. Front. Microbiol. 2018, 9, 2828. [Google Scholar] [CrossRef] [PubMed]
- Heuzenroeder, M.W.; Wong, C.Y. Distribution of two hemolytic toxin genes in clinical and environmental isolates of Aeromonas spp.: Correlation with virulence in a suckling mouse model. FEMS Microbiol. Lett. 1999, 174, 131–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.; Qin, Y. MinD plays an important role in Aeromonas hydrophila adherence to Anguilla japonica mucus. Gene 2015, 565, 275–281. [Google Scholar] [CrossRef]
- Xu, W.; Lin, W. Disentangling the abundance and structure of Vibrio communities in a semi-enclosed Bay with mariculture (Dongshan Bay, Southern China). Comput. Struct. Biotechnol. J. 2021, 19, 4381–4393. [Google Scholar] [CrossRef]
- Huang, L.; Zuo, Y. A metabolomic investigation into the temperature-dependent virulence of Pseudomonas plecoglossicida from large yellow croaker (Pseudosciaena crocea). J. Fish Dis. 2019, 42, 431–446. [Google Scholar] [CrossRef]
- Zuo, Y.; Zhao, L. Mechanisms underlying the virulence regulation of new Vibrio alginolyticus ncRNA Vvrr1 with a comparative proteomic analysis. Emerg. Microbes Infect. 2019, 8, 1604–1618. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Xin, G. Novel insights into host-pathogen interactions of large yellow croakers (Larimichthys crocea) and pathogenic bacterium Pseudomonas plecoglossicida using time-resolved dual RNA-seq of infected spleens. Zool. Res. 2020, 41, 314. [Google Scholar] [CrossRef]
- Li, R.; Yu, C. SOAP2: An improved ultrafast tool for short read alignment. Bioinformatics 2009, 25, 1966–1967. [Google Scholar] [CrossRef] [Green Version]
- Mortazavi, A.; Williams, B.A. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef]
- Huang, L.; Zuo, Y. The Zinc Nutritional Immunity of Epinephelus coioides Contributes to the Importance of znuC during Pseudomonas plecoglossicida Infection. Front. Immunol. 2021, 1575. [Google Scholar] [CrossRef]
- Choi, K.-H.; Schweizer, H.P.J.N.P. mini-Tn7 insertion in bacteria with single attTn7 sites: Example Pseudomonas Aeruginosa. Nat. Protoc. 2006, 1, 153–161. [Google Scholar] [CrossRef]
- Darsigny, M.; Babeu, J.-P. Hepatocyte nuclear factor-4α promotes gut neoplasia in mice and protects against the production of reactive oxygen species. Cancer Res. 2010, 70, 9423–9433. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Qiao, Y. Full-Length Transcriptome: A Reliable Alternative for Single-Cell RNA-Seq Analysis in the Spleen of Teleost without Reference Genome. Front. Immunol. 2021, 3974. [Google Scholar] [CrossRef]
- Sahu, I.; Das, B. Toxicity of crude extracellular products of Aeromonas hydrophila on Rohu, Labeo rohita (Ham.). Indian J. Microbiol. 2011, 51, 515–520. [Google Scholar] [CrossRef] [Green Version]
- He, R.; Wang, J. Effect of Ferredoxin Receptor FusA on the Virulence Mechanism of Pseudomonas plecoglossicida. Front. Cell. Infect. Microbiol. 2022, 255. [Google Scholar] [CrossRef]
- Abrami, L.; Fivaz, M. Adventures of a pore-forming toxin at the target cell surface. Trends Microbiol. 2000, 8, 168–172. [Google Scholar] [CrossRef]
- Galinier, R.; Portela, J. Biomphalysin, a new β pore-forming toxin involved in Biomphalaria glabrata immune defense against Schistosoma mansoni. PLoS Pathog. 2013, 9, e1003216. [Google Scholar] [CrossRef]
- Olson, R.; Gouaux, E.J.J.O.M.B. Crystal structure of the Vibrio cholerae cytolysin (VCC) pro-toxin and its assembly into a heptameric transmembrane pore. J. Mol. Biol. 2005, 350, 997–1016. [Google Scholar] [CrossRef]
- Walker, B.; Bayley, H.J.J.O.B.C. Key Residues for Membrane Binding, Oligomerization, and Pore Forming Activity of Staphylococcal α-Hemolysin Identified by Cysteine Scanning Mutagenesis and Targeted Chemical Modification (∗). J. Biol. Chem. 1995, 270, 23065–23071. [Google Scholar] [CrossRef] [Green Version]
- Kang, Y.-R.; Kim, A. Complete Genome Sequence of Aeromonas salmonicida subsp. masoucida Strain BR19001YR, Isolated from Diseased Korean Rockfish (Sebastes schlegelii). Microbiol. Resour. Announc. 2021, 10, e01281-20. [Google Scholar] [PubMed]
- Varshney, A.; Das, M. Aeromonas salmonicida as a causative agent for postoperative endophthalmitis. Middle East Afr. J. Ophthalmol. 2017, 24, 213. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.T.; Fernández-Bravo, A. Investigation of the virulence and genomics of Aeromonas salmonicida strains isolated from human patients. Infect. Genet. Evol. 2019, 68, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.T.; Intertaglia, L. AsaGEI2d: A new variant of a genomic island identified in a group of Aeromonas salmonicida subsp. salmonicida isolated from France, which bears the pAsa7 plasmid. FEMS Microbiol. Lett. 2021, 368, fnab021. [Google Scholar]
- Huang, L.; Liu, W. Integration of transcriptomic and proteomic approaches reveals the temperature-dependent virulence of Pseudomonas plecoglossicida. Front. Cell. Infect. Microbiol. 2018, 8, 207. [Google Scholar] [CrossRef] [Green Version]
- Rezaei, M.N.; Aslankoohi, E. Contribution of the tricarboxylic acid (TCA) cycle and the glyoxylate shunt in Saccharomyces cerevisiae to succinic acid production during dough fermentation. Int. J. Food Microbiol. 2015, 204, 24–32. [Google Scholar] [CrossRef]
- Massilamany, C.; Gangaplara, A. TCA cycle inactivation in Staphylococcus aureus alters nitric oxide production in RAW 264.7 cells. Mol. Cell. Biochem. 2011, 355, 75–82. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Huang, L. The TCA pathway is an important player in the regulatory network governing Vibrio alginolyticus adhesion under adversity. Front. Microbiol. 2016, 7, 40. [Google Scholar] [CrossRef] [Green Version]
- Troxell, B.; Zhang, J.-J. Pyruvate protects pathogenic spirochetes from H2O2 killing. PLoS ONE 2014, 9, e84625. [Google Scholar] [CrossRef] [Green Version]
- SchäR, J.; Stoll, R. Pyruvate carboxylase plays a crucial role in carbon metabolism of extra-and intracellularly replicating Listeria monocytogenes. J. Bacteriol. 2010, 192, 1774–1784. [Google Scholar] [CrossRef] [Green Version]
- Xie, T.; Pang, R. Cold tolerance regulated by the pyruvate metabolism in Vibrio parahaemolyticus. Front. Microbiol. 2019, 10, 178. [Google Scholar] [CrossRef] [Green Version]
- Bücker, R.; Heroven, A.K. The pyruvate-tricarboxylic acid cycle node: A focal point of virulence control in the enteric pathogen Yersinia pseudotuberculosis. J. Biol. Chem. 2014, 289, 30114–30132. [Google Scholar] [CrossRef] [Green Version]
- Harper, L.; Balasubramanian, D. Staphylococcus aureus responds to the central metabolite pyruvate to regulate virulence. MBio 2018, 9, e02272-17. [Google Scholar] [CrossRef]
- Steiner, B.D.; Eberly, A.R. Evidence of cross-regulation in two closely related pyruvate-sensing systems in uropathogenic Escherichia coli. J. Membr. Biol. 2018, 251, 65–74. [Google Scholar] [CrossRef]
- Kaiser, J.C.; Sen, S. The role of two branched-chain amino acid transporters in Staphylococcus aureus growth, membrane fatty acid composition and virulence. Mol. Microbiol. 2016, 102, 850–864. [Google Scholar] [CrossRef] [Green Version]
- Orasch, T.; Dietl, A.-M. The leucine biosynthetic pathway is crucial for adaptation to iron starvation and virulence in Aspergillus fumigatus. Virulence 2019, 10, 925–934. [Google Scholar] [CrossRef] [Green Version]
- Dutta, S.; Corsi, I.D. BrnQ-Type Branched-Chain Amino Acid Transporters Influence Bacillus anthracis Growth and Virulence. Mbio 2022, 13, e03640-21. [Google Scholar] [CrossRef]
- Tsuda, K.; Nagano, H. Modulation of fatty acid composition and growth in Sporosarcina species in response to temperatures and exogenous branched-chain amino acids. Appl. Microbiol. Biotechnol. 2017, 101, 5071–5080. [Google Scholar] [CrossRef]
- Zhu, K.; Ding, X. Exogenous isoleucine and fatty acid shortening ensure the high content of anteiso-C15:0 fatty acid required for low-temperature growth of Listeria monocytogenes. Appl. Environ. Microbiol. 2005, 71, 8002–8007. [Google Scholar] [CrossRef] [Green Version]
- Andreini, C.; Bertini, I. Metal ions in biological catalysis: From enzyme databases to general principles. JBIC J. Biol. Inorg. Chem. 2008, 13, 1205–1218. [Google Scholar] [CrossRef]
- Besold, A.N.; Gilston, B.A. Role of calprotectin in withholding zinc and copper from Candida albicans. Infect. Immun. 2018, 86, e00779-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassat, J.E.; Skaar, E.P.J.C.H. Iron in infection and immunity. Cell Host Microbe 2013, 13, 509–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hood, M.I.; Skaar, E.P.J.N.R.M. Nutritional immunity: Transition metals at the pathogen–host interface. Nat. Rev. Microbiol. 2012, 10, 525–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kehl-Fie, T.E.; Skaar, E.P. Nutritional immunity beyond iron: A role for manganese and zinc. Curr. Opin. Chem. Biol. 2010, 14, 218–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hantke, K.J.C.O.I.M. Bacterial zinc uptake and regulators. Curr. Opin. Microbiol. 2005, 8, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Andreini, C.; Banci, L. Zinc through the three domains of life. J. Proteome Res. 2006, 5, 3173–3178. [Google Scholar] [CrossRef] [PubMed]
- Corbin, B.D.; Seeley, E.H. Metal chelation and inhibition of bacterial growth in tissue abscesses. Science 2008, 319, 962–965. [Google Scholar] [CrossRef] [PubMed]
- Kehl-Fie, T.E.; Zhang, Y. MntABC and MntH contribute to systemic Staphylococcus aureus infection by competing with calprotectin for nutrient manganese. Infect. Immun. 2013, 81, 3395–3405. [Google Scholar] [CrossRef] [Green Version]
- Klein, J.S.; Lewinson, O.J.M. Bacterial ATP-driven transporters of transition metals: Physiological roles, mechanisms of action, and roles in bacterial virulence. Metallomics 2011, 3, 1098–1108. [Google Scholar] [CrossRef]
- D’orazio, M.; Mastropasqua, M.C. The capability of Pseudomonas aeruginosa to recruit zinc under conditions of limited metal availability is affected by inactivation of the ZnuABC transporter. Metallomics 2015, 7, 1023–1035. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.Z.; Jellbauer, S. Zinc sequestration by the neutrophil protein calprotectin enhances Salmonella growth in the inflamed gut. Cell Host Microbe 2012, 11, 227–239. [Google Scholar] [CrossRef] [Green Version]
- Qi, W.; Gao, Q. Immune responses and inorganic ion transport regulations of Epinephelus coioides in response to L321_RS13075 gene of Pseudomonas plecoglossicida. Fish Shellfish Immunol. 2022, 120, 599–609. [Google Scholar] [CrossRef]
- Monnet, V.J.C.; Cmls, M.L.S. Bacterial oligopeptide-binding proteins. Cell. Mol. Life Sci. CMLS 2003, 60, 2100–2114. [Google Scholar] [CrossRef]
- Wang, C.-H.; Lin, C.-Y. Effects of oligopeptide permease in group A Streptococcal infection. Infect. Immun. 2005, 73, 2881–2890. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.-K.; Wang, Y.-K. Identification of a Vibrio furnissii oligopeptide permease and characterization of its in vitro hemolytic activity. J. Bacteriol. 2007, 189, 8215–8223. [Google Scholar] [CrossRef] [Green Version]
- Samen, U.; Gottschalk, B. Relevance of peptide uptake systems to the physiology and virulence of Streptococcus agalactiae. J. Bacteriol. 2004, 186, 1398–1408. [Google Scholar] [CrossRef] [Green Version]
- Slamti, L.; Lereclus, D.J.T.E.J. A cell–cell signaling peptide activates the PlcR virulence regulon in bacteria of the Bacillus cereus group. EMBO J. 2002, 21, 4550–4559. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Huang, L. Contributions of the oligopeptide permeases in multistep of Vibrio alginolyticus pathogenesis. MicrobiologyOpen 2017, 6, e00511. [Google Scholar] [CrossRef]
- Sit, B.; Crowley, S.M. Active transport of phosphorylated carbohydrates promotes intestinal colonization and transmission of a bacterial pathogen. PLoS Pathog. 2015, 11, e1005107. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Wang, L. mcp, aer, cheB, and cheV contribute to the regulation of Vibrio alginolyticus (ND-01) adhesion under gradients of environmental factors. MicrobiologyOpen 2017, 6, e00517. [Google Scholar] [CrossRef]
- Freter, R.; O’brien, P. Secretory Immunity and Infection. In Adhesion and Chemotaxis as Determinants of Bacterial Association with Mucosal Surfaces; Springer: Berlin/Heidelberg, Germany, 1978; pp. 429–437. [Google Scholar]
- Bassler, B.; Gibbons, P. Chitin utilization by marine bacteria. Chemotaxis to chitin oligosaccharides by Vibrio furnissii. J. Biol. Chem. 1991, 266, 24268–24275. [Google Scholar] [CrossRef]
- Gupta, P.; Thomas, S.E. A fragment-based approach to assess the ligandability of ArgB, ArgC, ArgD and ArgF in the L-arginine biosynthetic pathway of Mycobacterium tuberculosis. Comput. Struct. Biotechnol. J. 2021, 19, 3491–3506. [Google Scholar] [CrossRef] [PubMed]
- Yassin, A.F.; Langenberg, S. Draft genome sequence of Actinotignum schaalii DSM 15541T: Genetic insights into the lifestyle, cell fitness and virulence. PLoS ONE 2017, 12, e0188914. [Google Scholar] [CrossRef] [PubMed]
- Abdelhamed, H.; Ramachandran, R. Contributions of a LysR transcriptional regulator to Listeria monocytogenes virulence and identification of its regulons. J. Bacteriol. 2020, 202, e00087-20. [Google Scholar] [CrossRef]
- Yang, K.; Shadkchan, Y. Contribution of ATPase copper transporters in animal but not plant virulence of the crossover pathogen Aspergillus flavus. Virulence 2018, 9, 1273–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.; Zhao, L. Dual RNA-Seq unveils Pseudomonas plecoglossicida htpG gene functions during host-pathogen interactions with Epinephelus coioides. Front. Immunol. 2019, 10, 984. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Wang, J.; Cai, H.; Lin, M.; Zhang, Y.; Huang, L. Enhanced Hemolytic Activity of Mesophilic Aeromonas salmonicida SRW-OG1 Is Brought about by Elevated Temperatures. Microorganisms 2022, 10, 2033. https://doi.org/10.3390/microorganisms10102033
Chen Y, Wang J, Cai H, Lin M, Zhang Y, Huang L. Enhanced Hemolytic Activity of Mesophilic Aeromonas salmonicida SRW-OG1 Is Brought about by Elevated Temperatures. Microorganisms. 2022; 10(10):2033. https://doi.org/10.3390/microorganisms10102033
Chicago/Turabian StyleChen, Yunong, Jiajia Wang, Hongyan Cai, Mao Lin, Youyu Zhang, and Lixing Huang. 2022. "Enhanced Hemolytic Activity of Mesophilic Aeromonas salmonicida SRW-OG1 Is Brought about by Elevated Temperatures" Microorganisms 10, no. 10: 2033. https://doi.org/10.3390/microorganisms10102033