
Short- and Long-Term Evaluation of a Fixed Dose of Beauveria bassiana Strain MS-8 Diluted in Various Doses of Kaolin as a Powder Formulation Applied to Rice Grains to Control Almond Moth, Ephestia cautella Walker (Lepidoptera: Pyralidae)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect rearing
2.2. Fungus and Dry Conidia Preparations
2.3. Grain Preparations
2.4. Effect of B. bassiana Powder Formulations on Larval Mortality and Adults Emerged in the Short-Term Trial
2.5. Persistence of B. bassiana Powder Formulations against E. cautella in the Long-Term Trial
2.6. Data Collection
2.7. Statistical Analysis
3. Results
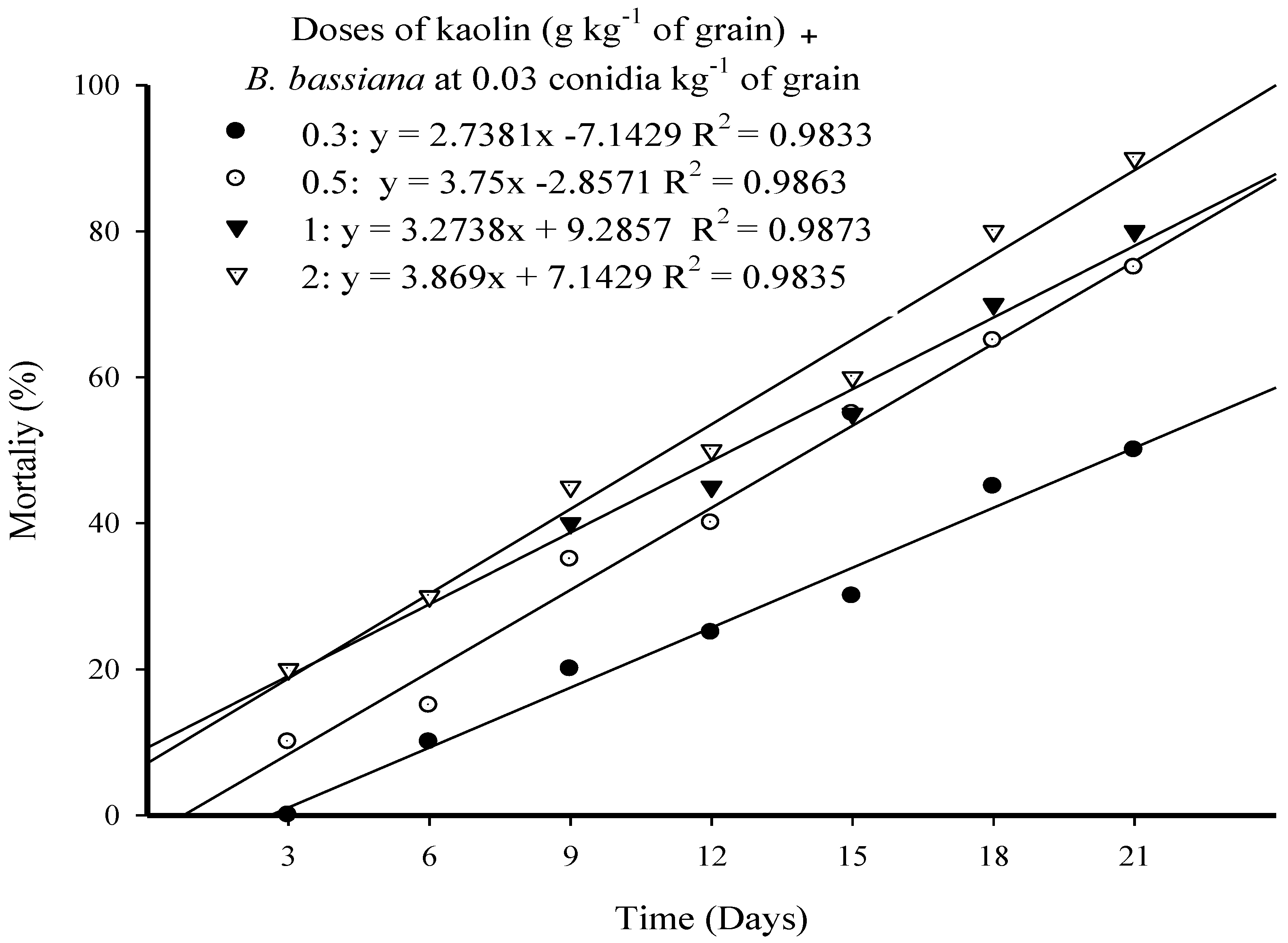
3.1. Effect of B. bassiana Powder Formulations on Larval Mortality
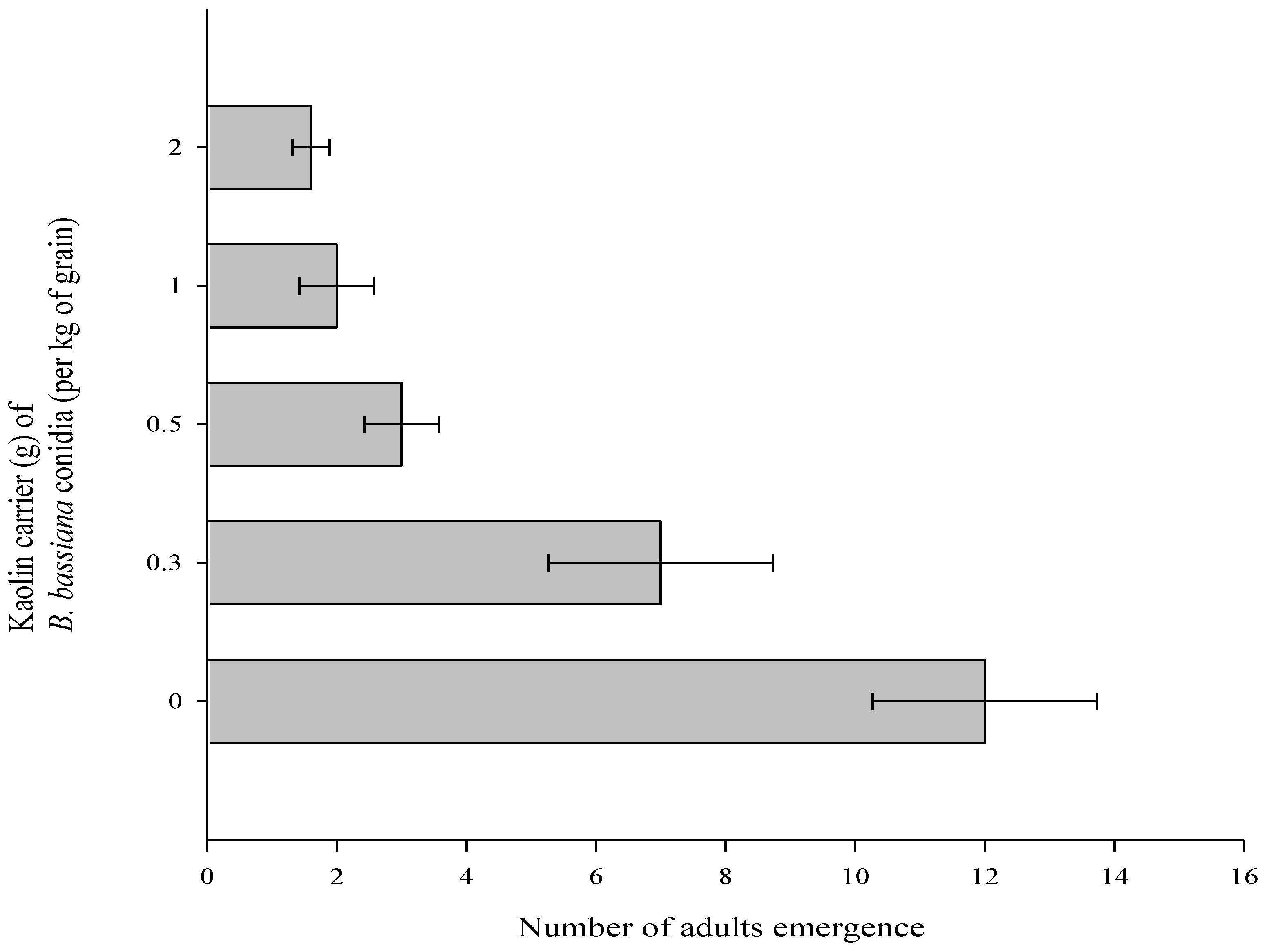
3.2. Effect of B. bassiana Powder Formulations on Number of Adults Emerged
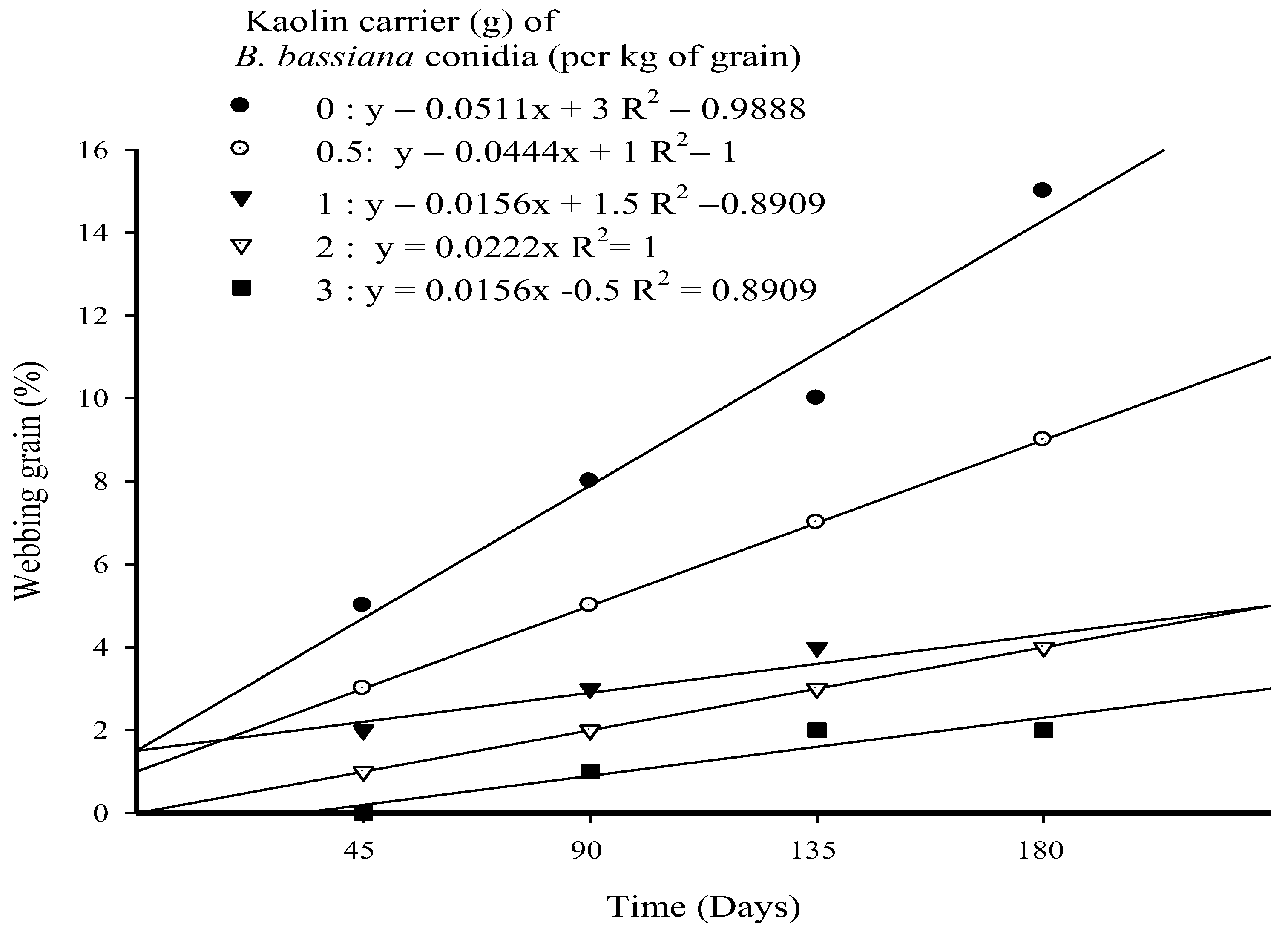
3.3. Webbed Grain (%) in Long-Term Trial
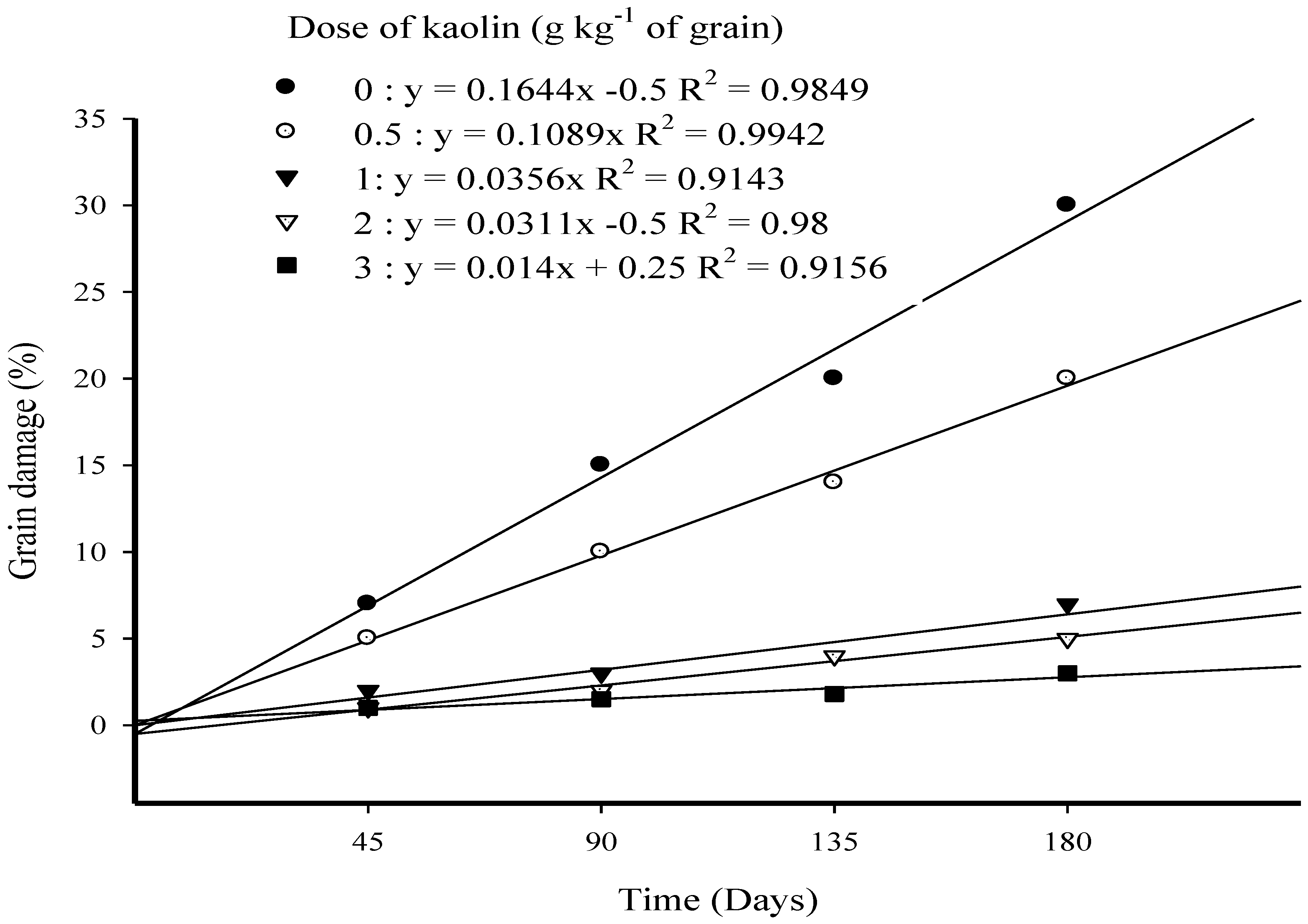
3.4. Grain Damage (%) in Long-Term Trial
3.5. Grain Weight Loss (%) in Long-Term Trial
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Madrid, F.; White, N.; Loschiavo, S. Insects in stored cereals, and their association with farming practices in southern Manitoba. Can. Entomol. 1990, 122, 515–523. [Google Scholar] [CrossRef]
- Tripathi, A.K.; Upadhyay, S.; Bhuiyan, M.; Bhattacharya, P. A review on prospects of essential oils as a biopesticide in insect-pest management. J. Pharmacogn. Phytother. 1991, 1, 52–63. [Google Scholar]
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2015, 144, 31–43. [Google Scholar] [CrossRef]
- Dhaliwal, G.; Jindal, V.; Dhawan, A. Insect pest problems and crop losses: Changing trends. Indian J. Ecol. 2010, 37, 1–7. [Google Scholar]
- IRRI. Global Rice Science Partnership (GRiSP); International Rice Research Institute: Los Baños, PI, USA; Africa Rice Center: Cotonou, Benin; International Center for Tropical Agriculture: Cali, Colombia, 2010. [Google Scholar]
- Youdeowei, A. Fighting Hunger and Poverty: IPM Contributions in Africa. In Proceedings of the Plenary Lecture, 15th International Plant Protection Congress, Beijing, China, 11 July 2003. [Google Scholar]
- Arbogast, R.; Chini, S.; Kendra, P. Infestation of stored saw palmetto berries by Cadra cautella (Lepidoptera: Pyralidae) and the host paradox in stored-product insects. Fla. Entomol. 2005, 88, 314–320. [Google Scholar] [CrossRef]
- Ofuya, T.I.; Lale, N.E.S. Pests of Stored Cereals and Pulses in Nigeria. Biology, Ecology and Control; Dave Collins Publishers: Akure, Nigeria, 2001. [Google Scholar]
- Semple, R.; Frio, A.; Hicks, P.; Lozare, J. Mycotoxin Prevention and Control in Food Grains: A Collaborative Publication of the UNDP/FAO Regional Network Inter-Country Cooperation on Preharvest Technology and Quality Control of Food grains (REGNET) and the ASEAN Grain Postharvest Programme, Bangkok, Thailand. Available online: https://www.fao.org/3/x5036e/5036E00.htm (accessed on 20 September 2022).
- Benhalima, H.; Chaudhry, M.; Mills, K.; Price, N. Phosphine resistance in stored-product insects collected from various grain storage facilities in Morocco. J. Stored Prod. Res. 2004, 40, 241–249. [Google Scholar] [CrossRef]
- Jittanun, C.; Chongrattanameteekul, W. Phosphine resistance in Thai local strains of Tribolium castaneum (Herbst) and their response to synthetic pheromone. Kasetsart J. (Nat. Sci.) 2012, 48, 9–16. [Google Scholar]
- Nguyen, T.T.; Collins, P.J.; Ebert, P.R. Inheritance and characterization of strong resistance to phosphine in Sitophilus oryzae (L.). PLoS ONE 2015. [Google Scholar] [CrossRef] [Green Version]
- Pimentel, M.; Faroni, L.D.A.; Guedes, R.; Sousa, A.; Tótola, M. Phosphine resistance in Brazilian populations of Sitophilus zeamais Motschulsky (Coleoptera: Curculionidae). J. Stored Prod. Res. 2009, 45, 71–74. [Google Scholar] [CrossRef]
- Araújo, R.A.; Williamson, M.S.; Bass, C.; Field, L.M.; Duce, I.R. Pyrethroid resistance in Sitophilus zeamais is associated with a mutation (T929I) in the voltage-gated sodium channel. Insect Mol. Biol. 2011, 20, 437–445. [Google Scholar] [CrossRef]
- Bughio, F.; Wilkins, R. Influence of malathion resistance status on survival and growth of Tribolium castaneum (Coleoptera: Tenebrionidae), when fed on flour from insect-resistant and susceptible grain rice cultivars. J. Stored Prod. Res. 2004, 40, 65–75. [Google Scholar] [CrossRef]
- Guedes, R.N.C.; Lima, J.G.; Santos, J.P.; Cruz, C.D. Resistance to DDT and pyrethroids in Brazilian populations of Sitophilus zeamais Motsch. (Coleoptera: Curculionidae). J. Stored Prod. Res. 1995, 31, 145–150. [Google Scholar] [CrossRef]
- Pereira, C.; Pereira, E.; Cordeiro, E.; Della Lucia, T.; Tótola, M.; Guedes, R. Organophosphate resistance in the maize weevil Sitophilus zeamais: Magnitude and behavior. Crop Prot. 2014, 28, 168–173. [Google Scholar] [CrossRef]
- Schöller, M.; Prozell, S.; Al-Kirshi, A.G.; Reichmuth, C. Towards biological control as a major component of integrated pest management in stored product protection. J. Stored Prod. Res. 1997, 33, 81–97. [Google Scholar] [CrossRef]
- Wakil, W.; Ghazanfar, M.U. Entomopathogenic fungus as a biological control agent against Rhyzopertha dominica F. (Coleoptera: Bostrychidae) on stored wheat. Arch. Phytopathol. Plant Prot. 2010, 43, 1236–1242. [Google Scholar] [CrossRef]
- Cherry, A.; Abalo, P.; Hell, K. A laboratory assessment of the potential of different strains of the entomopathogenic fungi Beauveria bassiana (Balsamo) Vuillemin and Metarhizium anisopliae (Metschnikoff) to control Callosobruchus maculatus (F.) (Coleoptera: Bruchidae) in stored cowpea. J. Stored Prod. Res. 2005, 41, 295–309. [Google Scholar] [CrossRef]
- Crespo, R.; Júrez, M.; Dal Bello, G.; Padin, S.; Fernández, G.C.; Pedrini, N. Increased mortality of Acanthoscelides obtectus by alkane-grown Beauveria bassiana. BioControl 2002, 47, 685–696. [Google Scholar] [CrossRef]
- Kaur, S.; Thakur, A.; Rajput, M. A laboratory assessment of the potential of Beauveria bassiana (Balsamo) Vuillemin as a biocontrol agent of Corcyra cephalonica Stainton (Lepidoptera: Pyralidae). J. Stored Prod. Res. 2014, 59, 185–189. [Google Scholar] [CrossRef]
- Sabbour, M.; Abd-El-Aziz, S.; Sherief, M. Efficacy of three entomopathogenic fungi alone or in combination with diatomaceous earth modifications for the control of three pyralid moths in stored grains. J. Plant Prot. Res. 2012, 52, 359–363. [Google Scholar] [CrossRef]
- Akbar, W.; Lord, J.C.; Nechols, J.R.; Howard, R.W. Diatomaceous earth increases the efficacy of Beauveria bassiana against Tribolium castaneum larvae and increases conidia attachment. J. Econ. Entomol. 2004, 97, 273–280. [Google Scholar] [CrossRef]
- Athanassiou, C.; Steenberg, T. Insecticidal effect of Beauveria bassiana (Balsamo) Vuillemin (Ascomycota: Hypocreales) in combination with three diatomaceous earth formulations against Sitophilus granarius (L.) (Coleoptera: Curculionidae). Biol. Control. 2007, 40, 411–416. [Google Scholar] [CrossRef]
- Batta, Y. Control of rice weevil (Sitophilus oryzae L., Coleoptera: Curculionidae) with various formulations of Metarhizium anisopliae. Crop Prot. 2004, 23, 103–108. [Google Scholar] [CrossRef]
- Riasat, T.; Wakil, W.; Ashfaq, M.; Sahi, S.T. Effect of Beauveria bassiana mixed with diatomaceous earth on mortality, mycosis and sporulation of Rhyzopertha dominica on stored wheat. Phytoparasitica 2011, 39, 325–331. [Google Scholar] [CrossRef]
- Zimmermann, G. The Galleria bait method for detection of entomopathogenic fungi in soil. J. Appl. Entomol. 2004, 102, 213–215. [Google Scholar] [CrossRef]
- Humber, R. Identification of entomopathogenic fungi. In Manual of Techniques in Invertebrate Pathology; Lacey, L.A., Ed.; Elsevier: Waltham, MA, USA, 2012. [Google Scholar]
- Gouli, V.; Provost, C.; Gouli, S.; Parker, B.L.; Skinner, M. Productivity of different species of entomopathogenic fungi based on one type of technology. J. Agric. Technol. 2013, 9, 571–580. [Google Scholar]
- Saeed, M.B.E.E.E.M. Biological Control of Three Grain Storage Pests: Maize Weevil, Sitophilus zeamais (Motschulsky), Almond Moth, Ephestia cautella (Walker) and Cigarette Beetle, Lasioderma serricorne (Fabricius), Using Novel Strains of Beauveria bassiana (Balsamo) Vuillemin in Powder Formulation. Doctoral dissertation, University of KwaZulu-Natal, Durban, South Africa, 2017. [Google Scholar]
- Salerno, G.; Rebora, M.; Kovalev, A.; Gorb, E.; Gorb, S. Kaolin nano-powder effect on insect attachment ability. J. Pest Sci. 2020, 93, 315–327. [Google Scholar] [CrossRef]
- Tefera, T.; Mugo, S.; Tende, R.; Likhayo, P. Methods of Screening Maize for Resistance to Stem Borers and Post-harvest Insect Pests; CIMMYT: Nairobi, Kenya, 2011. [Google Scholar]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Payne, R.W.; Murray, D.A.; Harding, S.A.; Baird, D.B.; Soutar, D.M. Genstat for Windows, 17th ed.; VSN International: Hemel Hempstead, UK, 1997. [Google Scholar]
- Samodra, H.; Ibrahim, Y. Effects of dust formulations of three entomopathogenic fungal isolates against Sitophilus oryzae (Coleoptera: Curculionidae) in rice grain. J. Biosci. 2006, 17, 1–7. [Google Scholar]
- Samodra, H.; Ibrahim, Y.B. Effectiveness of selected entomopathogenic fungi in packed rice grain at room temperature against Corcyra cephalonica Stainton. ASEAN J. Sci. Technol. Dev. 2006, 23, 183–192. [Google Scholar] [CrossRef] [Green Version]
- Ghribi, D.; Elleuch, M.; Abdelkefi, L.; Ellouze-Chaabouni, S. Evaluation of larvicidal potency of Bacillus subtilis SPB1 biosurfactant against Ephestia kuehniella (Lepidoptera: Pyralidae) larvae and influence of abiotic factors on its insecticidal activity. J. Stored Prod. Res. 2012, 48, 68–72. [Google Scholar] [CrossRef]
- Padin, S.; Dal Bello, G.; Fabrizio, M. Grain loss caused by Tribolium castaneum, Sitophilus oryzae and Acanthoscelides obtectus in stored durum wheat and beans treated with Beauveria bassiana. J. Stored Prod. Res. 2002, 38, 69–74. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Adult Emerged (%) | Grain Damage (%) | Webbed Grain (%) | Weight Loss (%) | Mortality (%) | |
|---|---|---|---|---|---|
| Adult emerged (%) | - | ||||
| Grain damage (%) | 0.993 *** | - | |||
| Webbed grain (%) | 0.994 *** | 0.989 ** | - | ||
| Weight loss (%) | 0.999 *** | 0.996 *** | 0.991 *** | - | |
| Mortality (%) | −0.996 *** | −0.981 ** | −0.999 *** | −0.995 *** | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saeed, M.B.; Laing, M.D.; Miller, R.M. Short- and Long-Term Evaluation of a Fixed Dose of Beauveria bassiana Strain MS-8 Diluted in Various Doses of Kaolin as a Powder Formulation Applied to Rice Grains to Control Almond Moth, Ephestia cautella Walker (Lepidoptera: Pyralidae). Microorganisms 2022, 10, 1971. https://doi.org/10.3390/microorganisms10101971
Saeed MB, Laing MD, Miller RM. Short- and Long-Term Evaluation of a Fixed Dose of Beauveria bassiana Strain MS-8 Diluted in Various Doses of Kaolin as a Powder Formulation Applied to Rice Grains to Control Almond Moth, Ephestia cautella Walker (Lepidoptera: Pyralidae). Microorganisms. 2022; 10(10):1971. https://doi.org/10.3390/microorganisms10101971
Chicago/Turabian StyleSaeed, Mohamed Baha, Mark D. Laing, and Ray M. Miller. 2022. "Short- and Long-Term Evaluation of a Fixed Dose of Beauveria bassiana Strain MS-8 Diluted in Various Doses of Kaolin as a Powder Formulation Applied to Rice Grains to Control Almond Moth, Ephestia cautella Walker (Lepidoptera: Pyralidae)" Microorganisms 10, no. 10: 1971. https://doi.org/10.3390/microorganisms10101971