
A Case of Persistent Human Pegivirus Infection in Two Separate Pregnancies of a Woman
 , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Samples
2.2. Pegivirus Sequence Analysis
2.3. RT-PCR Validation
2.4. Viral Antibody Analysis
3. Results
3.1. Maternal Medical History
3.2. Viral Sequence Analysis
3.3. VirScan Analysis
3.4. Detection of HPgV by PCR
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CHD | congenital heart disease |
| HCV | hepatitis C virus |
| HIV | human immunodeficiency virus |
| HPgV | human pegivirus |
| HPV | human papillomavirus |
| IVTP | intervillous tissue of the placenta |
| PBL | peripheral blood leukocytes |
| UTR | untranslated region |
References
- Stapleton, J.T.; Foung, S.; Muerhoff, A.S.; Bukh, J.; Simmonds, P. The GB Viruses: A Review and Proposed Classification of GBV-A, GBV-C (HGV), and GBV-D in Genus Pegivirus within the Family Flaviviridae. J. Gen. Virol. 2011, 92, 233–246. [Google Scholar] [CrossRef]
- Chowdhury, A.Y.; Tavis, J.E.; George, S.L. Human Pegivirus (GB Virus C) NS3 Protease Activity Inhibits Induction of the Type I Interferon Response and Is Not Inhibited by HCV NS3 Protease Inhibitors. Virology 2014, 456–457, 300–309. [Google Scholar] [CrossRef]
- Silva, A.d.S.N.; Silva, C.P.; Barata, R.R.; da Silva, P.V.R.; Monteiro, P.D.J.; Lamarão, L.; Burbano, R.M.R.; Nunes, M.R.T.; de Lima, P.D.L. Human Pegivirus (HPgV, GBV-C) RNA in Volunteer Blood Donors from a Public Hemotherapy Service in Northern Brazil. Virol. J. 2020, 17, 153. [Google Scholar] [CrossRef]
- Yu, Y.; Wan, Z.; Wang, J.-H.; Yang, X.; Zhang, C. Review of Human Pegivirus: Prevalence, Transmission, Pathogenesis, and Clinical Implication. Virulence 2022, 13, 324–341. [Google Scholar] [CrossRef]
- Zhang, X.; Li, W.; Zhou, C. Near-Complete Genome Sequence of a Human Pegivirus Variant Isolated from a Hepatitis E Virus-Infected Patient. Microbiol. Resour. Announc. 2019, 8, e01151-18. [Google Scholar] [CrossRef]
- Soliman, H.K.; Abouelhoda, M.; El Rouby, M.N.; Ahmed, O.S.; Esmat, G.; Hassan, Z.K.; Hafez, M.M.; Mehaney, D.A.; Selvaraju, M.; Darwish, R.K.; et al. Whole-Genome Sequencing of Human Pegivirus Variant from an Egyptian Patient Co-Infected with Hepatitis C Virus: A Case Report. Virol. J. 2019, 16, 132. [Google Scholar] [CrossRef]
- Stapleton, J.T.; Williams, C.F.; Xiang, J. GB Virus Type C: A Beneficial Infection? J. Clin. Microbiol. 2004, 42, 3915–3919. [Google Scholar] [CrossRef]
- Fama, A.; Larson, M.C.; Link, B.K.; Habermann, T.M.; Feldman, A.L.; Call, T.G.; Ansell, S.M.; Liebow, M.; Xiang, J.; Maurer, M.J.; et al. Human Pegivirus Infection and Lymphoma Risk: A Systematic Review and Meta-Analysis. Clin. Infect. Dis. 2020, 71, 1221–1228. [Google Scholar] [CrossRef]
- Mohr, E.L.; Stapleton, J.T. GB Virus Type C Interactions with HIV: The Role of Envelope Glycoproteins. J. Viral Hepat. 2009, 16, 757–768. [Google Scholar] [CrossRef]
- Cebriá-Mendoza, M.; Arbona, C.; Larrea, L.; Díaz, W.; Arnau, V.; Peña, C.; Bou, J.V.; Sanjuán, R.; Cuevas, J.M. Deep Viral Blood Metagenomics Reveals Extensive Anellovirus Diversity in Healthy Humans. Sci. Rep. 2021, 11, 6921. [Google Scholar] [CrossRef]
- Kandathil, A.J.; Cox, A.L.; Page, K.; Mohr, D.; Razaghi, R.; Ghanem, K.G.; Tuddenham, S.A.; Hsieh, Y.-H.; Evans, J.L.; Coller, K.E.; et al. Plasma Virome and the Risk of Blood-Borne Infection in Persons with Substance Use Disorder. Nat. Commun. 2021, 12, 6909. [Google Scholar] [CrossRef]
- Savassi-Ribas, F.; Pereira, J.G.; Horta, M.A.P.; Wagner, T.C.S.; Matuck, T.A.; Monteiro de Carvalho, D.B.; Mello, F.C.A.; Varella, R.B.; Soares, C.C. Human Pegivirus-1 Infection in Kidney Transplant Recipients: A Single-Center Experience. J. Med. Virol. 2020, 92, 2961–2968. [Google Scholar] [CrossRef]
- Vu, D.-L.; Cordey, S.; Simonetta, F.; Brito, F.; Docquier, M.; Turin, L.; van Delden, C.; Boely, E.; Dantin, C.; Pradier, A.; et al. Human Pegivirus Persistence in Human Blood Virome after Allogeneic Haematopoietic Stem-Cell Transplantation. Clin. Microbiol. Infect. 2019, 25, 225–232. [Google Scholar] [CrossRef]
- Izumi, T.; Sakata, K.; Okuzaki, D.; Inokuchi, S.; Tamura, T.; Motooka, D.; Nakamura, S.; Ono, C.; Shimokawa, M.; Matsuura, Y.; et al. Characterization of Human Pegivirus Infection in Liver Transplantation Recipients. J. Med. Virol. 2019, 91, 2093–2100. [Google Scholar] [CrossRef]
- George, S.L.; Wünschmann, S.; McCoy, J.; Xiang, J.; Stapleton, J.T. Interactions Between GB Virus Type C and HIV. Curr. Infect. Dis. Rep. 2002, 4, 550–558. [Google Scholar] [CrossRef]
- Berg, T.; Müller, A.R.; Platz, K.P.; Höhne, M.; Bechstein, W.O.; Hopf, U.; Wiedenmann, B.; Neuhaus, P.; Schreier, E. Dynamics of GB Virus C Viremia Early after Orthotopic Liver Transplantation Indicates Extrahepatic Tissues as the Predominant Site of GB Virus C Replication. Hepatology 1999, 29, 245–249. [Google Scholar] [CrossRef]
- Tanaka, E.; Kiyosawa, K.; Shimoda, K.; Hino, K.; Tacke, M.; Schmolke, S.; Engel, A.M.; Hess, G. Evolution of Hepatitis G Virus Infection and Antibody Response to Envelope Protein in Patients with Transfusion-Associated Non-A, Non-B Hepatitis. J. Viral Hepat. 1998, 5, 153–159. [Google Scholar] [CrossRef]
- McLinden, J.H.; Kaufman, T.M.; Xiang, J.; Chang, Q.; Klinzman, D.; Engel, A.M.; Hess, G.; Schmidt, U.; Houghton, M.; Stapleton, J.T. Characterization of an Immunodominant Antigenic Site on GB Virus C Glycoprotein E2 That Is Involved in Cell Binding. J. Virol. 2006, 80, 12131–12140. [Google Scholar] [CrossRef]
- Ruegamer, T.; Hoffmann, R.; Rohrhofer, A.; Audebert, F.; Salzberger, B.; Korn, K.; Schuster, P.; Eichler, J.; Schmidt, B. Inhibition of HIV-1 Infection by Human Pegivirus Type 1-Derived Peptides Is Affected by Human Pegivirus Type 1 Genotype and HIV-1 Coreceptor Tropism. AIDS 2018, 32, 1951–1957. [Google Scholar] [CrossRef]
- Wu, Z.; Wu, Y.; Zhang, W.; Merits, A.; Simmonds, P.; Wang, M.; Jia, R.; Zhu, D.; Liu, M.; Zhao, X.; et al. The First Nonmammalian Pegivirus Demonstrates Efficient In Vitro Replication and High Lymphotropism. J. Virol. 2020, 94, e01150-20. [Google Scholar] [CrossRef]
- McHutchison, J.G.; Nainan, O.V.; Alter, M.J.; Sedghi-Vaziri, A.; Detmer, J.; Collins, M.; Kolberg, J. Hepatitis C and G Co-Infection: Response to Interferon Therapy and Quantitative Changes in Serum HGV-RNA. Hepatology 1997, 26, 1322–1327. [Google Scholar] [CrossRef]
- Hoffmann, R.; Ruegamer, T.; Schaubächer, J.; Rohrhofer, A.; Kirmeß, P.; Fiebig, K.M.; Schmidt, B.; Eichler, J. Exploring Viral Interference Using Peptides: Molecular Determinants of HIV-1 Inhibition by a Peptide Derived from Human Pegivirus-1 Envelope Protein E2. ChemMedChem 2021, 16, 1290–1296. [Google Scholar] [CrossRef]
- Xiang, J.; Wünschmann, S.; Diekema, D.J.; Klinzman, D.; Patrick, K.D.; George, S.L.; Stapleton, J.T. Effect of Coinfection with GB Virus C on Survival among Patients with HIV Infection. N. Engl. J. Med. 2001, 345, 707–714. [Google Scholar] [CrossRef]
- Polgreen, P.M.; Xiang, J.; Chang, Q.; Stapleton, J.T. GB Virus Type C/Hepatitis G Virus: A Non-Pathogenic Flavivirus Associated with Prolonged Survival in HIV-Infected Individuals. Microbes Infect. 2003, 5, 1255–1261. [Google Scholar] [CrossRef]
- Lefrère, J.J.; Sender, A.; Mercier, B.; Mariotti, M.; Pernot, F.; Soulié, J.C.; Malvoisin, A.; Berry, M.; Gabai, A.; Lattes, F.; et al. High Rate of GB Virus Type C/HGV Transmission from Mother to Infant: Possible Implications for the Prevalence of Infection in Blood Donors. Transfusion 2000, 40, 602–607. [Google Scholar] [CrossRef]
- Paternoster, D.; Serena, A.; Santin, M.; Marchiori, S.; Surico, N.; Amoruso, E.; Longo, D.; Gussetti, N. GB Virus C Infection in Pregnancy: Maternal and Perinatal Importance of the Infection. Eur. J. Obs. Gynecol. Reprod. Biol. 2009, 144, 115–118. [Google Scholar] [CrossRef]
- Sperling, R.S.; Shapiro, D.E.; Coombs, R.W.; Todd, J.A.; Herman, S.A.; McSherry, G.D.; O’Sullivan, M.J.; Van Dyke, R.B.; Jimenez, E.; Rouzioux, C.; et al. Maternal Viral Load, Zidovudine Treatment, and the Risk of Transmission of Human Immunodeficiency Virus Type 1 from Mother to Infant. Pediatric AIDS Clinical Trials Group Protocol 076 Study Group. N. Engl. J. Med. 1996, 335, 1621–1629. [Google Scholar] [CrossRef]
- Zanetti, A.R.; Tanzi, E.; Paccagnini, S.; Principi, N.; Pizzocolo, G.; Caccamo, M.L.; D’Amico, E.; Cambiè, G.; Vecchi, L. Mother-to-Infant Transmission of Hepatitis C Virus. Lombardy Study Group on Vertical HCV Transmission. Lancet 1995, 345, 289–291. [Google Scholar] [CrossRef]
- Lin, H.H.; Kao, J.H.; Yeh, K.Y.; Liu, D.P.; Chang, M.H.; Chen, P.J.; Chen, D.S. Mother-to-Infant Transmission of GB Virus C/Hepatitis G Virus: The Role of High-Titered Maternal Viremia and Mode of Delivery. J. Infect. Dis. 1998, 177, 1202–1206. [Google Scholar] [CrossRef]
- Wylie, K.M.; Wylie, T.N.; Buller, R.; Herter, B.; Cannella, M.T.; Storch, G.A. Detection of Viruses in Clinical Samples by Use of Metagenomic Sequencing and Targeted Sequence Capture. J. Clin. Microbiol. 2018, 56, e01123-18. [Google Scholar] [CrossRef] [Green Version]
- Wylie, T.N.; Wylie, K.M.; Herter, B.N.; Storch, G.A. Enhanced Virome Sequencing Using Targeted Sequence Capture. Genome. Res. 2015, 25, 1910–1920. [Google Scholar] [CrossRef]
- Xu, G.J.; Kula, T.; Xu, Q.; Li, M.Z.; Vernon, S.D.; Ndung’u, T.; Ruxrungtham, K.; Sanchez, J.; Brander, C.; Chung, R.T.; et al. Viral Immunology. Comprehensive Serological Profiling of Human Populations Using a Synthetic Human Virome. Science 2015, 348, aaa0698. [Google Scholar] [CrossRef]
- Wylie, T.N.; Wylie, K.M. ViroMatch: A Computational Pipeline for the Detection of Viral Sequences from Complex Metagenomic Data. Microbiol. Resour. Announc. 2021, 10, e01468-20. [Google Scholar] [CrossRef]
- Peng, Y.; Leung, H.C.; Yiu, S.M.; Chin, F.Y. IDBA-UD: A de Novo Assembler for Single-Cell and Metagenomic Sequencing Data with Highly Uneven Depth. Bioinformatics 2012, 28, 1420–1428. [Google Scholar] [CrossRef]
- Ruby, J.G.; Bellare, P.; Derisi, J.L. PRICE: Software for the Targeted Assembly of Components of (Meta) Genomic Sequence Data. G3 2013, 3, 865–880. [Google Scholar] [CrossRef]
- Milne, I.; Bayer, M.; Stephen, G.; Cardle, L.; Marshall, D. Tablet: Visualizing Next-Generation Sequence Assemblies and Mappings. Methods Mol. Biol. 2016, 1374, 253–268. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Pickett, B.E.; Sadat, E.L.; Zhang, Y.; Noronha, J.M.; Squires, R.B.; Hunt, V.; Liu, M.; Kumar, S.; Zaremba, S.; Gu, Z.; et al. ViPR: An Open Bioinformatics Database and Analysis Resource for Virology Research. Nucleic Acids. Res. 2012, 40, D593–D598. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (ITOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Mota, L.D.D.; Finger-Jardim, F.; Silva, C.M.; Germano, F.N.; Nader, M.M.; Gonçalves, C.V.; Luquez, K.Y.S.; Chies, J.A.B.; Groll, A.V.; Hora, V.P.D.; et al. Molecular and Clinical Profiles of Human Pegivirus Type 1 Infection in Individuals Living with HIV-1 in the Extreme South of Brazil. BioMed Res. Int. 2019, 2019, 8048670. [Google Scholar] [CrossRef]
- Fama, A.; Xiang, J.; Link, B.K.; Allmer, C.; Klinzman, D.; Feldman, A.L.; Nowakowski, G.S.; Liebow, M.; Larson, M.C.; Maurer, M.J.; et al. Human Pegivirus Infection and Lymphoma Risk and Prognosis: A North American Study. Br. J. Haematol. 2018, 182, 644–653. [Google Scholar] [CrossRef]
- Mohan, D.; Wansley, D.L.; Sie, B.M.; Noon, M.S.; Baer, A.N.; Laserson, U.; Larman, H.B. PhIP-Seq Characterization of Serum Antibodies Using Oligonucleotide-Encoded Peptidomes. Nat. Protoc. 2018, 13, 1958–1978. [Google Scholar] [CrossRef]
- Mina, M.J.; Kula, T.; Leng, Y.; Li, M.; de Vries, R.D.; Knip, M.; Siljander, H.; Rewers, M.; Choy, D.F.; Wilson, M.S.; et al. Measles Virus Infection Diminishes Preexisting Antibodies That Offer Protection from Other Pathogens. Science 2019, 366, 599–606. [Google Scholar] [CrossRef]
- Pou, C.; Nkulikiyimfura, D.; Henckel, E.; Olin, A.; Lakshmikanth, T.; Mikes, J.; Wang, J.; Chen, Y.; Bernhardsson, A.K.; Gustafsson, A.; et al. The Repertoire of Maternal Anti-Viral Antibodies in Human Newborns. Nat. Med. 2019, 25, 591–596. [Google Scholar] [CrossRef]
- Coller, K.E.; Bruce, V.; Cassidy, M.; Gersch, J.; Frankel, M.B.; Vallari, A.; Cloherty, G.; Hackett, J.; Evans, J.L.; Page, K.; et al. Chronic Human Pegivirus 2 without Hepatitis C Virus Co-Infection. Emerg. Infect. Dis. 2020, 26, 265–272. [Google Scholar] [CrossRef]
- Santos, L.M.; Lobato, R.C.; Barral, M.F.M.; Goncalves, C.V.; da Hora, V.P.; Martinez, A.M.B. Prevalence and Vertical Transmission of Human Pegivirus among Pregnant Women Infected with HIV. Int. J. Gynaecol. Obs. 2017, 138, 113–118. [Google Scholar] [CrossRef]
- Nahmias, A.J.; Walls, K.W.; Stewart, J.A.; Herrmann, K.L.; Flynt, W.J. The ToRCH Complex-Perinatal Infections Associated with Toxoplasma and Rubella, Cytomegol- and Herpes Simplex Viruses. Pediatr. Res. 1971, 5, 405–406. [Google Scholar] [CrossRef]
- Castro-Mejia, J.L.; Deng, L.; Vogensen, F.K.; Reyes, A.; Nielsen, D.S. Extraction and Purification of Viruses from Fecal Samples for Metagenome and Morphology Analyses. Methods Mol. Biol. 2018, 1838, 49–57. [Google Scholar] [CrossRef]
- Ye, Z.; Wang, L.; Yang, T.; Chen, L.; Wang, T.; Chen, L.; Zhao, L.; Zhang, S.; Zheng, Z.; Luo, L.; et al. Maternal Viral Infection and Risk of Fetal Congenital Heart Diseases: A Meta-Analysis of Observational Studies. J. Am. Heart Assoc. 2019, 8, e011264. [Google Scholar] [CrossRef] [Green Version]
- Sharma, V.; Goessling, L.S.; Mullen, C.; Muller, A.; Dailey, B.; Miller, A.; Carvaja, H.; Brar, A.K.; Eghtesady, P. Coxsackieviral Infection during the First Trimester of Pregnancy Leads to Ventricular Septal Defect. Circulation 2017, 136, A20801. [Google Scholar]
- Wang, T.; Li, Q.; Chen, L.; Ni, B.; Sheng, X.; Huang, P.; Zhang, S.; Qin, J. Maternal Viral Infection in Early Pregnancy and Risk of Congenital Heart Disease in Offspring: A Prospective Cohort Study in Central China. Clin. Epidemiol. 2022, 14, 71–82. [Google Scholar] [CrossRef] [PubMed]
- UniProt Consortium UniProt: The Universal Protein Knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [CrossRef]
- Berg, M.G.; Lee, D.; Coller, K.; Frankel, M.; Aronsohn, A.; Cheng, K.; Forberg, K.; Marcinkus, M.; Naccache, S.N.; Dawson, G.; et al. Discovery of a Novel Human Pegivirus in Blood Associated with Hepatitis C Virus Co-Infection. PLoS Pathog. 2015, 11, e1005325. [Google Scholar] [CrossRef]
- Xiang, J.; Martinez-Smith, C.; Gale, M.; Chang, Q.; Labrecque, D.R.; Schmidt, W.N.; Stapleton, J.T. GB Virus Type C NS5A Sequence Polymorphisms: Association with Interferon Susceptibility and Inhibition of PKR-Mediated EIF2alpha Phosphorylation. J. Interferon Cytokine Res. 2005, 25, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Jordier, F.; Deligny, M.-L.; Barré, R.; Robert, C.; Galicher, V.; Uch, R.; Fournier, P.-E.; Raoult, D.; Biagini, P. Human Pegivirus Isolates Characterized by Deep Sequencing from Hepatitis C Virus-RNA and Human Immunodeficiency Virus-RNA-Positive Blood Donations, France. J. Med. Virol. 2019, 91, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Vitrenko, Y.; Kostenko, I.; Kulebyakina, K.; Sorochynska, K. Prevalence of Human Pegivirus-1 and Sequence Variability of Its E2 Glycoprotein Estimated from Screening Donors of Fetal Stem Cell-Containing Material. Virol. J. 2017, 14, 167. [Google Scholar] [CrossRef] [Green Version]




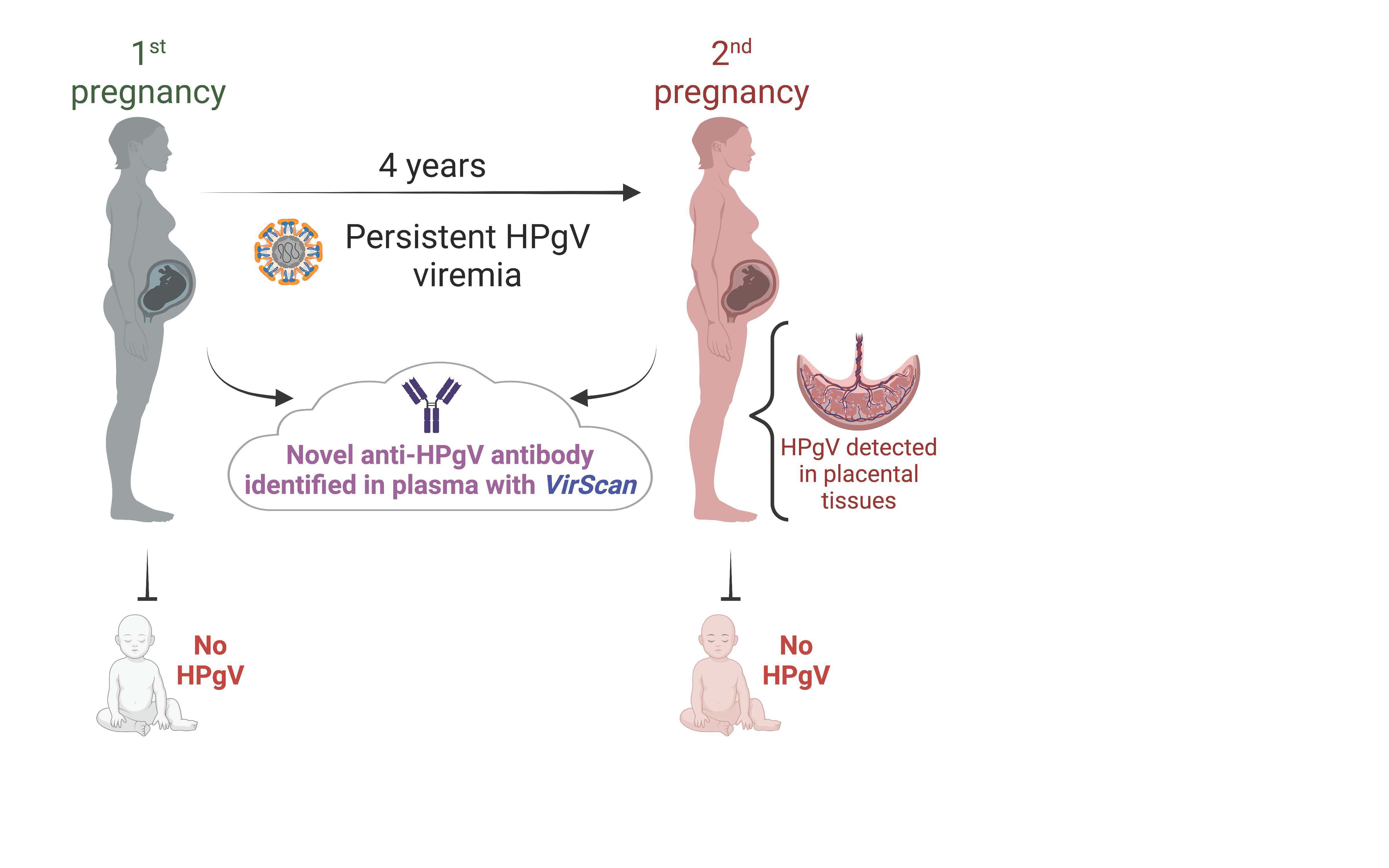{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pregnancy | Maternal Plasma | PBL | Decidua | IVTP | Cord Blood | Amnion | Chorion |
|---|---|---|---|---|---|---|---|
| 1st | ++ | +/− | − | ||||
| 2nd | ++ | +/− | ++ | ++ | − | +/− | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garand, M.; Huang, S.S.Y.; Goessling, L.S.; Santillan, D.A.; Santillan, M.K.; Brar, A.; Wylie, T.N.; Wylie, K.M.; Eghtesady, P. A Case of Persistent Human Pegivirus Infection in Two Separate Pregnancies of a Woman. Microorganisms 2022, 10, 1925. https://doi.org/10.3390/microorganisms10101925
Garand M, Huang SSY, Goessling LS, Santillan DA, Santillan MK, Brar A, Wylie TN, Wylie KM, Eghtesady P. A Case of Persistent Human Pegivirus Infection in Two Separate Pregnancies of a Woman. Microorganisms. 2022; 10(10):1925. https://doi.org/10.3390/microorganisms10101925
Chicago/Turabian StyleGarand, Mathieu, Susie S. Y. Huang, Lisa S. Goessling, Donna A. Santillan, Mark K. Santillan, Anoop Brar, Todd N. Wylie, Kristine M. Wylie, and Pirooz Eghtesady. 2022. "A Case of Persistent Human Pegivirus Infection in Two Separate Pregnancies of a Woman" Microorganisms 10, no. 10: 1925. https://doi.org/10.3390/microorganisms10101925