
Colistin Dependency among Colistin-Heteroresistant Acinetobacter baumannii Isolates

Abstract
:1. Introduction
2. Materials and Methods
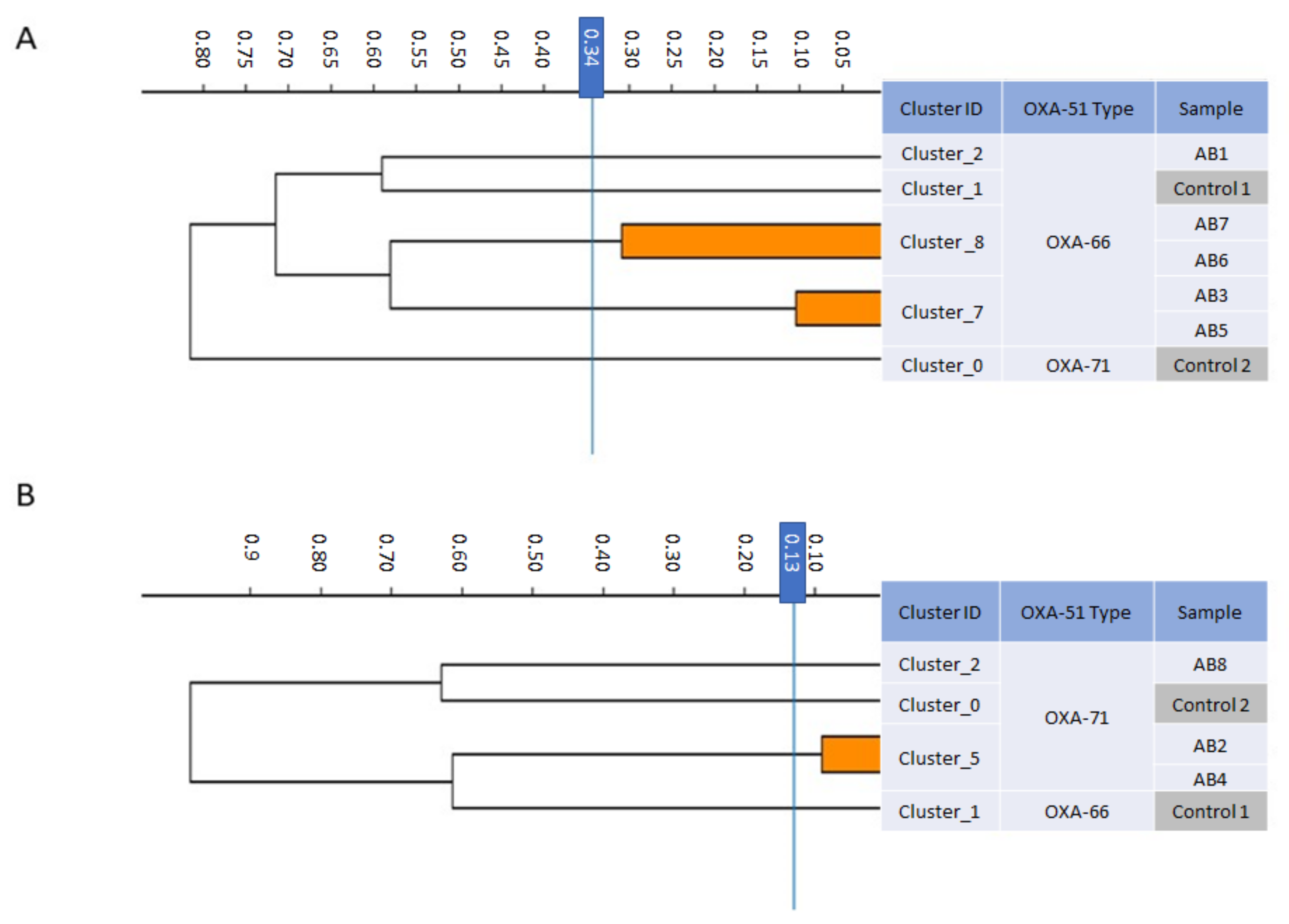
2.1. Description of the Isolate
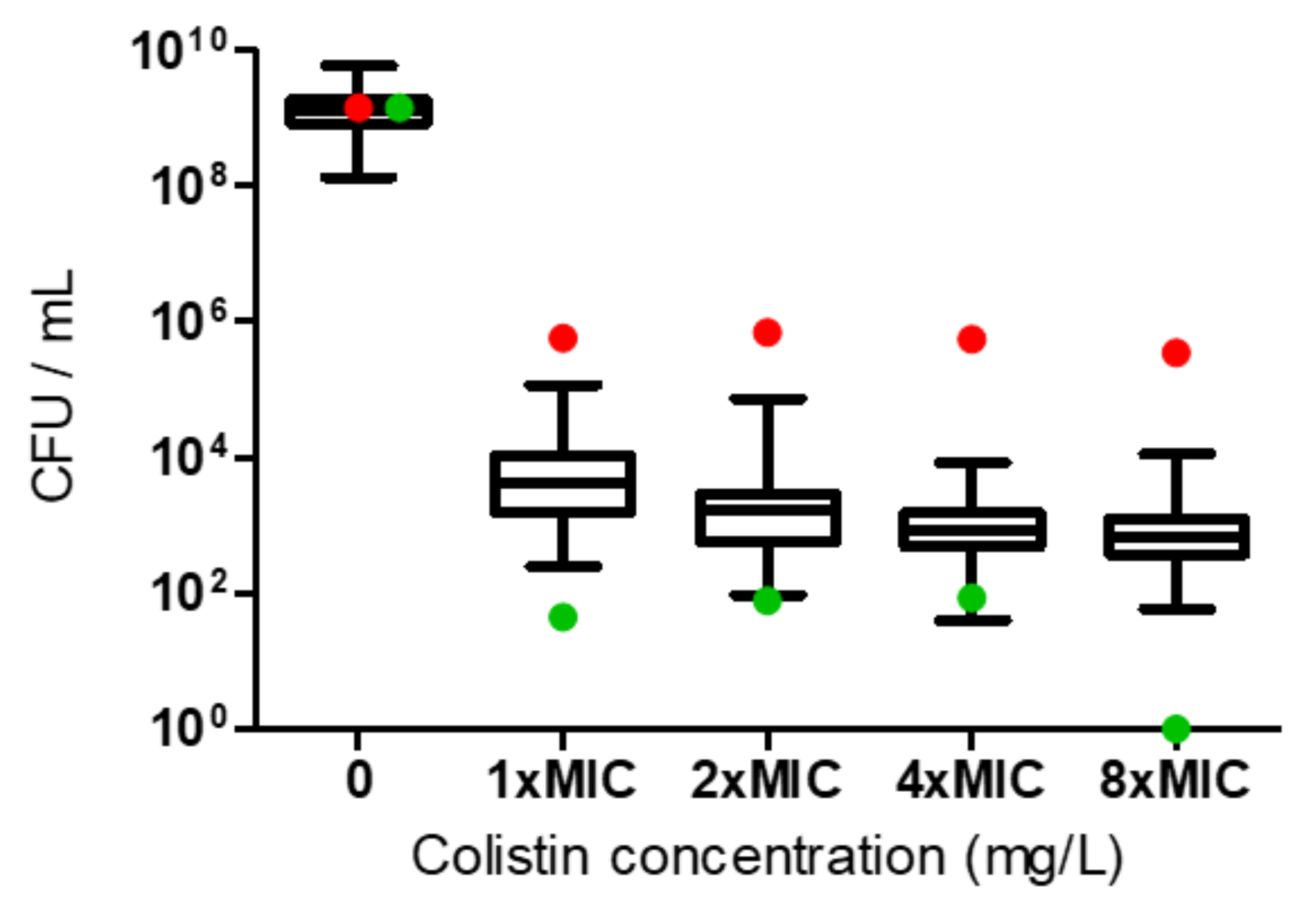
2.2. Determination of Colistin HR
2.3. Broth Microdilution (BMD)
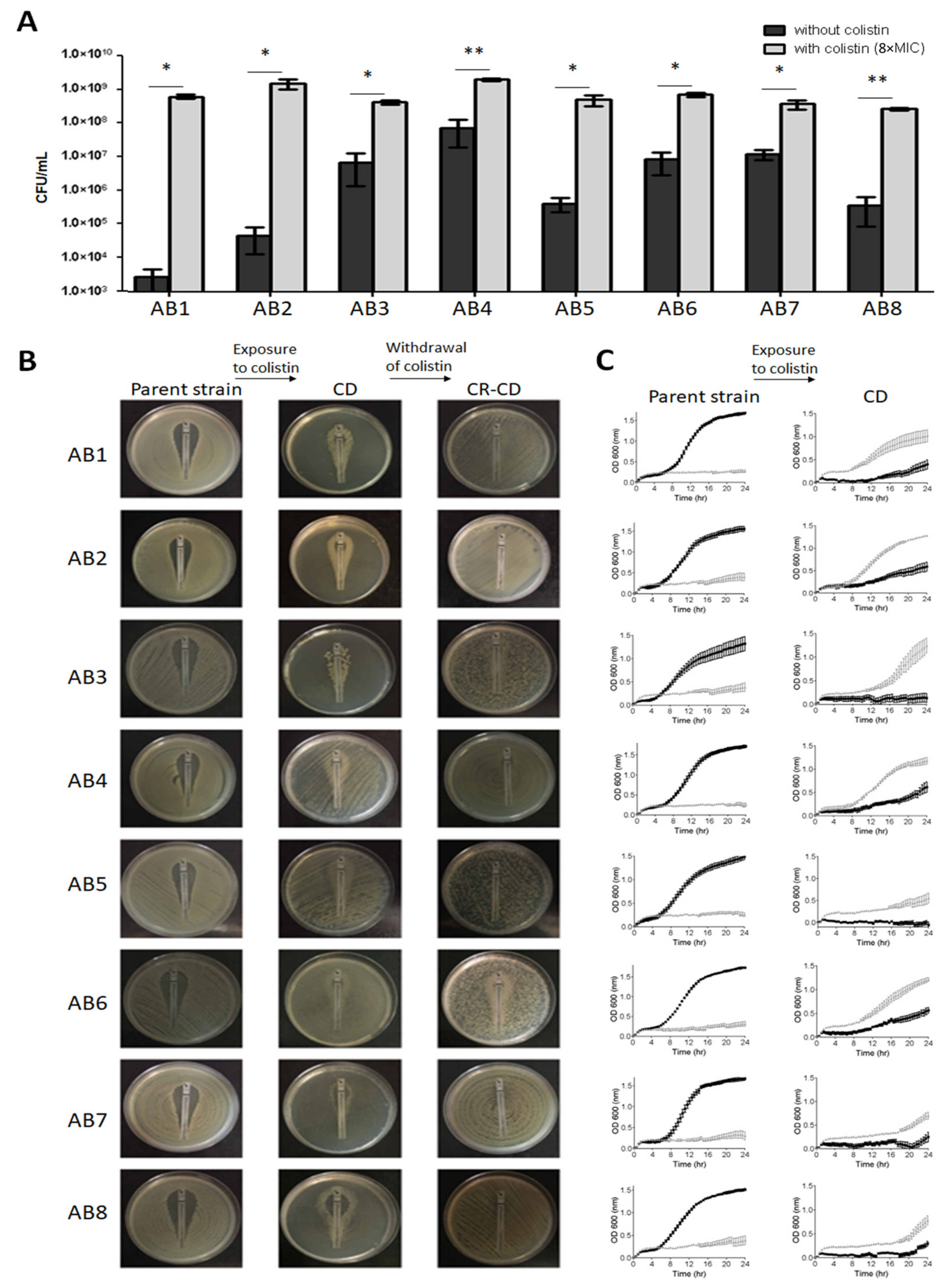
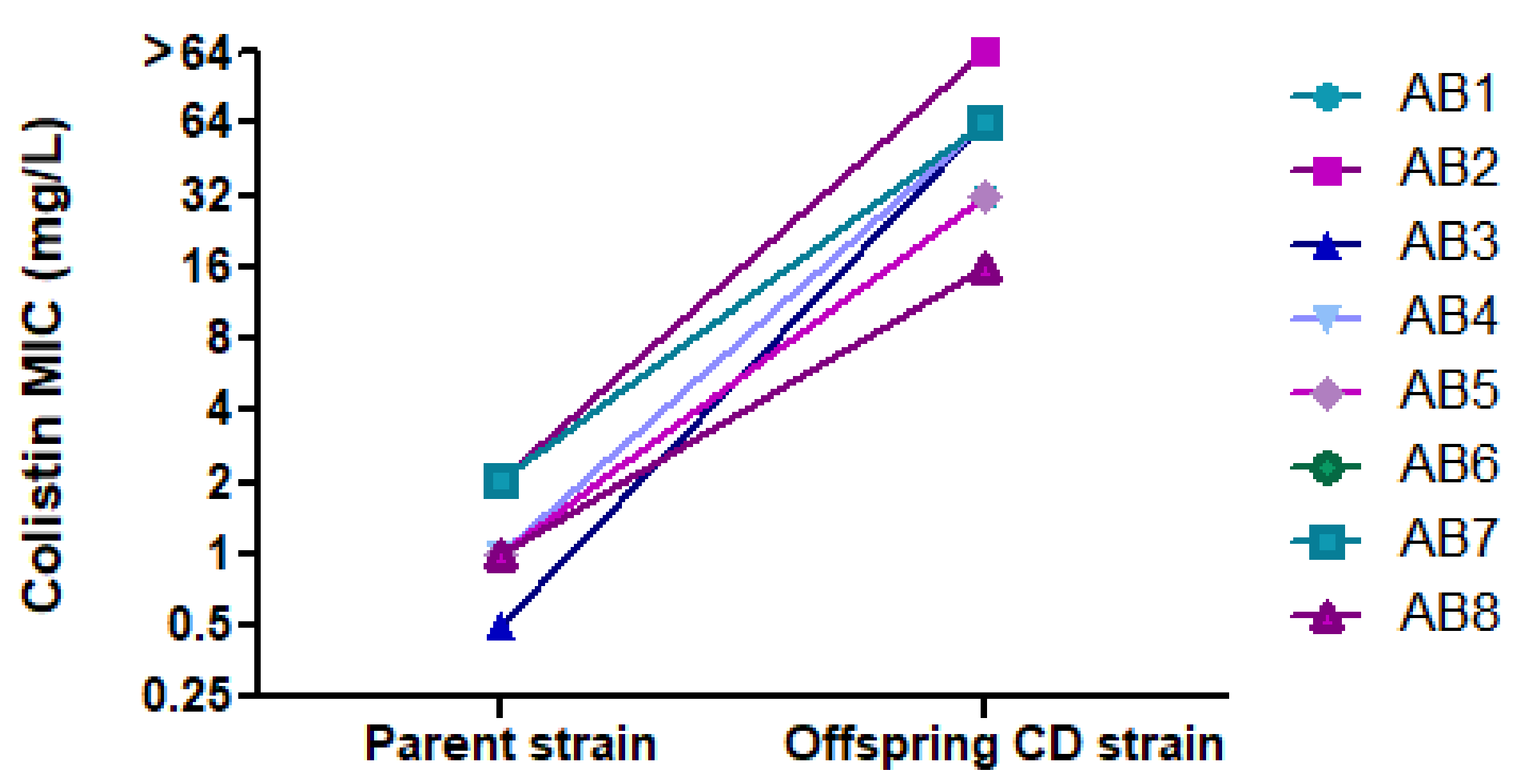
2.4. Exposure to Colistin and Determination of CD
2.5. Withdrawal of Colistin and Emergence of Overt Resistance
2.6. Colistin Etest
2.7. Growth Curves
2.8. Disk Diffusion
2.9. FT-IR Spectroscopy
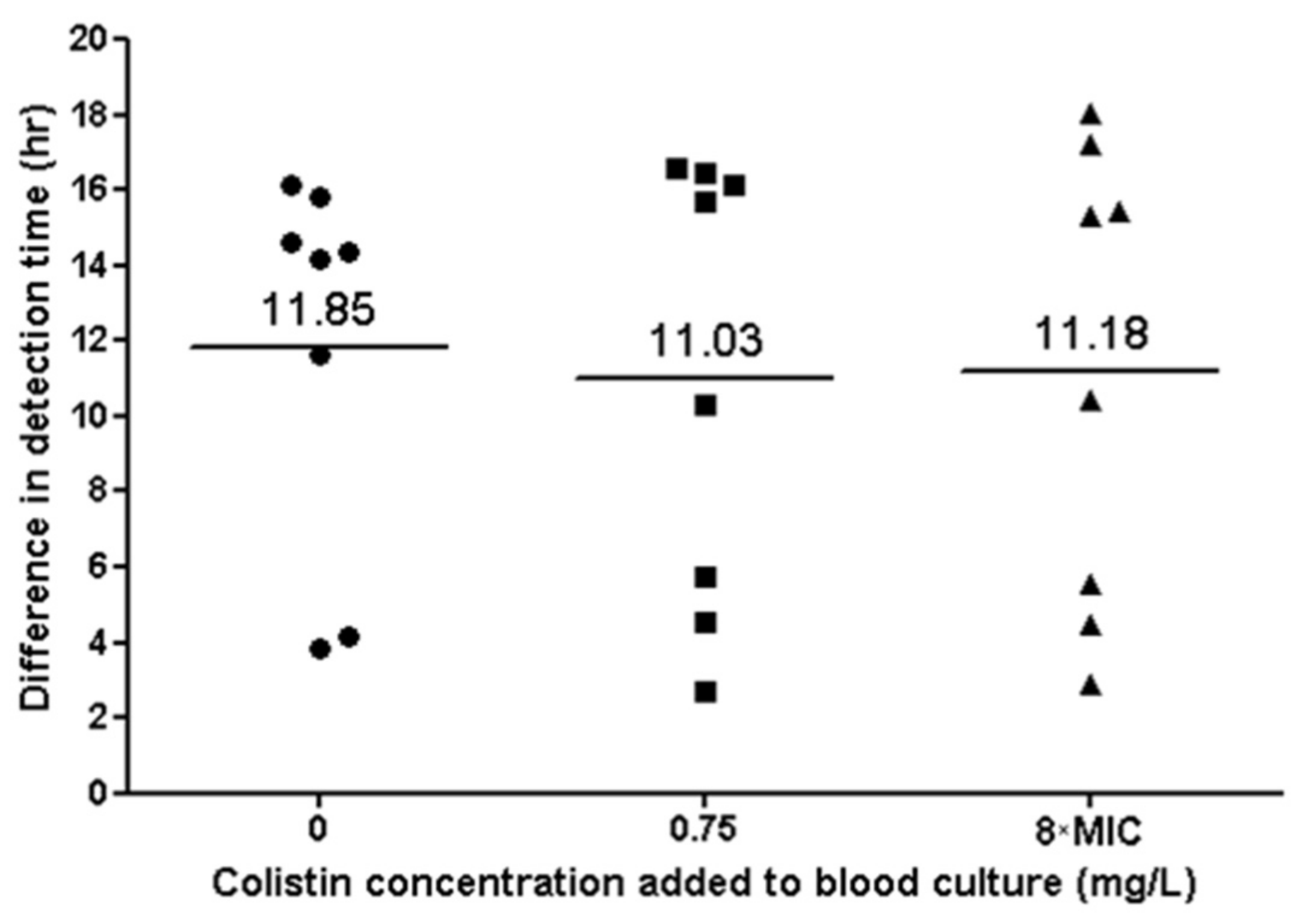
2.10. Blood Cultures
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fraimow, H.S.; Jungkind, D.L.; Lander, D.W.; Delso, D.R.; Dean, J.L. Urinary tract infection with an Enterococcus faecalis isolate that requires vancomycin for growth. Ann. Intern. Med. 1994, 121, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Hawley, J.S.; Murray, C.K.; Jorgensen, J.H. Development of colistin-dependent Acinetobacter baumannii-Acinetobacter calcoaceticus complex. Antimicrob. Agents Chemother. 2007, 51, 4529–4530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Global Priority List of Antibiotic-Resistant Batceria to Guide Research, Discovery, and Development of New Antibiotics. 2017. Available online: https://www.who.int/medicines/publications/WHO-PPL-Short_Summary_25Feb-ET_NM_WHO.pdf (accessed on 14 November 2021).
- Lee, J.Y.; Chung, E.S.; Ko, K.S. Transition of colistin dependence into colistin resistance in Acinetobacter baumannii. Sci. Rep. 2017, 7, 14216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, Y.K.; Lee, J.Y.; Wi, Y.M.; Ko, K.S. High rate of colistin dependence in Acinetobacter baumannii. J. Antimicrob. Chemother. 2016, 71, 2346–2348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, B.A.; Hamouda, A.; Towner, K.J.; Amyes, S.G.B. OXA-51-like β-lactamases and their association with particular epidemic lineages of Acinetobacter baumannii. Clin. Microbiol. Infect. 2008, 14, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Frenk, S.; Rakovitsky, N.; Kon, H.; Rov, R.; Abramov, S.; Lurie-Weinberger, M.N.; Schwartz, D.; Pinco, E.; Lellouche, J.; Carmeli, Y. OXA-900, a novel OXA sub-family carbapenemase identified in Citrobacter freundii, evades detection by commercial molecular diagnostics tests. Microorganisms 2021, 9, 1898. [Google Scholar] [CrossRef] [PubMed]
- Sherman, E.X.; Wozniak, J.E.; Weiss, D.S. Methods to evaluate colistin heteroresistance in Acinetobacter baumannii. Methods Mol. Biol. 2019, 1946, 39. [Google Scholar] [CrossRef]
- International Organization for Standardization (ISO) 20776-1:2019. Susceptibility Testing of Infectious Agents and Evaluation of Performance of Antimicrobial Susceptibility Test Devices—Part 1: Broth Micro-Dilution Reference Method for Testing the In Vitro Activity of Antimicrobial Agents against Rapidly Growing Aerobic Bacteria Involved in Infectious Diseases; ISO: Geneva, Switzerland, 2019. [Google Scholar]
- European Committee on Antimicrobial Susceptibility Testing (EUCAST). Recommendations for MIC Determination of Colistin (Polymyxin E) as Recommended by the Joint CLSI-EUCAST Polymyxin Breakpoints Working Group. 2016. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/General_documents/Recommendations_for_MIC_determination_of_colistin_March_2016.pdf (accessed on 14 November 2021).
- European Committee on Antimicrobial Susceptibility Testing (EUCAST). EUCAST Reading Guide for Broth Microdilution, Version 2.0. 2020. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Disk_test_documents/2020_manuals/Reading_guide_BMD_v_2.0_2020.pdf (accessed on 14 November 2021).
- European Committee on Antimicrobial Susceptibility Testing (EUCAST). Breakpoint Tables for Interpretation of MICs and Zone Diameters. 2021. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/v_11.0_Breakpoint_Tables.pdf (accessed on 14 November 2021).
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, M100, 30th ed.; CLSI: Wayne, PA, USA, 2020. [Google Scholar]
- García-Quintanilla, M.; Carretero-Ledesma, M.; Moreno-Martínez, P.; Martín-Peña, R.; Pachón, J.; McConnell, M.J. Lipopolysaccharide loss produces partial colistin dependence and collateral sensitivity to azithromycin, rifampicin and vancomycin in Acinetobacter baumannii. Int. J. Antimicrob. Agents 2015, 46, 696–702. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Fifth Informational Supplement. CLSI Document M100-S25; CLSI: Wayne, PA, USA, 2015. [Google Scholar]
- Rakovitsky, N.; Frenk, S.; Kon, H.; Schwartz, D.; Temkin, E.; Solter, E.; Paikin, S.; Cohen, R.; Schwaber, M.J.; Carmeli, Y.; et al. Fourier transform infrared spectroscopy is a new option for outbreak investigation: A retrospective analysis of an extended-spectrum-beta-lactamase-producing Klebsiella pneumoniae outbreak in a neonatal intensive care unit. J. Clin. Microbiol. 2020, 58, e00098-20. [Google Scholar] [CrossRef] [PubMed]
- Ko, K.S.; Choi, Y.; Lee, J.Y. Old drug, new findings: Colistin resistance and dependence of Acinetobacter baumannii. Precis. Future Med. 2017, 1, 159–167. [Google Scholar] [CrossRef] [Green Version]
- Marchaim, D.; Navon-Venezia, S.; Leavitt, A.; Chmelnitsky, I.; Schwaber, M.J.; Carmeli, Y. Molecular and epidemiologic study of polyclonal outbreaks of multidrug-resistant Acinetobacter baumannii infection in an Israeli hospital. Infect. Control Hosp. Epidemiol. 2007, 28, 945–950. [Google Scholar] [CrossRef] [PubMed]
- Frenk, S.; Temkin, E.; Lurie-Weinberger, M.N.; Keren-Paz, A.; Rov, R.; Rakovitsky, N.; Wullfhart, L.; Nutman, A.; Daikos, G.L.; Skiada, A.; et al. Large scale whole genome sequencing of carbapenem-resistant Acinetobacter baumannii isolates reveals patterns of dissemination of ST clades associated with antibiotic resistance. J. Antimicrob. Chemother. accepted.
- Mammina, C.; Bonura, C.; Aleo, A.; Calà, C.; Caputo, G.; Cataldo, M.C.; Di Benedetto, A.; Distefano, S.; Fasciana, T.; Labisi, M.; et al. Characterization of Acinetobacter baumannii from intensive care units and home care patients in Palermo, Italy. Clin. Microbiol. Infect. 2011, 17, E12–E15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hojabri, Z.; Pajand, O.; Bonura, C.; Aleo, A.; Giammanco, A.; Mammina, C. Molecular epidemiology of Acinetobacter baumannii in Iran: Endemic and epidemic spread of multiresistant isolates. J. Antimicrob. Chemother. 2014, 69, 2383–2387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flayhart, D.; Borek, A.P.; Wakefield, T.; Dick, J.; Carroll, K.C. Comparison of BACTEC PLUS blood culture media to BacT/Alert FA blood culture media for detection of bacterial pathogens in samples containing therapeutic levels of antibiotics. J. Clin. Microbiol. 2007, 45, 816–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, K.V.; Turner, N.N.; Lancaster, D.P.; Shah, A.R.; Chandler, L.J.; Friedman, D.F.; Blecker-Shelly, D.L. Superior sensitivity and decreased time to detection with the bactec peds Plus/F system compared to the BacT/Alert pediatric FAN blood culture system. J. Clin. Microbiol. 2013, 51, 4083–4086. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CD vs. Parent Strain | With Colistin vs. Without | |||
|---|---|---|---|---|
| Growth in Presence of Colistin | Growth in Absence of Colistin | Growth of Parent Strain | Growth of CD Strain | |
| Growth Rate (hr−1) | 0.06 * | −0.11 * | −0.13 ** | 0.04 * |
| Maximal Growth (lnOD) | 0.67 * | −1.24 * | −1.25 ** | 0.65 * |
| Lag time (hr) | −13.41 * | 11.05 * | 16.58 ** | −7.89 * |
| Meropenem (MEM) | Imipenem (IMP) | Ertapenem (ETP) | Ceftazidime (CAZ) | Ceftriaxone (CRO) | Sulfamethoxazole Trimethoprim (SXT) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P | CD | CR-CD | P | CD | CR-CD | P | CD | CR-CD | P | CD | CR-CD | P | CD | CR-CD | P | CD | CR-CD | |
| AB1 | 0 | 39 | 35 | 0 | 35 | 33 | 0 | 31 | 33 | 0 | 34 | 24 | 0 | 35 | 21 | 18 | 36 | 31 |
| AB2 | 0 | 41 | 41 | 10 | 36 | 39 | 0 | 37 | 34 | 0 | 27 | 26 | 0 | 28 | 22 | 14 | 30 | 32 |
| AB3 | 9 | 37 | 38 | 10 | 35 | 36 | 0 | 33 | 34 | 0 | 40 | 30 | 0 | 35 | 34 | 0 | 0 | 0 |
| AB4 | 0 | 33 | 37 | 0 | 31 | 23 | 0 | 31 | 26 | 0 | 29 | 23 | 0 | 31 | 20 | 0 | 34 | 22 |
| AB5 | 8 | 36 | 34 | 9 | 35 | 33 | 0 | 33 | 30 | 0 | 30 | 30 | 0 | 31 | 30 | 0 | 0 | 0 |
| AB6 | 0 | 30 | 37 | 0 | 37 | 36 | 0 | 27 | 35 | 0 | 40 | 39 | 0 | 45 | 31 | 19 | 45 | 38 |
| AB7 | 0 | 40 | 11 | 0 | 38 | 11 | 0 | 40 | 0 | 0 | 36 | 0 | 0 | 36 | 0 | 16 | 34 | 0 |
| AB8 | 11 | 45 | 13 | 0 | 45 | 11 | 0 | 45 | 0 | 11 | >45 | 0 | 0 | 45 | 0 | 0 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kon, H.; Hameir, A.; Temkin, E.; Keren-Paz, A.; Schwartz, D.; Schechner, V.; Carmeli, Y. Colistin Dependency among Colistin-Heteroresistant Acinetobacter baumannii Isolates. Microorganisms 2022, 10, 58. https://doi.org/10.3390/microorganisms10010058
Kon H, Hameir A, Temkin E, Keren-Paz A, Schwartz D, Schechner V, Carmeli Y. Colistin Dependency among Colistin-Heteroresistant Acinetobacter baumannii Isolates. Microorganisms. 2022; 10(1):58. https://doi.org/10.3390/microorganisms10010058
Chicago/Turabian StyleKon, Hadas, Amichay Hameir, Elizabeth Temkin, Alona Keren-Paz, David Schwartz, Vered Schechner, and Yehuda Carmeli. 2022. "Colistin Dependency among Colistin-Heteroresistant Acinetobacter baumannii Isolates" Microorganisms 10, no. 1: 58. https://doi.org/10.3390/microorganisms10010058