
Extracellular c-di-GMP Plays a Role in Biofilm Formation and Dispersion of Campylobacter jejuni

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. C. jejuni Strains and Growth Conditions
2.2. Chemicals Used in This Study
2.3. Biofilm Formation and Dispersion Assays
2.4. Swarm Assay
2.5. Effect of Extracellular c-di-GMP on C. jejuni Growth
2.6. Auto-Aggregation Assay
2.7. Adherence Assay with Cultured Caco-2 Cells In Vitro
2.8. RT-PCR Analysis
2.9. Confocal Laser Scanning Microscopy
2.10. Statistical Analysis
3. Results
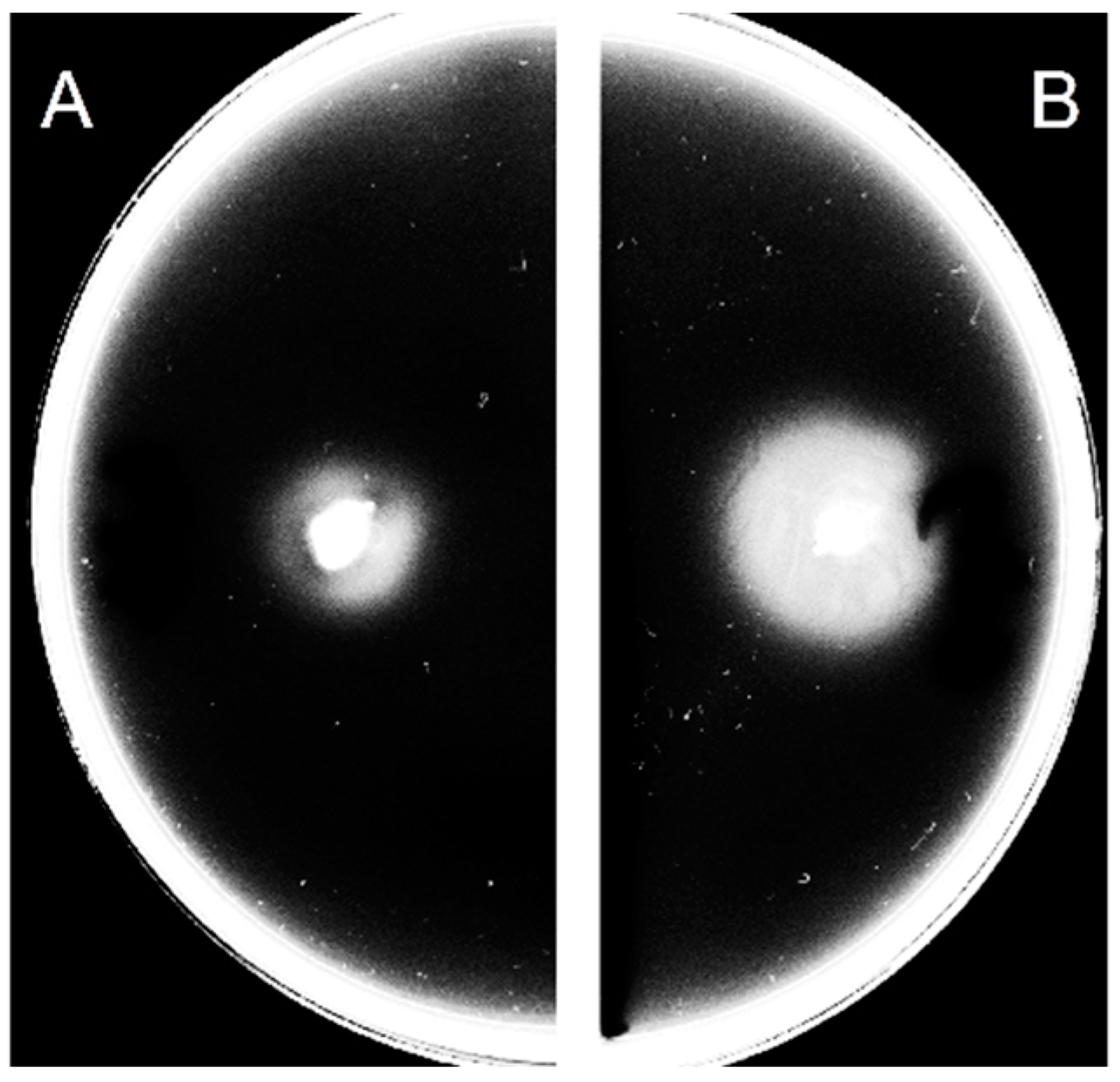
3.1. Extracellular c-di-GMP Affects the Motility of C. jejuni
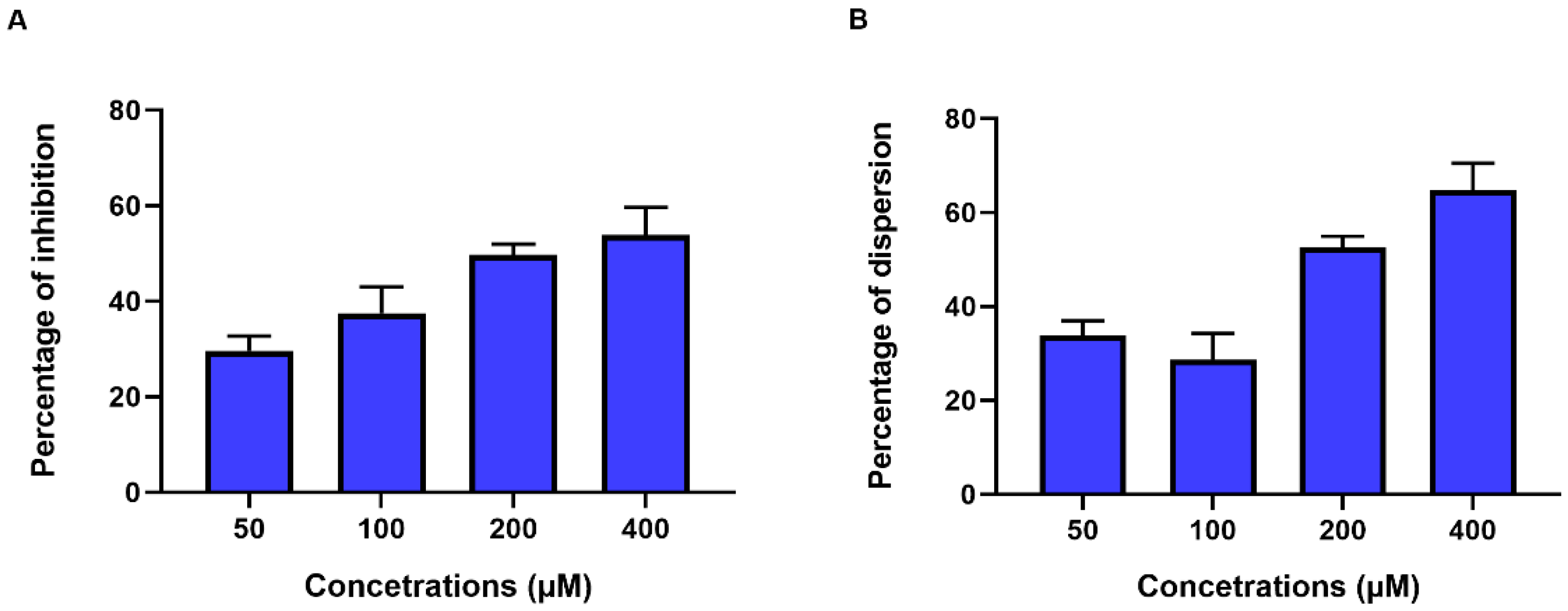
3.2. Extracellular c-di-GMP Affects Biofilm Formation by C. jejuni
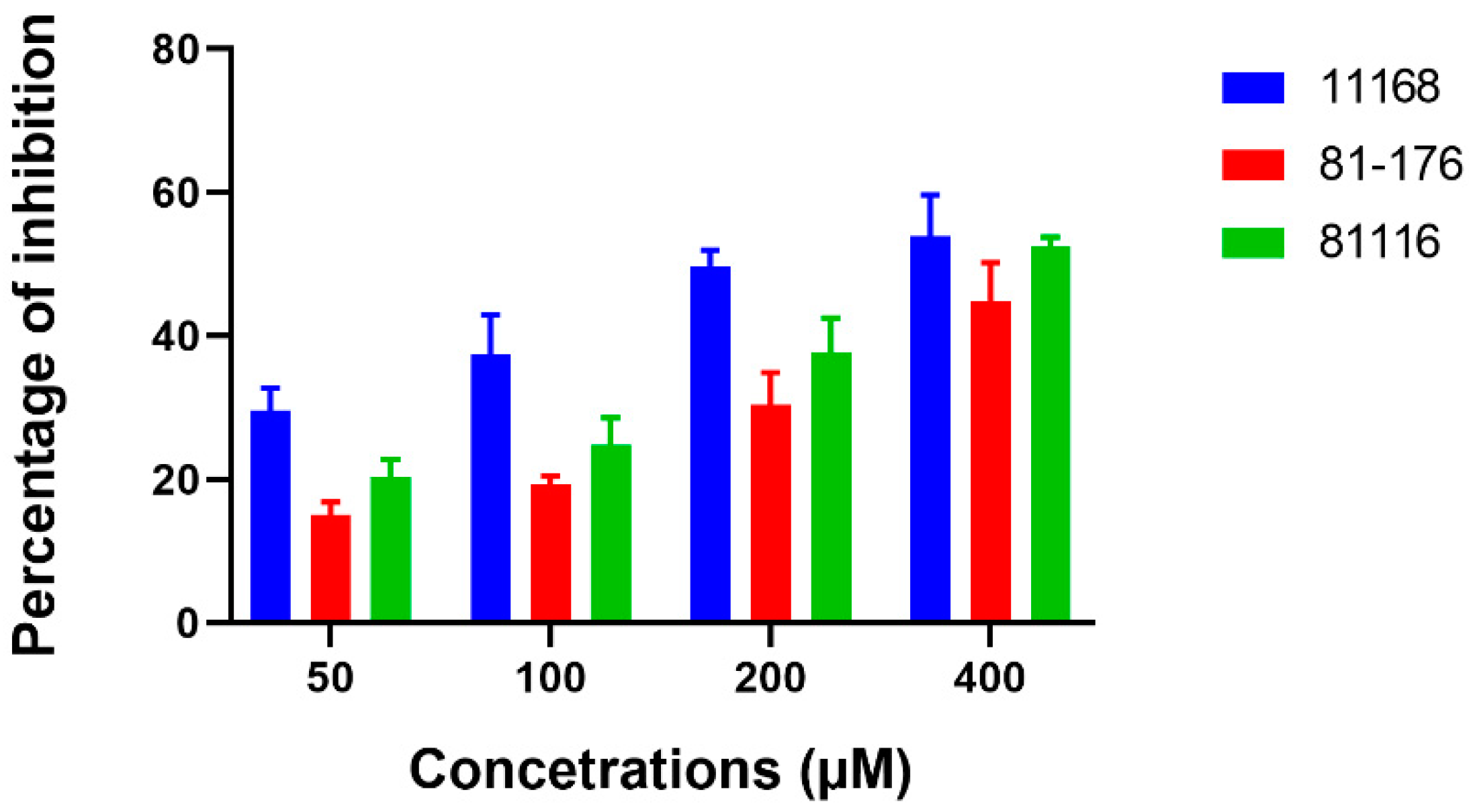
3.3. Extracellular c-di-GMP Affects Biofilm Formation by Different Campylobacter Strains
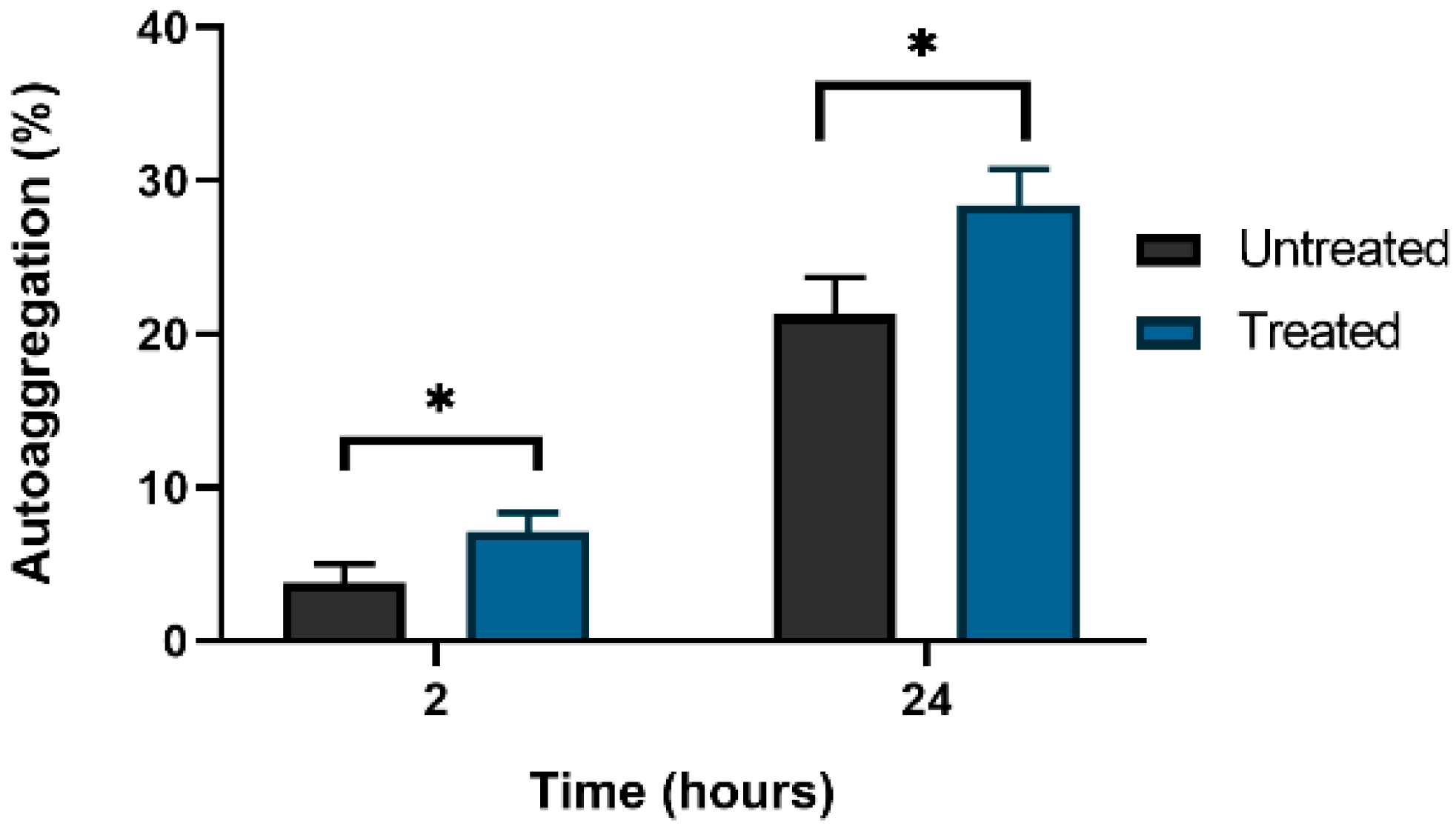
3.4. Extracellular c-di-GMP Treatment Effects Auto-Aggregation of C. jejuni
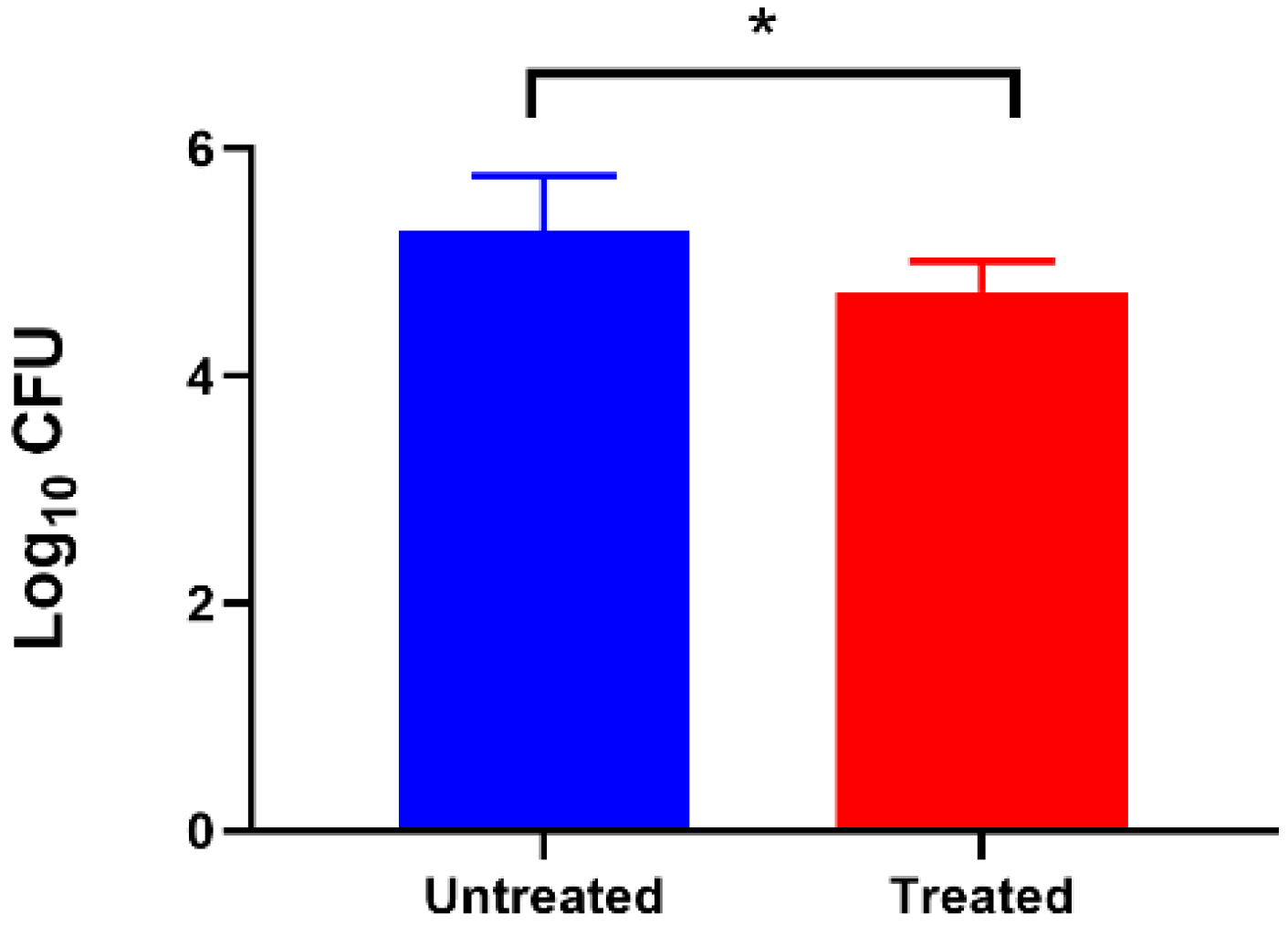
3.5. Effect of Extracellular c-di-GMP on Adhesion of C. jejuni to Caco-2 Cell Lines
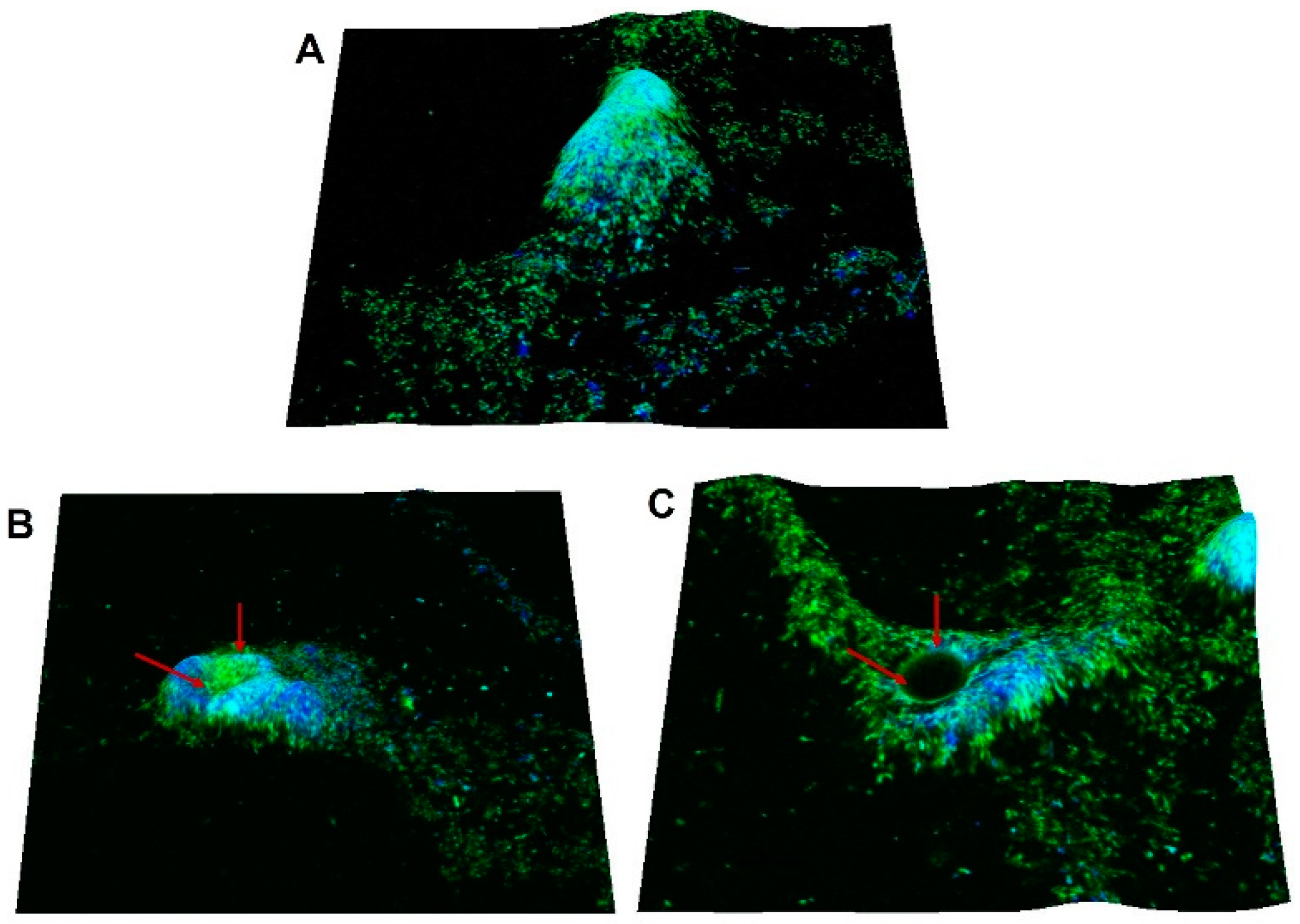
3.6. Dispersal Effect of Extracellular c-di-GMP on C. jejuni Biofilm Can Be Visualized
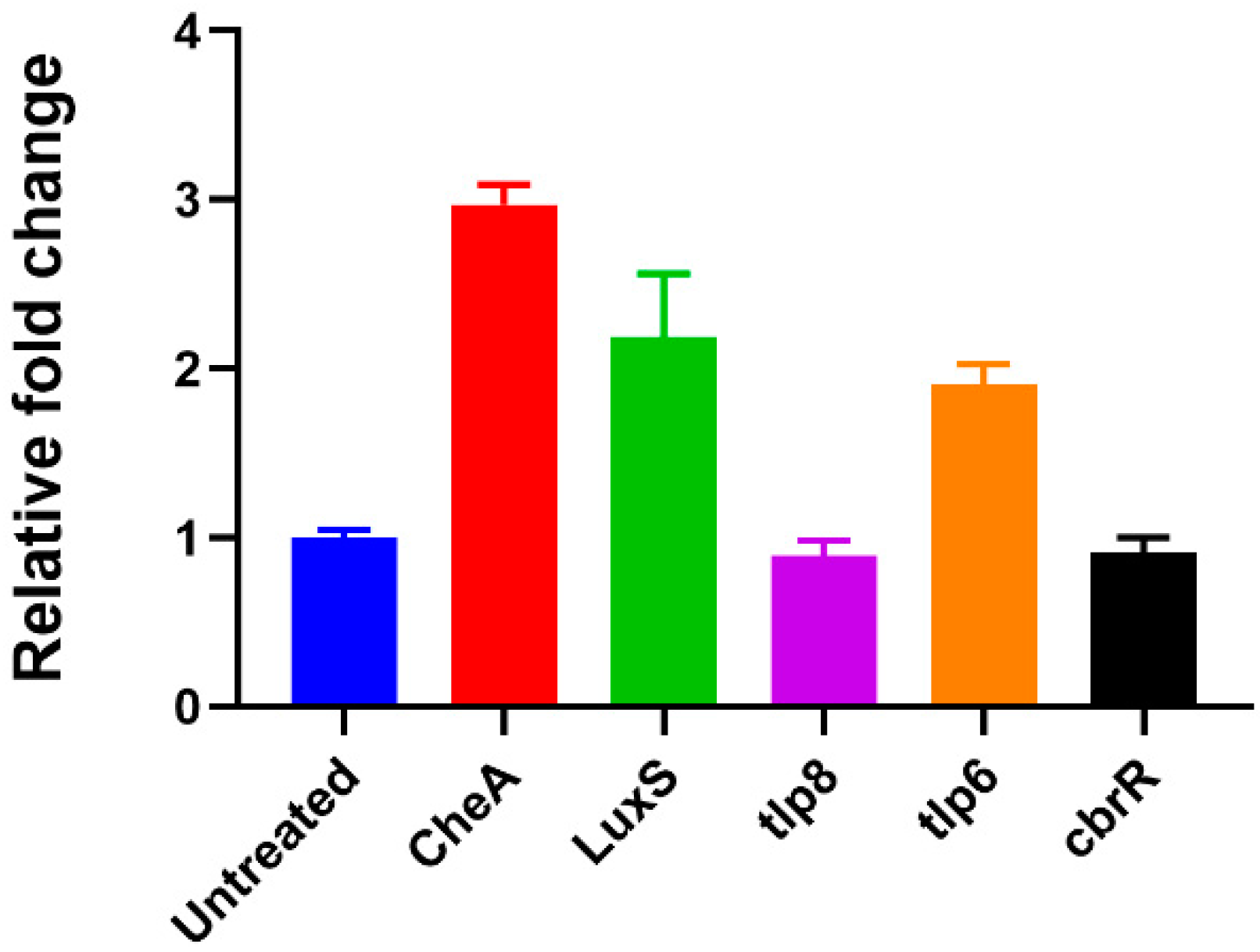
3.7. Extracellular c-di-GMP Dysregulates Specific Gene Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Buzby, J.C.; Allos, B.M.; Roberts, T. The economic burden of Campylobacter-associated Guillain-Barré syndrome. J. Infect. Dis 1997, 176, S192–S197. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. The Global View of Campylobacteriosis: Report of an Expert Consultation; WHO Newsletters; World Health Organization: Utrecht, The Netherlands, 2013.
- Salloway, S.; Mermel, L.; Seamans, M.; Aspinall, G.; Shin, J.N.; Kurjanczyk, L.; Penner, J. Miller-Fisher syndrome associated with Campylobacter jejuni bearing lipopolysaccharide molecules that mimic human ganglioside GD3. Infect. Immun. 1996, 64, 2945–2949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van den Berg, B.; Walgaard, C.; Drenthen, J.; Fokke, C.; Jacobs, B.C.; van Doorn, P.A. Guillain-Barre syndrome: Pathogenesis, diagnosis, treatment and prognosis. Nat. Rev. Neurol. 2014, 10, 469–482. [Google Scholar] [CrossRef] [PubMed]
- Whiley, H.; van den Akker, B.; Giglio, S.; Bentham, R. The role of environmental reservoirs in human campylobacteriosis. Int. J. Environ. Res. Public Health 2013, 10, 5886–5907. [Google Scholar] [CrossRef] [Green Version]
- Ellis-Iversen, J.; Ridley, A.; Morris, V.; Sowa, A.; Harris, J.; Atterbury, R.; Sparks, N.; Allen, V. Persistent environmental reservoirs on farms as risk factors for Campylobacter in commercial poultry. Epidemiol. Infect. 2012, 140, 916–924. [Google Scholar] [CrossRef] [PubMed]
- Carrascosa, C.; Raheem, D.; Ramos, F.; Saraiva, A.; Raposo, A. Microbial biofilms in the food industry—A comprehensive review. Int. J. Environ. Res. Public Health 2021, 18, 2014. [Google Scholar] [CrossRef] [PubMed]
- Bronowski, C.; James, C.E.; Winstanley, C. Role of environmental survival in transmission of Campylobacter jejuni. FEMS Microbiol. Lett. 2014, 356, 8–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Backert, S.; Tegtmeyer, N.; Cróinín, T.Ó.; Boehm, M.; Heimesaat, M.M. Human campylobacteriosis. In Campylobacter; Elsevier: Amsterdam, The Netherlands, 2017; pp. 1–25. [Google Scholar]
- Igwaran, A.; Okoh, A.I. Human campylobacteriosis: A public health concern of global importance. Heliyon 2019, 5, e02814. [Google Scholar] [CrossRef] [PubMed]
- Luangtongkum, T.; Jeon, B.; Han, J.; Plummer, P.; Logue, C.M.; Zhang, Q. Antibiotic Resistance in Campylobacter: Emergence, Transmission and Persistence. Future Microbiol. 2009, 4, 189–200. [Google Scholar] [PubMed] [Green Version]
- Ma, L.; Konkel, M.E.; Lu, X. Antimicrobial resistance gene transfer from Campylobacter jejuni in mono-and dual-species biofilms. Appl. Environ. Microbiol. 2021, 87, e00659-21. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.L.; Fratamico, P.M. Fluoroquinolone resistance in Campylobacter. J. Food Prot. 2010, 73, 1141–1152. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Publishes List of Bacteria for Which New Antibiotics Are Urgently Needed; WHO Newsletters; World Health Organization: Geneva, Switzerland, 2017.
- García-Sánchez, L.; Melero, B.; Jaime, I.; Rossi, M.; Ortega, I.; Rovira, J. Biofilm formation, virulence and antimicrobial resistance of different Campylobacter jejuni isolates from a poultry slaughterhouse. Food Microbiol. 2019, 83, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Magajna, B.A.; Schraft, H. Campylobacter jejuni biofilm cells become viable but non-culturable (VBNC) in low nutrient conditions at 4 C more quickly than their planktonic counterparts. Food Control 2015, 50, 45–50. [Google Scholar] [CrossRef]
- Lv, R.; Wang, K.; Feng, J.; Heeney, D.D.; Liu, D.; Lu, X. Detection and quantification of viable but non-culturable Campylobacter jejuni. Front. Microbiol. 2020, 10, 2920. [Google Scholar] [CrossRef] [PubMed]
- Pitkänen, T. Review of Campylobacter spp. in drinking and environmental waters. J. Microbiol. Methods 2013, 95, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Stetsenko, V.; Efimochkina, N.; Pichugina, T. Growth and persistence of Campylobacter jejuni in foodstuffs. Bull. Exp. Biol. Med. 2019, 166, 759–765. [Google Scholar] [CrossRef]
- Kim, S.-H.; Chelliah, R.; Ramakrishnan, S.R.; Perumal, A.S.; Bang, W.-S.; Rubab, M.; Daliri, E.B.-M.; Barathikannan, K.; Elahi, F.; Park, E. Review on stress tolerance in Campylobacter jejuni. Front. Cell. Infect. Microbiol. 2020, 10, 596570. [Google Scholar] [CrossRef] [PubMed]
- Reuter, M.; Mallett, A.; Pearson, B.M.; van Vliet, A.H. Biofilm formation by Campylobacter jejuni is increased under aerobic conditions. Appl. Environ. Microbiol. 2010, 76, 2122–2128. [Google Scholar] [CrossRef] [Green Version]
- Hanning, I.; Jarquin, R.; Slavik, M. Campylobacter jejuni as a secondary colonizer of poultry biofilms. J. Appl. Microbiol. 2008, 105, 1199–1208. [Google Scholar] [CrossRef] [PubMed]
- Hanning, I.; Donoghue, D.; Jarquin, R.; Kumar, G.; Aguiar, V.; Metcalf, J.; Reyes-Herrera, I.; Slavik, M. Campylobacter biofilm phenotype exhibits reduced colonization potential in young chickens and altered in vitro virulence. Poult. Sci. 2009, 88, 1102–1107. [Google Scholar] [CrossRef] [PubMed]
- Alary, M.; Nadeau, D. An outbreak of Campylobacter enteritis associated with a community water supply. Can. J. Public Health. 1989, 81, 268–271. [Google Scholar]
- Sung, K.; Khan, S. Biofilm development by Campylobacter Jejuni. Biofilms Food Environ. 2015, 24, 29–50. [Google Scholar]
- Ica, T.; Caner, V.; Istanbullu, O.; Nguyen, H.D.; Ahmed, B.; Call, D.R.; Beyenal, H. Characterization of mono- and mixed-culture Campylobacter jejuni biofilms. Appl. Environ. Microbiol 2012, 78, 1033–1038. [Google Scholar] [CrossRef] [Green Version]
- Zhong, X.; Wu, Q.; Zhang, J.; Ma, Z.; Wang, J.; Nie, X.; Ding, Y.; Xue, L.; Chen, M.; Wu, S. Campylobacter jejuni biofilm formation under aerobic conditions and inhibition by ZnO nanoparticles. Front. Microbiol. 2020, 11, 207. [Google Scholar] [CrossRef]
- Joshua, G.P.; Guthrie-Irons, C.; Karlyshev, A.; Wren, B. Biofilm formation in Campylobacter jejuni. Microbiology 2006, 152, 387–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iovine, N.M. Resistance mechanisms in Campylobacter jejuni. Virulence 2013, 4, 230–240. [Google Scholar] [CrossRef] [Green Version]
- Mishra, R.; Panda, A.K.; De Mandal, S.; Shakeel, M.; Bisht, S.S.; Khan, J. Natural anti-biofilm agents: Strategies to control biofilm-forming pathogens. Front. Microbiol. 2020, 11, 2640. [Google Scholar] [CrossRef] [PubMed]
- Elgamoudi, B.A.; Korolik, V. Campylobacter biofilms: Potential of natural compounds to disrupt Campylobacter jejuni transmission. Int. J. Mol. Sci. 2021, 22, 12159. [Google Scholar] [CrossRef] [PubMed]
- Roy, R.; Tiwari, M.; Donelli, G.; Tiwari, V. Strategies for combating bacterial biofilms: A focus on anti-biofilm agents and their mechanisms of action. Virulence 2018, 9, 522–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenal, U.; Reinders, A.; Lori, C. Cyclic di-GMP: Second messenger extraordinaire. Nat. Rev. Microbiol. 2017, 15, 271–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, C.J.; Utada, A.; Davis, K.R.; Thongsomboon, W.; Sanchez, D.Z.; Banakar, V.; Cegelski, L.; Wong, G.C.; Yildiz, F.H. C-di-GMP regulates motile to sessile transition by modulating MshA pili biogenesis and near-surface motility behavior in Vibrio cholerae. PLoS Pathog. 2015, 11, e1005068. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.H.; Koseoglu, V.K.; Guvener, Z.T.; Myers-Morales, T.; Reed, J.M.; D’Orazio, S.E.; Miller, K.W.; Gomelsky, M. Cyclic di-GMP-dependent signaling pathways in the pathogenic Firmicute Listeria monocytogenes. PLoS Pathog. 2014, 10, e1004301. [Google Scholar] [CrossRef] [PubMed]
- Kalia, D.; Merey, G.; Nakayama, S.; Zheng, Y.; Zhou, J.; Luo, Y.; Guo, M.; Roembke, B.T.; Sintim, H.O. Nucleotide, c-di-GMP, c-di-AMP, cGMP, cAMP, (p)ppGpp signaling in bacteria and implications in pathogenesis. Chem. Soc. Rev. 2013, 42, 305–341. [Google Scholar] [CrossRef] [PubMed]
- Romling, U.; Galperin, M.Y.; Gomelsky, M. Cyclic di-GMP: The first 25 years of a universal bacterial second messenger. Microbiol. Mol. Biol. Rev. 2013, 77, 1–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chua, S.L.; Liu, Y.; Yam, J.K.; Chen, Y.; Vejborg, R.M.; Tan, B.G.; Kjelleberg, S.; Tolker-Nielsen, T.; Givskov, M.; Yang, L. Dispersed cells represent a distinct stage in the transition from bacterial biofilm to planktonic lifestyles. Nat. Commun. 2014, 5, 4462. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Jayaraman, N.; Chatterji, D. Small-molecule inhibition of bacterial biofilm. ACS Omega 2020, 5, 3108–3115. [Google Scholar] [CrossRef]
- Poulin, M.B.; Kuperman, L.L. Regulation of biofilm exopolysaccharide production by cyclic di-guanosine monophosphate. Front. Microbiol. 2021, 12, 730980. [Google Scholar] [CrossRef] [PubMed]
- Reichhardt, C.; Parsek, M.R. Confocal laser scanning microscopy for analysis of Pseudomonas aeruginosa biofilm architecture and matrix localization. Front. Microbiol. 2019, 10, 677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomelsky, M.; Galperin, M.Y. Bacterial second messengers, cGMP and c-di-GMP, in a quest for regulatory dominance. EMBO J. 2013, 32, 2421–2423. [Google Scholar] [CrossRef] [Green Version]
- Brouillette, E.; Hyodo, M.; Hayakawa, Y.; Karaolis, D.K.; Malouin, F. 3′,5′-cyclic diguanylic acid reduces the virulence of biofilm-forming Staphylococcus aureus strains in a mouse model of mastitis infection. Antimicrob. Agents Chemother. 2005, 49, 3109–3113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karaolis, D.K.; Means, T.K.; Yang, D.; Takahashi, M.; Yoshimura, T.; Muraille, E.; Philpott, D.; Schroeder, J.T.; Hyodo, M.; Hayakawa, Y. Bacterial c-di-GMP is an immunostimulatory molecule. J. Immunol. 2007, 178, 2171–2181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfe, A.J.; Visick, K.L. Get the message out: Cyclic-Di-GMP regulates multiple levels of flagellum-based motility. J. Bacteriol. 2008, 190, 463–475. [Google Scholar] [CrossRef] [PubMed]
- David, K.R.K.; Rashid, M.H.; Chythanya, R.; Luo, W.; Hyodo, M.; Hayakawa, Y. c-di-GMP (3′-5′-Cyclic Diguanylic Acid) inhibits Staphylococcus aureus cell-cell interactions and biofilm formation. Antimicrob. Agents Chemother. 2005, 49, 1029–1038. [Google Scholar]
- Yan, W.; Qu, T.; Zhao, H.; Su, L.; Yu, Q.; Gao, J.; Wu, B. The effect of c-di-GMP (3’-5’-cyclic diguanylic acid) on the biofilm formation and adherence of Streptococcus mutans. Microbiol. Res. 2010, 165, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Cox, C.A.; Bogacz, M.; El Abbar, F.M.; Browning, D.D.; Hsueh, B.Y.; Waters, C.M.; Lee, V.T.; Thompson, S.A. The Campylobacter jejuni response regulator and Cyclic-Di-GMP binding CbrR is a novel regulator of flagellar motility. Microorganisms 2021, 10, 86. [Google Scholar] [CrossRef] [PubMed]
- Day; Hartley-Tassell, L.E.; Shewell, L.K.; King, R.M.; Tram, G.; Day, S.K.; Semchenko, E.A.; Korolik, V. Variation of chemosensory receptor content of Campylobacter jejuni strains and modulation of receptor gene expression under different in vivo and in vitro growth conditions. BMC Microbiol. 2012, 12, 128. [Google Scholar]
- Elgamoudi, B.A.; Taha, T.; Korolik, V. Inhibition of Campylobacter jejuni biofilm formation by D-amino acids. Antibiotics 2020, 9, 836. [Google Scholar] [CrossRef] [PubMed]
- Tram, G.; Korolik, V.; Day, C.J. MBDS solvent: An improved method for assessment of biofilms. Adv. Microbiol. 2013, 3, 5. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.; Patil, A.; Prabhune, A.; Goel, G. Inhibition of quorum-sensing-mediated biofilm formation in Cronobacter sakazakii strains. Microbiology 2016, 162, 1708–1714. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.-R.; Wu, Y.-P.; Elena, G.; Zhong, K.; Gao, H. Insight into the effect of quinic acid on biofilm formed by Staphylococcus aureus. RSC Adv. 2019, 9, 3938–3945. [Google Scholar] [CrossRef] [Green Version]
- Sorroche, F.G.; Spesia, M.B.; Zorreguieta, Á.; Giordano, W. A positive correlation between bacterial autoaggregation and biofilm formation in native Sinorhizobium meliloti isolates from Argentina. Appl. Environ. Microbiol. 2012, 78, 4092–4101. [Google Scholar] [CrossRef] [Green Version]
- Day, C.J.; King, R.M.; Shewell, L.K.; Tram, G.; Najnin, T.; Hartley-Tassell, L.E.; Wilson, J.C.; Fleetwood, A.D.; Zhulin, I.B.; Korolik, V. A direct-sensing galactose chemoreceptor recently evolved in invasive strains of Campylobacter jejuni. Nat. Commun. 2016, 7, 13206. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; He, Y.; Irwin, P.L.; Jin, T.; Shi, X. Antibacterial activity and mechanism of action of zinc oxide nanoparticles against Campylobacter jejuni. Appl. Environ. Microbiol. 2011, 77, 2325–2331. [Google Scholar] [CrossRef] [Green Version]
- Yan, X. Omics, Microbial Modeling and Technologies for Foodborne Pathogens; DEStech Publications, Inc.: Lancaster, PA, USA, 2012. [Google Scholar]
- Wösten, M.M.; Wagenaar, J.A.; van Putten, J.P. The FlgS/FlgR two-component signal transduction system regulates the fla regulon in Campylobacter jejuni. J. Biol. Chem. 2004, 279, 16214–16222. [Google Scholar] [CrossRef]
- Drozd, M.; Chandrashekhar, K.; Rajashekara, G. Polyphosphate-mediated modulation of Campylobacter jejuni biofilm growth and stability. Virulence 2014, 5, 680–690. [Google Scholar] [CrossRef] [Green Version]
- McLennan, M.K.; Ringoir, D.D.; Frirdich, E.; Svensson, S.L.; Wells, D.H.; Jarrell, H.; Szymanski, C.M.; Gaynor, E.C. Campylobacter jejuni biofilms up-regulated in the absence of the stringent response utilize a calcofluor white-reactive polysaccharide. J. Bacteriol. 2008, 190, 1097–1107. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.; Veeregowda, D.H.; van de Belt-Gritter, B.; Busscher, H.J.; van der Mei, H.C. Extracellular polymeric matrix production and relaxation under fluid shear and mechanical pressure in Staphylococcus aureus biofilms. Appl. Environ. Microbiol. 2018, 84, e01516-17. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Li, S.; Liu, X.; Wang, Z.; Jiang, M.; Wang, R.; Xie, L.; Liu, Q.; Xie, X.; Shang, D. Sensing of autoinducer-2 by functionally distinct receptors in prokaryotes. Nat. Commun. 2020, 11, 5371. [Google Scholar] [CrossRef]
- Cloak, O.M.; Solow, B.T.; Briggs, C.E.; Chen, C.-Y.; Fratamico, P.M. Quorum sensing and production of autoinducer-2 in Campylobacter spp., Escherichia coli O157: H7, and Salmonella enterica serovar Typhimurium in foods. Appl. Environ. Microbiol. 2002, 68, 4666–4671. [Google Scholar] [CrossRef] [Green Version]
- Bezek, K.; Kurinčič, M.; Knauder, E.; Klančnik, A.; Raspor, P.; Bucar, F.; Smole Možina, S. Attenuation of adhesion, biofilm formation and quorum sensing of Campylobacter jejuni by Euodia ruticarpa. Phytother Res. 2016, 30, 1527–1532. [Google Scholar] [CrossRef]
- Tram, G.; Klare, W.P.; Cain, J.A.; Mourad, B.; Cordwell, S.J.; Day, C.J.; Korolik, V. Assigning a role for chemosensory signal transduction in Campylobacter jejuni biofilms using a combined omics approach. Sci. Rep. 2019, 10, 6829. [Google Scholar] [CrossRef] [Green Version]
- Tram, G.; Klare, W.P.; Cain, J.A.; Mourad, B.; Cordwell, S.J.; Korolik, V.; Day, C.J. RNA sequencing data sets identifying differentially expressed transcripts during Campylobacter jejuni biofilm formation. Microbiol. Resour. Announc. 2020, 9, e00982-19. [Google Scholar] [CrossRef] [Green Version]
- Rumbaugh, K.P.; Sauer, K. Biofilm dispersion. Nat. Rev. Microbiol. 2020, 18, 571–586. [Google Scholar] [CrossRef]
- Sintim, H.O.; Smith, J.A.; Wang, J.; Nakayama, S.; Yan, L. Paradigm shift in discovering next-generation anti-infective agents: Targeting quorum sensing, c-di-GMP signaling and biofilm formation in bacteria with small molecules. Future Med. Chem. 2010, 2, 1005–1035. [Google Scholar] [CrossRef]
- Kaplan, J.B. Biofilm dispersal: Mechanisms, clinical implications, and potential therapeutic uses. J. Dent. Res. 2010, 89, 205–218. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Conover, M.; Lu, H.; Parsek, M.R.; Bayles, K.; Wozniak, D.J. Assembly and development of the Pseudomonas aeruginosa biofilm matrix. PLoS Pathog. 2009, 5, e1000354. [Google Scholar] [CrossRef] [Green Version]
- Goodwine, J.; Gil, J.; Doiron, A.; Valdes, J.; Solis, M.; Higa, A.; Davis, S.; Sauer, K. Pyruvate-depleting conditions induce biofilm dispersion and enhance the efficacy of antibiotics in killing biofilms in vitro and in vivo. Sci. Rep. 2019, 9, 3763. [Google Scholar] [CrossRef] [Green Version]
- Tereň, M.; Shagieva, E.; Vondrakova, L.; Viktorova, J.; Svarcova, V.; Demnerova, K.; Michova, H.T. Mutagenic strategies against luxS gene affect the early stage of biofilm formation of Campylobacter jejuni. J. Appl. Genet. 2021, 63, 145–157. [Google Scholar] [CrossRef]
- Püning, C.; Su, Y.; Lu, X.; Gölz, G. Molecular mechanisms of Campylobacter biofilm formation and quorum sensing. Fight. Campylobacter Infect. 2021, 431, 293–319. [Google Scholar] [CrossRef]
- Solano, C.; Echeverz, M.; Lasa, I. Biofilm dispersion and quorum sensing. Curr. Opin. Microbiol. 2014, 18, 96–104. [Google Scholar] [CrossRef] [Green Version]
- Waters, C.M.; Lu, W.; Rabinowitz, J.D.; Bassler, B.L. Quorum sensing controls biofilm formation in Vibrio cholerae through modulation of Cyclic Di-GMP levels and repression of vpsT. J. Bacteriol. 2008, 190, 2527–2536. [Google Scholar] [CrossRef] [Green Version]
- Elvers, K.T.; Park, S.F. Quorum sensing in Campylobacter jejuni: Detection of a luxS encoded signalling molecule. Microbiology 2002, 148, 1475–1481. [Google Scholar] [CrossRef] [Green Version]
- Nwoko, E.-s.Q.; Okeke, I.N. Bacteria autoaggregation: How and why bacteria stick together. Biochem. Soc. Trans. 2021, 49, 1147–1157. [Google Scholar] [CrossRef]
- Trunk, T.; Khalil, H.S.; Leo, J.C. Bacterial autoaggregation. AIMS Microbiol. 2018, 4, 140. [Google Scholar] [CrossRef]
- Formosa-Dague, C.; Feuillie, C.; Beaussart, A.; Derclaye, S.; Kucharikova, S.; Lasa, I.; Van Dijck, P.; Dufrene, Y.F. Sticky matrix: Adhesion mechanism of the staphylococcal polysaccharide intercellular adhesin. ACS Nano 2016, 10, 3443–3452. [Google Scholar] [CrossRef]
- Laganenka, L.; Colin, R.; Sourjik, V. Chemotaxis towards autoinducer 2 mediates autoaggregation in Escherichia coli. Nat. Commun. 2016, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Ramić, D.; Klančnik, A.; Možina, S.S.; Dogsa, I. Elucidation of the AI-2 communication system in the food-borne pathogen Campylobacter jejuni by whole-cell-based biosensor quantification. Biosens. Bioelectron. 2022, 30, 114439. [Google Scholar] [CrossRef]
- Song, S.; Wood, T.K. The primary physiological roles of autoinducer 2 in Escherichia coli are chemotaxis and biofilm formation. Microorganisms 2021, 9, 386. [Google Scholar] [CrossRef]
- Xu, L.; Xin, L.; Zeng, Y.; Yam, J.K.H.; Ding, Y.; Venkataramani, P.; Cheang, Q.W.; Yang, X.; Tang, X.; Zhang, L.-H. A cyclic di-GMP–binding adaptor protein interacts with a chemotaxis methyltransferase to control flagellar motor switching. Sci. Signal. 2016, 9, ra102. [Google Scholar] [CrossRef]
- Korolik, V. The role of chemotaxis during Campylobacter jejuni colonisation and pathogenesis. Curr. Opin. Microbiol. 2019, 47, 32–37. [Google Scholar] [CrossRef]
- Jeon, B.; Itoh, K.; Misawa, N.; Ryu, S. Effects of quorum sensing on flaA transcription and autoagglutination in Campylobacter jejuni. Microbiol. Immunol. 2003, 47, 833–839. [Google Scholar] [CrossRef]
- Steinberger, O.; Lapidot, Z.; Ben-Ishai, Z.; Amikam, D. Elevated expression of the CD4 receptor and cell cycle arrest are induced in Jurkat cells by treatment with the novel cyclic dinucleotide 3′, 5′-cyclic diguanylic acid. FEBS Lett. 1999, 444, 125–129. [Google Scholar] [CrossRef] [Green Version]
- Townsley, L.; Yannarell, S.M.; Huynh, T.N.; Woodward, J.J.; Shank, E.A. Cyclic di-AMP acts as an extracellular signal that impacts Bacillus subtilis biofilm formation and plant attachment. mBio 2018, 9, e00341-18. [Google Scholar] [CrossRef] [Green Version]
- Unterholzner, L. Beyond sensing DNA: A role for cGAS in the detection of extracellular cyclic di-nucleotides. EMBO Rep. 2019, 20, e47970. [Google Scholar] [CrossRef]
- Chen, Z.-h.; Schaap, P. Secreted cyclic di-GMP induces stalk cell differentiation in the eukaryote Dictyostelium discoideum. J. Bacteriol. 2016, 198, 27–31. [Google Scholar] [CrossRef] [Green Version]
- Schaap, P. Cyclic di-nucleotide signaling enters the eukaryote domain. IUBMB Life 2013, 65, 897–903. [Google Scholar] [CrossRef]
- Amarante-Mendes, G.P.; Adjemian, S.; Branco, L.M.; Zanetti, L.C.; Weinlich, R.; Bortoluci, K.R. Pattern recognition receptors and the host cell death molecular machinery. Front. Immunol. 2018, 9, 2379. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elgamoudi, B.A.; Starr, K.S.; Korolik, V. Extracellular c-di-GMP Plays a Role in Biofilm Formation and Dispersion of Campylobacter jejuni. Microorganisms 2022, 10, 2030. https://doi.org/10.3390/microorganisms10102030
Elgamoudi BA, Starr KS, Korolik V. Extracellular c-di-GMP Plays a Role in Biofilm Formation and Dispersion of Campylobacter jejuni. Microorganisms. 2022; 10(10):2030. https://doi.org/10.3390/microorganisms10102030
Chicago/Turabian StyleElgamoudi, Bassam A., Kirstie S. Starr, and Victoria Korolik. 2022. "Extracellular c-di-GMP Plays a Role in Biofilm Formation and Dispersion of Campylobacter jejuni" Microorganisms 10, no. 10: 2030. https://doi.org/10.3390/microorganisms10102030