
Talaromyces–Insect Relationships



1
Research Centre for Olive, Fruit and Citrus Crops, Council for Agricultural Research and Economics, 81100 Caserta, Italy
2
Department of Agricultural Sciences, University of Naples Federico II, 80055 Portici, Italy
*
Author to whom correspondence should be addressed.
Microorganisms 2022, 10(1), 45; https://doi.org/10.3390/microorganisms10010045
Submission received: 19 November 2021
/
Revised: 21 December 2021
/
Accepted: 23 December 2021
/
Published: 26 December 2021
(This article belongs to the Special Issue Ecological and Molecular Interactions between Insects and Fungi)
Abstract
:Facing the urgent need to reduce the input of agrochemicals, in recent years, the ecological relationships between plants and their associated microorganisms have been increasingly considered as an essential tool for improving crop production. New findings and data have been accumulated showing that the application of fungi can go beyond the specific role that has been traditionally assigned to the species, employed in integrated pest management as entomopathogens or mycoparasites, and that strains combining both aptitudes can be identified and possibly used as multipurpose biocontrol agents. Mainly considered for their antagonistic relationships with plant pathogenic fungi, species in the genus Talaromyces have been more and more widely reported as insect associates in investigations carried out in various agricultural and non-agricultural contexts. Out of a total of over 170 species currently accepted in this genus, so far, 27 have been found to have an association with insects from 9 orders, with an evident increasing trend. The nature of their mutualistic and antagonistic relationships with insects, and their ability to synthesize bioactive compounds possibly involved in the expression of the latter kind of interactions, are analyzed in this paper with reference to the ecological impact and applicative perspectives in crop protection.
1. Introduction
Traditionally, the role of fungi in the contrasting biological adversities of plants has been considered with reference to their assignment to functional categories, such as ‘mycoparasites’, ‘entomopathogens’, ‘nematophagous fungi’, etc. More recently, the diffusion of the holistic approach considering the ecological interrelations among plant-associated organisms has impaired such a rigid distinction. Indeed, the recurring evidence that many fungi can play several mutualistic roles stimulates a reappraisal of their effects on plant health, as well as of the way they can be employed in the integrated management of pests and pathogens of crops [1]. A number of widespread entomopathogens, such as Beauveria bassiana and Lecanicillium/Akanthomyces spp., have also qualified as being able to perform antagonism against many plant pathogens, so that they can be employed as dual or multipurpose biocontrol agents [2,3,4]. Likewise, Trichoderma spp., until recently essentially regarded as fungal antagonists and mycoparasites, have also started to be considered for the role they are able to play against several pests [5,6].
Current evidence concerning the capacity of Talaromyces, another Ascomycetes genus including several species reported as antagonists of plant pathogens [7,8,9], to directly interact with insects is examined in this paper in view of a more comprehensive consideration of their role in crop protection and possible applications in the biocontrol of these key pests.
2. Occurrence of Talaromyces spp. as Insect Associates
Limited insights have been published on the association of Talaromyces spp. (Eurotiales, Trichocomaceae) with insects, depending on their traditional categorization in the guild of fungal antagonists. As well as the increasing awareness of their ability to play multiple ecological roles, the recent accumulation of data on these Ascomycetes is due to the introduction of molecular tools in fungal taxonomy. In fact, until the enunciation of the principle ‘one fungus, one name’ about 10 years ago, the genus name Talaromyces was basically used for symmetrically biverticillate Penicillia producing a perfect stage in axenic culture. Conversely, species for which no teleomorph could be observed were classified in the polyphyletic genus Penicillium. Phylogenetic analyses based on molecular markers pointed out this inconsistency, so that species in the Penicillium subgenus Biverticillium were officially separated and formally assigned to Talaromyces [10]. At the same time, the spread of DNA sequencing in the identification of fungi collected from any ecological context has enabled their correct (or less approximate) classification, and provided a remarkable boost to investigations aiming at a general description of the mycobiomes of plants and other organisms.
The references provided in Table 1, which do not include cases of mycophagy, confirm the increasing trend in reports of Talaromyces spp. in associations with insects. In fact, the first documented finding dates back to 1990, when isolates of Talaromyces funiculosus (at that time known as Penicillium funiculosum) were recovered from larvae of Aedes cantans and Aedes communis (Diptera, Culicidae) collected in the Kiev region, Ukraine [11]. Three more reports were published in the 1990s, concerning the finding of Talaromyces diversus (identified as Penicillium diversum) and Talaromyces trachyspermus from meconia in nests of the paper wasp Polistes hebraeus (= P. olivaceus) (Hymenoptera, Vespidae) in La Reunion island [12], Talaromyces purpureogenus (identified as Penicillium purpurogenum) on several species of mosquitoes in Brazil [13] and Talaromyces duclauxii (as Penicillium duclauxii) from dead larvae and adults of the cave cricket Troglophilus neglectus (Orthoptera, Rhaphidophoridae) in Slovenia [14]. All other findings have been reported in papers published after 2004; after having been quite infrequent until 2015, the number of articles has more than doubled in the last five years.
Out of a total of over 170 species accepted in the last genus revision [56], just 27 have been reported in associations with insects so far. However, some of these findings refer to new or infrequent species, supporting the expectation that exploring this symbiotic context can be fruitful in terms of unveiling hidden biodiversity. In fact, Talaromyces cecidicola was originally described as Penicillium cecidicola based on isolates from galls formed by unidentified cynipid wasps on scrub oaks (Quercus pacifica) in the western United States [18]. This species is sister to Talaromyces dendriticus (originally described as Penicillium dendriticum), which was also isolated from galls produced by an unidentified insect on Eucalyptus leaves in Australia [18]. Other gallicolous species, Talaromyces erythromellis and Talaromyces pseudostromaticus (again both originally described as Penicillium species), were identified from galls of another cynipid wasp (Diplolepis rosae) on Rosa sitchensis in Canada [18].
Besides gallicolous cynipids, interactions with Hymenoptera have resulted from investigations concerning nests built by some eusocial species of these insects. These include the above-mentioned finding of T. diversus and T. trachyspermus from nests of P. hebraeus [12], Talaromyces variabilis (currently synonymized with Talaromyces wortmannii) from nests of another paper wasp (Ropalidia marginata) [54] and three new species (Talaromyces brasiliensis, Talaromyces pigmentosus and Talaromyces mycothecae), which have been described based on isolates recovered from nests of the Brazilian stingless bee Melipona scutellaris (Hymenoptera, Apidae), along with the known species Talaromyces scorteus and T. wortmannii [17]. Other records come from the nests of several species of leaf-cutting ants in Texas (USA) [33], Brazil [50] and Argentina [37]. However, at least in the case of the anthill gardens, the occurrence of three common species (Talaromyces rugulosus, Talaromyces verruculosus and T. purpureogenus) does not seem to be related to any specific function, rather depending on saprophytism or mycoparasitism.
Uncircumstantial associations with other well-known eusocial insects, the termites (Blattodea, Termitidae and Rhinotermitidae), have been documented for a few isolates: Talaromyces flavus from the coarse debris of wood infested by Reticulitermes sp. in Mississippi (USA) [24], Talaromyces spp. from the nests of Coptotermes formosanus under experimental rearing conditions in Florida (USA) [40] and combs of the fungus-growing species Macrotermes carbonarius in Vietnam [39], while Talaromyces stollii was reported from the combs of Macrotermes barneyi in China [47].
All other findings concern living or dead insects belonging to Coleoptera, Diptera, Hemiptera, Hymenoptera, Lepidoptera, Orthoptera, Thysanoptera and Trichoptera. Each of the last three orders has just a single occasional citation concerning the above-mentioned strains of T. duclauxii from T. neglectus [14], a strain of T. verruculosus from an unidentified thrips in Thailand [49] and a strain of T. purpureogenus from the gut of a larva of an aquatic shredder (Triplectides sp.) (Trichoptera, Leptoceridae) in Amazonia (Brazil). The latter strain displayed cellulolytic properties, which can be inferred to possibly contribute to the digestion of plant matter [36]. Conversely, more systematic and abundant occurrences are those of Talaromyces spp. in the abdomen fat body and gut of the brown planthopper (Nilaparvata lugens) (Hemiptera, Delphacidae) in China [45,46], which deserves further investigation in view of the assessment of eventual functional relationships. Moreover, the occurrence of Talaromyces in low relative abundance resulted in the analysis of the endomycobiome of another planthopper species, Delphacodes kuscheli [57].
A functional role based on cellulolytic properties could also be implicated in the association of Talaromyces spp. with xylicolous Coleoptera. This is the case of Talaromyces pulveris, which has been very recently described based on an isolate from bore dust of the deathwatch beetle (Xestobium rufovillosum, Anobiidae) in France [32], while other findings concern the gut of the ribbed pine borer (Rhagium inquisitor, Cerambycidae) collected in Tatarstan (Russia) [43] and of the ambrosia/bark beetles (Curculionidae, Platypodinae and Scolytinae). In particular, many cases are known for species in the latter group, starting from Polygraphus poligraphus and Ips spp. infesting coniferous forests in the Czech Republic where two species (T. rugulosus and T. variabilis) were represented in the core mycobiome, as well as Talaromyces minioluteus, which was only isolated from Ips sexdentatus [29]. In fact, other strains of Talaromyces atroroseus and Talaromyces pinophilus, respectively recovered from living and dead adults of the pistachio bark beetle (Chaetoptelius vestitus) in Tunisia, have been characterized with reference to cellulase and protease production [16]. More findings concern a strain of unidentified species from living adults of the pinhole borer (Platypus cylindrus) infesting oaks at the Astroni Nature Reserve in southern Italy (Nicoletti, unpublished), multiple isolates of Talaromyces radicus, T. variabilis and T. purpureogenus from Dendroctonus spp. associated with coniferous plants in France [34], T. trachyspermus from adults of Ips typographus trapped in Norway spruce stands in northwestern Italy [48], T. minioluteus and T. purpureogenus from adults of Ips acuminatus collected in the Ukraine [28], and T. verruculosus from the mycangia of adults of Xylosandrus crassiusculus from specimens collected in Florida (USA) and China [51]. The epibiotic association with three Tomicus species (T. brevipilosus, T. minor and T. yunnanensis) of multiple Talaromyces spp. anonymously identified as ‘OTUs’ has been reported from the Yunnan province of the latter country [44]. Moreover, in a similar study carried out in Mexico, a high prevalence of T. purpureogenus was observed in association with Xyleborus affinis and Xyleborus bispinatus [35]. Finally, the occurrence of T. rugulosus was detected in gardens of Xyleborinus saxesenii under laboratory rearing conditions [38].
Isolations from the gut were also obtained in the case of haematophagous species such as the kissing bug Triatoma infestans (Hemiptera, Reduviidae) from Argentina [27] and the mosquito Aedes aegypti from Puerto Rico [41], both deserving further investigation. In fact, the discovery of three species (Talaromyces ruber, Talaromyces aff. helicus and T. purpureogenus) on T. infestans is indicative of a possible natural spread of Talaromyces in association with these noxious bugs. Moreover, a strain of unidentified species from A. aegypti was found to facilitate the infection of mosquitoes with the dengue virus (DENV), thereby possibly enhancing virus transmission. This modulation is related to the down-regulation of digestive enzyme genes and trypsin activity upon exposure to factors secreted by the fungus in the insect’s gut. The experimental finding that the fungus can be acquired by mosquitoes through sugar feeding, and that it is able to successfully colonize the midgut for a period of at least 25 days, is indicative of a functional relationship that requires better elucidation [41]. Uncircumstantial relationships with a blood sucking insect have also been considered with regard to isolates of T. variabilis from the larvae of Simulium goeldii (Diptera, Simuliidae) in Amazonia (Brazil) [53].
Other findings are seemingly occasional and do not represent indications of possible functional relevance. This is the case of two Egyptian reports concerning T. funiculosus from dead larvae and pupae of the vegetable leafminer (Liriomyza sativae) (Diptera, Agromyzidae) [26] and T. pinophilus from an unspecified stage of the red palm weevil (Rhynchophorus ferrugineus) (Coleoptera, Curculionidae) [30], as well as of Talaromyces sp. from an unidentified mud dauber wasp in Australia [42], Talaromyces versatilis from dead individuals of the giant honey bee (Apis dorsata) (Hymenoptera, Apidae) in Sri Lanka [52] and T. variabilis from adults of Diabrotica sp. (Coleoptera, Chrysomelidae) and Neomyopites sp. (Diptera, Tephritidae) collected on the subshrub Espeletia pycnophylla in Colombia [55]. In the course of an investigation carried out in Lebanon, the species Talaromyces amestolkiae exhibited the capacity to adapt to insect hosts from different orders, with several strains recovered from corpses of Capnodis tenebrionis (Coleoptera, Buprestidae), Culex sp. (Diptera, Culicidae) and unidentified species of Pyrrhocoridae (Hemiptera) and Pyralidae (Lepidoptera) [15]. Concerning the last order, two strains of T. flavus and several strains of T. pinophilus were recovered from larvae and pupae of the antophagous and carpophagous generations of the olive moth (P. oleae) (Plutellidae) in Portugal [23]. Moreover, strains of T. trachyspermus and T. flavus were recovered in China from larvae of the honeycomb moth (Galleria mellonella) (Pyralidae) used as bait [21,22], and the latter species was found in frass collected in tunnels of the maize stalk borer (Busseola fusca) (Noctuidae) in South Africa [25]. Finally, T. flavus was reported from mines of the long-legged flies Thrypticus truncatus and Thrypticus sagittatus (Diptera, Dolichopodidae) on water hyacinth (E. crassipes) in Argentina [20].
3. Experimental Evidence of Anti-Insectan Effects
It is quite obvious that the isolation of fungi from insects and/or their nests merely represents an indication of ecological interactions that do not necessarily have adaptive or functional implications. However, the intent to exploit anti-insectan properties in biocontrol has stimulated investigations aimed at assessing if the observed associations may eventually result in detrimental effects on several insect pests.
Experimental assays were carried out with a strain of T. flavus isolated from larval breeding sites of the mosquito Anopheles albimanus (Diptera, Culicidae) in the coastal plain of Chiapas (Mexico), which induced low mortality (16.6%) on nymphs of the kissing bug (Triatoma dimidiata). Nymphs died after a long period following inoculation (21 days), and the fungus exhibited no sporulation; however, when administered to adults, mortality rose to 75%, with a lower interval (16 days). This difference possibly derived from the loss of conidia, which may have occurred during molting after the topical inoculation of nymphs [58].
The degree of susceptibility to the anti-insectan effects of Talaromyces may depend on the insect species and may be affected by the experimental conditions. In preliminary tests, high concentrations of conidia of strains of T. funiculosus killed second instar larvae of the mosquitoes Culex pipiens ‘form molestus’ and A. aegypti [11]. Conidial suspension of another strain of this species affected vitality and oviposition in assays carried out against L. sativae on tomato [26]. Strains of T. amestolkiae recovered from the corpses of several insects were found to only slightly affect the survival of adults of the vinegar fly (Drosophila melanogaster) (Diptera, Drosophilidae) and the tiger mosquito (Aedes albopictus) following inoculation under laboratory conditions [15]. A strain of T. verruculosus isolated from an unidentified thrips caused mortality at a low level (13–23%) in bioassays carried out on thrips (Ceratothripoides claratris) (Thysanoptera, Thripidae), mealybugs (Pseudococcus cryptus) (Hemiptera, Pseudococcidae) and whiteflies (Bemisia tabaci) (Hemiptera, Aleyrodidae) [49]. A low rate of mortality (about 25%) was also caused by another isolate of T. verruculosus from larva of Bactrocera oleae (Diptera, Tephritidae) in assays carried out on larvae of the Mediterranean flour moth (Ephestia kuehniella) (Lepidoptera, Pyralidae) [31]. Finally, in laboratory assays carried out on the black bean aphid (Aphis fabae) and the Russian wheat aphid (Diuraphis noxia), a strain of T. pinophilus recovered from R. ferrugineus caused 30% and 50% mortality, respectively [30].
Experimental evidence of anti-insectan effects was also achieved in investigations concerning Talaromyces strains obtained from other sources. This is the case of T. minioluteus found in the laboratory diet of the spotted wing drosophila (Drosophila suzukii), which affected flies’ development by extending the pupal stage by 16.22%, shortening adult longevity by 15.52% and reducing survival by 71.67% [59]. Moreover, a few strains recovered from soil in several locations in Indonesia yielded quite positive results. In particular, strains of Talaromyces sayulitensis from the rhizosphere of pineapple, corn and pepper were found to be able to infect and cause the mortality (16.67–46.67%) of cocoa bugs (Helopeltis sp.) (Hemiptera, Miridae) [60,61]. Moreover, a strain of T. diversus, isolated from soil cropped to mustard using mealworm larvae (Tenebrio molitor) (Coleoptera, Tenebrionidae) as bait, was found to possess insecticidal activity as assayed by both conidial suspension and culture filtrate against the cotton leafworm (Spodoptera litura) (Lepidoptera, Noctuidae) [19], while the use of a strain of T. pinophilus from a rubber tree plantation was proposed in the formulation of a bioinsecticide for the control of the coffee borer beetle (Hypothenemus hampei) (Coleoptera, Curculionidae, Scolytinae) [62]. Finally, a strain of T. verruculosus recovered from soil in West Bengal was reported for its notable entomopathogenic effects in laboratory assays carried out against the cotton aphid (Aphis gossypii) (Heteroptera, Aphididae). It must be noted that, probably due to species name similarity, this strain was incorrectly reported as Penicillium verrucosum, despite evidence concerning its morphology and ITS sequencing provided in the original report being clearly indicative of T. verruculosus [63]. In this respect, it is recommended that the authors make the necessary amendment to the data deposited in GenBank in order to avoid that further misleading identifications may occur in the future.
4. The Role of Secondary Metabolites
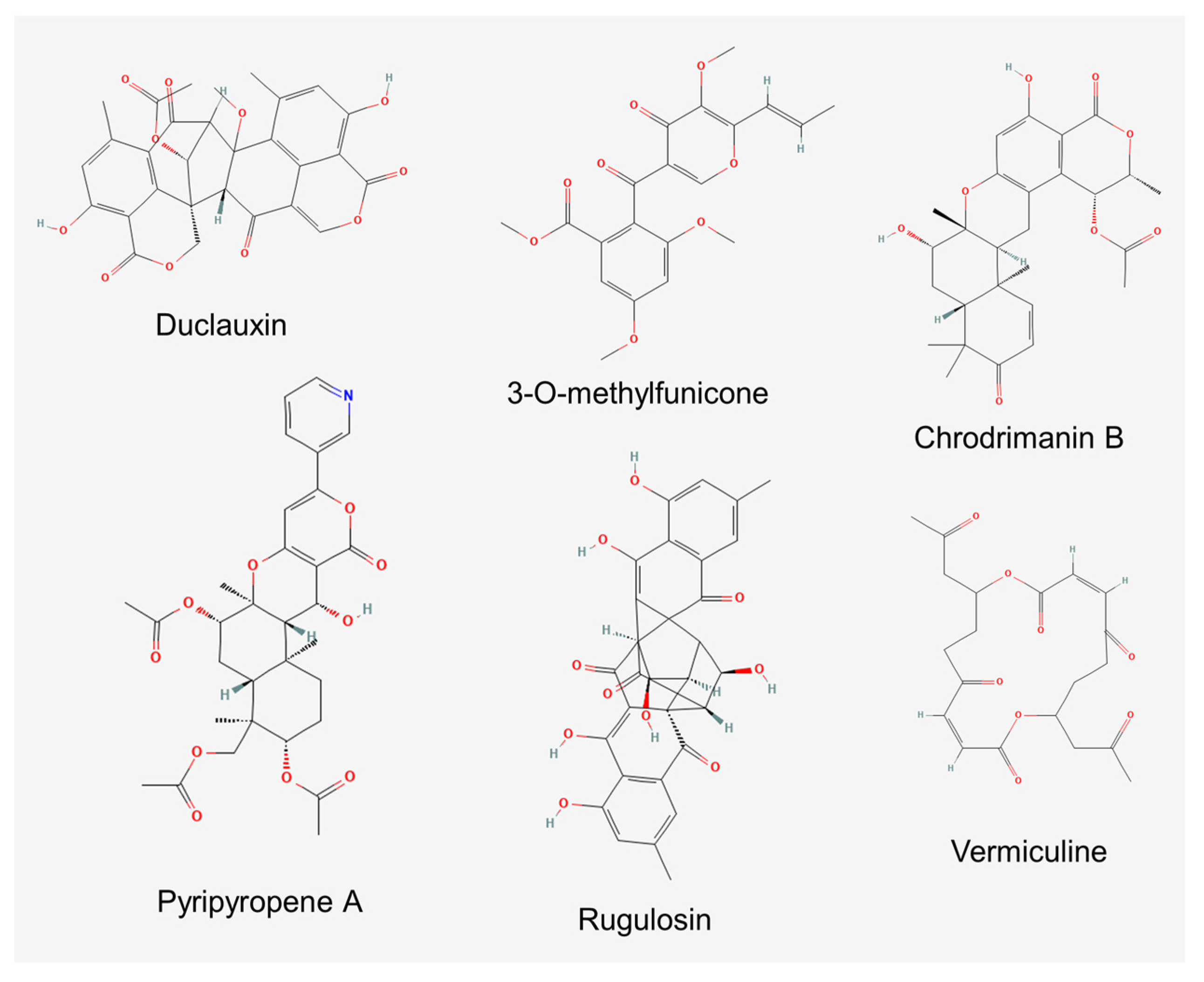
Despite the recognized relevance of bioactive secondary metabolites in shaping the relationships between fungi and arthropods [64], none of the above-mentioned studies addressed whether the observed anti-insectan properties are due to the release of these products by the insect-associated fungi. Indeed, Talaromyces species are renowned as producers of a wide array of bioactive compounds [65,66,67], and some clues that insect-associated strains may also represent a source of bioactive products have also arisen. This is shown in the case of the hymenopteran-derived strains of T. versatilis and Talaromyces sp. that display antibacterial properties in their culture extracts or purified compounds [52,68], and the three new species described from isolations from the nests of M. scutellaris. Among the latter, T. mycothecae produced the isocoumarin dimer antibiotic duclauxin (Figure 1) and some derivatives of the same, while T. brasiliensis and T. pigmentosus were found to produce several unknown secondary metabolites [17].
Conversely, several studies involving Talaromyces strains obtained from other sources have provided more circumstantial evidence that these fungi may produce anti-insectan compounds. In fact, dichloromethane and methanol extracts from cultures of a strain of T. funiculosus displayed various activities in assays carried out on insects; this includes aphicidal activity against the green peach aphid (Myzus persicae) (Hemiptera, Aphididae), ovicidal activity against the Colorado potato beetle (Leptinotarsa decemlineata) (Coleoptera, Chrysomelidae), adulticide activity against the large milkweed bug (Oncopeltus fasciatus) (Hemiptera, Lygaeidae) and juvenile hormone mimetic activity on the German cockroach Blattella germanica (Blattodea, Ectobiidae) [69]. Moreover, dichloromethane extracts obtained from the cultures of strains of T. funiculosus, T. purpureogenus and T. rugulosus from cereal grains displayed various degrees of toxicity in assays carried out on nymphs of O. fasciatus [70].
Similar evidence has also been obtained for some purified compounds (Figure 1). For instance, phlegmacin B1 from a soil strain of Talaromyces sp., a dimeric pre-anthraquinone possessing inhibitory properties towards chitinases of the Asian corn borer (Ostrinia furnacalis) (Lepidoptera, Crambidae), may impair larval development during molting [71]. Moreover, 3-O-methylfunicone produced by T. pinophilus, a benzo-γ-pyrone known for its notable antifungal and antitumor properties [72,73], displayed aphicidal effects as assayed on the pea aphid (Acyrthosiphon pisum) (Hemiptera, Aphididae) [74].
Anti-insectan properties have been investigated in more detail in the case of chrodrimanins, meroterpenoids produced by several Talaromyces spp. [75,76,77,78,79]. Chrodrimanins B (Figure 1), D, E and F exhibited insecticidal activity when added to the diet of third instar larvae of silkworm (Bombyx mori) (Lepidoptera, Bombycidae), with LD50 values of 10, 20, 10 and 50 µg/g, respectively [77,80]. Based on the observation that the treated larvae underwent paralysis, the bioactivity of chrodrimanin B was investigated using patch-clamp electrophysiology on ligand-gated ion channels of larval neurons. The compound was found to have no effect on membrane currents when tested at 1 μM. However, when delivered at the same concentration for 1 min prior to co-application with γ-aminobutyric acid (GABA), it completely blocked the GABA-induced current, also displaying minor actions on acetylcholine- and l-glutamate-induced currents. Moreover, chrodrimanins A, B and D were also tested on a wild-type isoform of the GABA receptor of silkworms and were found to be able to attenuate the peak current amplitude of the GABA response, with an IC50 of 1.66 nM. The order of the blocking potency of chrodrimanins (B > D > A) was in accordance with their reported insecticidal effects. Chrodrimanin B appears to be a selective blocker of insect GABA receptors since its activity on the human GABA receptor was approximately 103-fold lower [81].
Pyripyropene A (Figure 1) is another meroterpenoid reported as a secondary metabolite in Talaromyces [82]. This product displayed high activity against M. persicae, not only in laboratory assays at a concentration of 1.25 ppm but also in applications on cabbage plants through both foliar sprays and soil drenching. Structure-related bioactivity studies showed that the pyridine nucleus of this compound is an important pharmacophore since its replacement with a phenyl ring in the analogue phenylpyropene A caused the loss of insecticidal activity. Moreover, the effectiveness of derivatives lacking the acetyl ester groups was remarkably lower, suggesting the possibility of improving bioactivity by modifying these parts of the molecule [83]. The compound confirmed good activity in further assays carried out on A. gossypii, the mealybug Pseudococcus comstocki and two whiteflies (B. tabaci and Trialeurodes vaporariorum) (Hemiptera, Aleyrodidae), while it was inactive against the rice leaf bug (Trigonotylus caelestialium) (Hemiptera, Miridae), the planthoppers N. lugens and Nephotettix cincticeps (Hemiptera, Cicadellidae), the diamondback moth (Plutella xylostella) (Lepidoptera, Plutellidae), S. litura and the western flower thrips (Frankliniella occidentalis) (Thysanoptera, Thripidae) [84].
Besides evidence of its direct insecticidal effects, pyripyropene A has been considered as a model for the synthesis of molecules with improved bioactivities [85,86,87]. In this respect, it has also been characterized for acetyl-CoA acyl transferase inhibitory effects, which are indicative of its potential insecticidal properties [88]; however, the above-mentioned study by Horikoshi et al. [83] pointed out that this mechanism is not able to explain the observed bioactivity. On the other hand, the culture extracts of many fungi have been found to possess acetylcholinesterase (AchE) inhibitory properties, which is considered a primary target for insecticides [89]. The production of AchE inhibitors has also been reported for some Talaromyces strains [23,90], and some products have been purified and characterized for this bioactivity, such as talaromycesone A and talaroxanthenone [91].
Initially reported as a coloring matter produced by Penicillium rugulosum (= T. rugulosus) [92], rugulosin (Figure 1) is another product that has been thoroughly investigated for its insecticidal properties. In fact, this bis-anthraquinone was first characterized for toxicity against D. melanogaster (ED50 27.6 µg/mL) [93]. Later on, toxic effects were described on Sf9, a cell line derived from ovarian cells of the fall armyworm (Spodoptera frugiperda) (ID50 1.2 µg/mL) [94] and in direct feeding assays on larvae of the spruce budworm (Choristoneura fumiferana) (Lepidoptera, Tortricidae) [95,96]. Moreover, toxic effects resulted when larvae of the latter insect fed on the needles of white spruce (Picea glauca) colonized by a rugulosin-producing endophytic fungus, which contained the toxin at a concentration that was effective at retarding the larval growth in vitro [97,98]. In line with the presumptive role of anthraquinones in defensive mutualism established between plants and endophytic fungi [99], and with reference to the known endophytic occurrence of T. rugulosus and other Talaromyces spp. producing these compounds [100,101,102], this mechanism could affect insect herbivory and protect plants where these fungi are able to develop endophytically. It is interesting to note that in the study by Miller et al. [97], a similar effect was observed after the inoculation of spruce needles with an unidentified endophytic strain producing vermiculine (Figure 1), a macrodiolide antibiotic that so far has only been reported from T. flavus [103].
5. Conclusions
The analysis of the available literature has disclosed that many Talaromyces spp. are able to establish ecological relationships with insects. At least 8 out of 27 species have been first or exclusively recovered from entomological specimens, indicating that insects represent a source of unknown biodiversity with reference to this fungal genus. Indeed, the recovery of Talaromyces strains even from freshwater and troglophile insects represents evidence that more cases of interesting associations could be disclosed as a result of investigations carried out in diverse ecological contexts.
However, in most instances, the reported interactions must be recorded as neutral, in the absence of circumstantial evidence clearly referable to either mutualistic or antagonistic effects. Apart from citations merely concerning occurrence, to be considered as occasional unless corroborated by additional findings in the future, the symbiotic interactions could be inferred as being mutualistic in the case of gut-associated strains that might contribute to the digestion of specific feed. Conversely, in addition to the cases where detrimental effects on insect viability have been experimentally demonstrated, indirect antagonistic behavior can be conjectured in the case of species/strains found in the gardens of leaf-cutting ants and in combs of Macrotermes spp. with reference to mycoparasitism possibly exerted against the cropped fungi.
Antagonistic interactions of fungi are often mediated by the production of bioactive metabolites, which could either affect the insect’s development when direct contact is established or be responsible for toxic or phagodeterrent effects when these products are released in plants by endophytic strains. Indeed, this aspect may have a substantial impact in crop protection, with reference to the possibility to exploit the resident mycoflora and/or artificially-administered strains holding this property. Species such as T. flavus, T. pinophilus and T. purpureogenus are already known to be producers of bioactive compounds, playing a fundamental role in antagonism exerted against plant pathogens. Considering that these species are also increasingly reported as plant growth promoters [104], more accurate investigations concerning their anti-insectan aptitude might disclose additional positive effects on plant health and integrate the profile of multipurpose strains to be employed in crop management.
Author Contributions
Conceptualization, R.N.; resources, R.N. and A.B.; data curation, A.B.; writing—original draft preparation, R.N.; writing—review and editing, R.N. and A.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jaber, L.R.; Ownley, B.H. Can we use entomopathogenic fungi as endophytes for dual biological control of insect pests and plant pathogens? Biol. Control 2018, 116, 36–45. [Google Scholar] [CrossRef]
- McKinnon, A.C.; Saari, S.; Moran-Diez, M.E.; Meyling, N.V.; Raad, M.; Glare, T.R. Beauveria bassiana as an endophyte: A critical review on associated methodology and biocontrol potential. BioControl 2017, 62, 1–17. [Google Scholar] [CrossRef]
- Nicoletti, R.; Becchimanzi, A. Endophytism of Lecanicillium and Akanthomyces. Agriculture 2020, 10, 205. [Google Scholar] [CrossRef]
- Ownley, B.H.; Gwinn, K.D.; Vega, F.E. Endophytic fungal entomopathogens with activity against plant pathogens: Ecology and evolution. BioControl 2010, 55, 113–128. [Google Scholar] [CrossRef]
- Lorito, M.; Woo, S.L. Trichoderma: A multi-purpose tool for integrated pest management. In Principles of Plant-Microbe Interactions: Microbes for Sustainable Agriculture; Lugtenberg, B., Ed.; Springer International Publishing: Cham, Switzerland, 2015; pp. 345–353. ISBN 978-3-319-08575-3. [Google Scholar]
- Poveda, J. Trichoderma as biocontrol agent against pests: New uses for a mycoparasite. Biol. Control 2021, 159, 104634. [Google Scholar] [CrossRef]
- Kakvan, N.; Heydari, A.; Zamanizadeh, H.R.; Rezaee, S.; Naraghi, L. Development of new bioformulations using Trichoderma and Talaromyces fungal antagonists for biological control of sugar beet damping-off disease. Crop. Prot. 2013, 53, 80–84. [Google Scholar] [CrossRef]
- Kazerooni, E.A.; Rethinasamy, V.; Al-Sadi, A.M. Talaromyces pinophilus inhibits Pythium and Rhizoctonia-induced damping-off of cucumber. J. Plant. Pathol. 2019, 101, 377–383. [Google Scholar] [CrossRef]
- Naraghi, L.; Heydari, A.; Rezaee, S.; Razavi, M.; Jahanifar, H. Study on antagonistic effects of Talaromyces flavus on Verticillium albo-atrum, the causal agent of potato wilt disease. Crop. Prot. 2010, 29, 658–662. [Google Scholar] [CrossRef]
- Yilmaz, N.; Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Samson, R.A. Polyphasic taxonomy of the genus Talaromyces. Stud. Mycol. 2014, 78, 175–341. [Google Scholar] [CrossRef] [Green Version]
- Rybalchenko, V.M.; Koval, Ė.Z.; Sheremet, V.P. Toxicity of Penicillium funiculosum Thom against larvae of blood-sucking mosquitoes. Mikol. Fitopatol. 1990, 24, 116–121. [Google Scholar]
- Fouillaud, M.; Morel, G. Fungi associated with nests of the paper wasp Polistes hebraeus (Hymenoptera: Vespidae) on La Reunionisland. Environ. Entomol. 1995, 24, 298–305. [Google Scholar] [CrossRef]
- Da Costa, G.L.; de Oliveira, P.C. Penicillium species in mosquitoes from two Brazilian regions. J. Basic Microbiol. 1998, 38, 343–347. [Google Scholar] [CrossRef]
- Gunde-Cimerman, N.; Zalar, P.; Jeram, S. Mycoflora of cave cricket Troglophilus neglectus cadavers. Mycopathologia 1998, 141, 111–114. [Google Scholar] [CrossRef]
- Jaber, S.; Mercier, A.; Knio, K.; Brun, S.; Kambris, Z. Isolation of fungi from dead Arthropods and identification of a new mosquito natural pathogen. Parasites Vectors 2016, 9, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadj Taieb, K.; Gharsallah, H.; Ksentini, I.; Leclerque, A.; Schuster, C.; Cheffi, M.; Naayma, S.; Triki, M.A.; Ksantini, M. Screening of biological activities of fungi associated with pistachio bark beetle, Chaetoptelius vestitus (Coleoptera, Curculionidae), infesting pistachio cultivations in Tunisia. J. Appl. Microbiol. 2020, 128, 1472–1485. [Google Scholar] [CrossRef]
- Barbosa, R.N.; Bezerra, J.D.P.; Souza-Motta, C.M.; Frisvad, J.C.; Samson, R.A.; Oliveira, N.T.; Houbraken, J. New Penicillium and Talaromyces species from honey, pollen and nests of stingless bees. Antonie Van Leeuwenhoek 2018, 111, 1883–1912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seifert, K.; Hoekstra, E.; Frisvad, J.; Louis-Seize, G. Penicillium cecidicola, a new species on cynipid insect galls on Quercus pacifica in the western United States. Stud. Mycol. 2004, 50, 517–523. [Google Scholar]
- Herlinda, S.; Efendi, R.A.; Suharjo, R.; Hasbi, H.; Setiawan, A.; Elfita, E.; Verawaty, M. New emerging entomopathogenic fungi isolated from soil in South Sumatra (Indonesia) and their filtrate and conidial insecticidal activity againts Spodoptera litura. Biodiversitas 2020, 21, 5102–5113. [Google Scholar]
- Hernández, M.C.; Pildain, M.B.; Novas, M.V.; Sacco, J.; Lopez, S.E. Mycobiota associated with larval mines of Thrypticus truncatus and T. sagittatus (Diptera, Dolichopodidae) on waterhyacinth, Eichhornia crassipes, in Argentina. Biol. Control 2007, 41, 321–326. [Google Scholar] [CrossRef]
- Sun, B.-D.; Liu, X.-Z. Occurrence and diversity of insect-associated fungi in natural soils in China. Appl. Soil Ecol. 2008, 39, 100–108. [Google Scholar] [CrossRef]
- Sun, B.-D.; Yu, H.; Chen, A.J.; Liu, X.-Z. Insect-associated fungi in soils of field crops and orchards. Crop. Prot. 2008, 27, 1421–1426. [Google Scholar] [CrossRef]
- Oliveira, I.; Pereira, J.A.; Lino-Neto, T.; Bento, A.; Baptista, P. Fungal diversity associated to the olive moth, Prays oleae Bernard: A survey for potential entomopathogenic fungi. Microb. Ecol. 2012, 63, 964–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirker, G.T.; Wagner, T.; Diehl, S. Relationship between wood-inhabiting fungi and Reticulitermes spp. Int. Biodeter. Biodegrad. 2012, 72, 18–25. [Google Scholar] [CrossRef]
- Ncube, E.; Truter, M.; Flett, B.C.; Van den Berg, J.; Erasmus, A.; Viljoen, A. Fungal mycoflora associated with Busseola fusca Frass in maize plants. Afr. Entomol. 2020, 28, 394–405. [Google Scholar] [CrossRef]
- Abdella, M.M.; Ali, I.; El Henidi, A.H.; Mostafa, S.M. Bioactivity of Penicillium spores suspension and plant extracts against Liriomyza sativae (Blachard) and their associated parasitoids. Fayoum J. Agric. Res. Devel. 2006, 20, 1–10. [Google Scholar] [CrossRef]
- Marti, G.A.; García, J.J.; Cazau, M.C.; López Lastra, C.C. Fungal flora of the digestive tract of Triatoma infestans (Hemiptera: Reduviidae) from Argentina. Bol. Soc. Argentina Bot. 2007, 42, 175–179. [Google Scholar]
- Davydenko, K.; Vasaitis, R.; Menkis, A. Fungi associated with Ips acuminatus (Coleoptera: Curculionidae) in Ukraine with a special emphasis on pathogenicity of ophiostomatoid species. Eur. J. Entomol. 2017, 114, 77–85. [Google Scholar] [CrossRef]
- Chakraborty, A.; Modlinger, R.; Ashraf, M.Z.; Synek, J.; Schlyter, F.; Roy, A. Core mycobiome and their ecological relevance in the gut of five Ips bark beetles (Coleoptera: Curculionidae: Scolytinae). Front. Microbiol. 2020, 11, 2134. [Google Scholar] [CrossRef]
- Abdel Galil, F.A.; Moharram, A.M.; Mahmoud, M.A.; Hafez, W.M.M. Biocontrol of bean and wheat aphids by fungi isolated from indigenous and invasive insects collected from different locations in Minia governorate, Egypt. Egypt. Acad. J. Biol. Sci. 2019, 11, 79–90. [Google Scholar] [CrossRef] [Green Version]
- Gharsallah, H.; Ksentini, I.; Naayma, S.; Hadj Taieb, K.; Abdelhedi, N.; Schuster, C.; Triki, M.A.; Ksantini, M.; Leclerque, A. Identification of fungi in Tunisian olive orchards: Characterization and biological control potential. BMC Microbiol. 2020, 20, 1–13. [Google Scholar] [CrossRef]
- Crous, P.W.; Cowan, D.A.; Maggs-Kölling, G.; Yilmaz, N.; Larsson, E.; Angelini, C.; Brandrud, T.E.; Dearnaley, J.D.W.; Dima, B.; Dovana, F.; et al. Fungal planet description sheets: 1112–1181. Persoonia 2020, 45, 251. [Google Scholar] [CrossRef]
- Rodrigues, A.; Mueller, U.G.; Ishak, H.D.; Bacci, M.; Pagnocca, F.C. Ecology of microfungal communities in gardens of fungus-growing ants (Hymenoptera: Formicidae): A year-long survey of three species of attine ants in Central Texas. FEMS Microbiol. Ecol. 2011, 78, 244–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dohet, L.; Gregoire, J.-C.; Berasategui, A.; Kaltenpoth, M.; Biedermann, P.H. Bacterial and fungal symbionts of parasitic Dendroctonus bark beetles. FEMS Microbiol. Ecol. 2016, 92, fiw129. [Google Scholar] [CrossRef] [Green Version]
- Ibarra-Juarez, L.A.; Desgarennes, D.; Vázquez-Rosas-Landa, M.; Villafan, E.; Alonso-Sánchez, A.; Ferrera-Rodríguez, O.; Moya, A.; Carrillo, D.; Cruz, L.; Carrión, G. Impact of rearing conditions on the ambrosia beetle’s microbiome. Life 2018, 8, 63. [Google Scholar] [CrossRef] [Green Version]
- Belmont-Montefusco, E.L.; Nacif-Marçal, L.; de Assunção, E.N.; Hamada, N.; Nunes-Silva, C.G. Cultivable cellulolytic fungi isolated from the gut of Amazonian aquatic insects. Acta Amaz. 2020, 50, 346–354. [Google Scholar] [CrossRef]
- Marfetan, J.A.; Folgarait, P.J. Diversity of mycobiota in colonies of different species of leaf-cutting ants and sampling sites across Argentina. Ecol. Austral. 2017, 27, 449–461. [Google Scholar]
- Nuotclà, J.A.; Diehl, J.M.C.; Taborsky, M. Habitat quality determines dispersal decisions and fitness in a beetle–fungus mutualism. Front. Ecol. Evol. 2021, 9, 242. [Google Scholar] [CrossRef]
- Guedegbe, H.J.; Miambi, E.; Pando, A.; Roman, J.; Houngnandan, P.; Rouland-Lefevre, C. Occurrence of fungi in combs of fungus-growing termites (Isoptera: Termitidae, Macrotermitinae). Mycol. Res. 2009, 113, 1039–1045. [Google Scholar] [CrossRef] [PubMed]
- Chouvenc, T.; Bardunias, P.; Efstathion, C.A.; Chakrabarti, S.; Elliott, M.L.; Giblin-Davis, R.; Su, N.-Y. Resource opportunities from the nest of dying subterranean termite (Isoptera: Rhinotermitidae) colonies: A laboratory case of ecological succession. Ann. Entomol. Soc. Am. 2013, 106, 771–778. [Google Scholar] [CrossRef]
- Angleró-Rodríguez, Y.I.; Talyuli, O.A.; Blumberg, B.J.; Kang, S.; Demby, C.; Shields, A.; Carlson, J.; Jupatanakul, N.; Dimopoulos, G. An Aedes aegypti-associated fungus increases susceptibility to dengue virus by modulating gut trypsin activity. Elife 2017, 6, e28844. [Google Scholar] [CrossRef] [PubMed]
- Kalansuriya, P.; Quezada, M.; Espósito, B.P.; Capon, R.J. Talarazines A–E: Noncytotoxic iron (III) chelators from an Australian mud dauber wasp-associated fungus, Talaromyces sp. (CMB-W045). J. Nat. Prod. 2017, 80, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Ziganshina, E.E.; Mohammed, W.S.; Shagimardanova, E.I.; Vankov, P.Y.; Gogoleva, N.E.; Ziganshin, A.M. Fungal, bacterial, and archaeal diversity in the digestive tract of several beetle larvae (Coleoptera). BioMed Res. Int. 2018, 2018, 6765438. [Google Scholar] [CrossRef]
- Wang, H.-M.; Liu, F.; Zhang, S.-F.; Kong, X.-B.; Lu, Q.; Zhang, Z. Epibiotic fungal communities of three Tomicus spp. infesting pines in southwestern China. Microorganisms 2020, 8, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.; Song, Y.; Shentu, X.; Yu, X. Antimicrobials affect the fat body microbiome and increase the brown planthopper mortality. Front. Physiol. 2021, 12, 347. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Shi, J.; Xiong, Z.; Shentu, X.; Yu, X. Three antimicrobials alter gut microbial communities and causing different mortality of brown planthopper, Nilaparvata lugens Stål. Pesticide Biochem. Physiol. 2021, 174, 104806. [Google Scholar] [CrossRef]
- Nagam, V.; Aluru, R.; Shoaib, M.; Dong, G.-R.; Li, Z.; Pallaval, V.B.; Ni, J.-F. Diversity of fungal isolates from fungus-growing termite Macrotermes barneyi and characterization of bioactive compound from Xylaria escharoidea. Insect Sci. 2021, 28, 392–402. [Google Scholar] [CrossRef]
- Giordano, L.; Garbelotto, M.; Nicolotti, G.; Gonthier, P. Characterization of fungal communities associated with the bark beetle Ips typographus varies depending on detection method, location, and beetle population levels. Mycol. Progr. 2013, 12, 127–140. [Google Scholar] [CrossRef]
- Panyasiri, C.; Attathom, T.; Poehling, H.-M. Pathogenicity of entomopathogenic fungi-potential candidates to control insect pests on tomato under protected cultivation in Thailand. J. Plant. Dis. Prot. 2007, 114, 278–287. [Google Scholar] [CrossRef]
- Reis, B.M.; dos, S.; Silva, A.; Alvarez, M.R.; de Oliveira, T.B.; Rodrigues, A. Fungal communities in gardens of the leafcutter ant Atta cephalotes in forest and cabruca agrosystems of southern Bahia state (Brazil). Fungal Biol. 2015, 119, 1170–1178. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Ruan, Y.-Y.; Stanley, E.L.; Skelton, J.; Hulcr, J. Plasticity of mycangia in Xylosandrus ambrosia beetles. Insect Sci. 2019, 26, 732–742. [Google Scholar] [CrossRef] [Green Version]
- Sandeepani, H.P.; Ratnaweera, P.B. Antibacterial activity of entomopathogenic fungi isolated from Vespa affinis and Apis dorsata in Sri Lanka. In Proceedings of the 1st International Conference on Frontiers in Chemical Technology, Colombo, Sri Lanka, 20–22 July 2020; p. 29. [Google Scholar]
- Fonseca, Q.R.; Sarquis, M.I.; de, M.; Hamada, N.; Alencar, Y.B. Occurrence of filamentous fungi in Simulium goeldii Cerqueira & Nunes de Mello (Diptera: Simuliidae) larvae in central Amazonia, Brazil. Braz. J. Microbiol. 2008, 39, 282–285. [Google Scholar]
- Jayaprakash, A.; Ebenezer, P. A new report on mycobiota associated with Ropalidia marginata paper nests. Indian J. Sci. Technol. 2010, 3, 6–8. [Google Scholar] [CrossRef]
- Galíndez-Chicaíza, E.; Lagos-Mora, L.E.; Castillo-Belalcázar, G.; Salazar-González, C.; Betancourth-García, C. Hongos detectados en la entomofauna asociada a Espeletia pycnophylla. Rev. UDCA Actualidad Divulgación Científica 2020, 23, 2. [Google Scholar]
- Houbraken, J.; Kocsubé, S.; Visagie, C.M.; Yilmaz, N.; Wang, X.-C.; Meijer, M.; Kraak, B.; Hubka, V.; Bensch, K.; Samson, R.A. Classification of Aspergillus, Penicillium, Talaromyces and related genera (Eurotiales): An overview of families, genera, subgenera, sections, series and species. Stud. Mycol. 2020, 95, 5–169. [Google Scholar] [CrossRef] [PubMed]
- Brentassi, M.E.; Medina, R.; de la Fuente, D.; Franco, M.E.; Toledo, A.V.; Saparrat, M.C.; Balatti, P.A. Endomycobiome associated with females of the planthopper Delphacodes kuscheli (Hemiptera: Delphacidae): A metabarcoding approach. Heliyon 2020, 6, e04634. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Martínez, M.G.; Cirerol-Cruz, B.E.; Torres-Estrada, J.L.; López, M.H.R. Potential for entomopathogenic fungi to control Triatoma dimidiata (Hemiptera: Reduviidae), a vector of chagas disease in Mexico. Rev. Soc. Bras. Med. Trop. 2014, 47, 716–722. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.-H.; Xu, N.; Chen, H.; Liu, Q.; Pu, Q.-Y.; Qin, D.-Y.; Zhai, Y.-F.; Yu, Y. Impact of selected fungi from an artificial diet on the growth and development of Drosophila suzukii (Diptera: Drosophilidae). J. Asia-Pac. Entomol. 2017, 20, 141–149. [Google Scholar] [CrossRef]
- Fitriana, Y.; Suharjo, R.; Swibawa, I.G.; Purnomo; Lestari, P.; Merdiana, E. Influence of culture medium on the sporulation and viability of Aspergillus spp. and Talaromyces spp. entomopathogenic fungi. J. Hama Dan Penyakit Tumbuh. Trop. 2018, 18, 12–22. [Google Scholar] [CrossRef] [Green Version]
- Pangesti, I.R.; Suharjo, R.; Fitriana, Y.; Merdiana, E.; Pasaribu, L.T.; Swibawa, I.G.; Hidayat, K.F. Study of Talaromyces spp. with multiple beneficial task. In Proceedings of the 5th Conference Inter-Academia Asia 2018, Shizuoka, Japan, 3–5 December 2018; p. 25. [Google Scholar]
- Khaerati, K.; Indriati, G.; Wardiana, E. The effectiveness of bioinsecticide based on entomopathogenic fungi of Talaromyces pinophilus and vegetable oil on coffee berry borer. J. Tanam. Ind. Dan Penyegar 2020, 7, 93–108. [Google Scholar] [CrossRef]
- Mukherjee, A.; Debnath, P.; Ghosh, S.K.; Medda, P.K. Biological control of papaya aphid (Aphis gossypii Glover) using entomopathogenic fungi. Vegetos 2020, 33, 1–10. [Google Scholar] [CrossRef]
- Berestetskiy, A.; Hu, Q. The chemical ecology approach to reveal fungal metabolites for Arthropod pest management. Microorganisms 2021, 9, 1379. [Google Scholar] [CrossRef] [PubMed]
- Frisvad, J.C. Taxonomy, chemodiversity, and chemoconsistency of Aspergillus, Penicillium, and Talaromyces species. Front. Microbiol. 2015, 5, 773. [Google Scholar] [CrossRef] [Green Version]
- Nicoletti, R.; Manzo, E.; Ciavatta, M.L. Occurence and bioactivities of funicone-related compounds. Int. J. Mol. Sci. 2009, 10, 1430–1444. [Google Scholar] [CrossRef]
- Nicoletti, R.; Salvatore, M.M.; Andolfi, A. Secondary metabolites of mangrove-associated strains of Talaromyces. Mar. Drugs 2018, 16, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalansuriya, P.; Khalil, Z.G.; Salim, A.A.; Capon, R.J. Talarophenol sulfate and talarophilones from the Australian mud dauber wasp-associated fungus, Talaromyces sp. CMB-W045. Tetrahedron Lett. 2019, 60, 151157. [Google Scholar] [CrossRef]
- Santamarina, M.P.; Jimenez, M.; Sanchis, V.; Garcia, F.; Hernandez, E. A strain of Penicillium funiculosum Thorn with activity against Panonychus ulmi Koch (Acar., Tetranychidae). J. Appl. Entomol. 1987, 103, 471–476. [Google Scholar] [CrossRef]
- Castillo, M.A.; Moya, P.; Cantín, A.; Miranda, M.A.; Primo, J.; Hernández, E.; Primo-Yúfera, E. Insecticidal, anti-juvenile hormone, and fungicidal activities of organic extracts from different Penicillium species and their isolated active components. J. Agric. Food Chem. 1999, 47, 2120–2124. [Google Scholar] [CrossRef]
- Chen, L.; Liu, T.; Duan, Y.; Lu, X.; Yang, Q. Microbial secondary metabolite, phlegmacin B1, as a novel inhibitor of insect chitinolytic enzymes. J. Agric. Food Chem. 2017, 65, 3851–3857. [Google Scholar] [CrossRef]
- Nicoletti, R.; Scognamiglio, M.; Fiorentino, A. Structural and bioactive properties of 3-O-methylfunicone. Mini Rev. Med. Chem. 2014, 14, 1043–1047. [Google Scholar] [CrossRef]
- De Stefano, S.; Nicoletti, R.; Milone, A.; Zambardino, S. 3-o-Methylfunicone, a fungitoxic metabolite produced by the fungus Penicillium pinophilum. Phytochemistry 1999, 52, 1399–1401. [Google Scholar] [CrossRef]
- Vinale, F.; Nicoletti, R.; Lacatena, F.; Marra, R.; Sacco, A.; Lombardi, N.; d’Errico, G.; Digilio, M.C.; Lorito, M.; Woo, S.L. Secondary metabolites from the endophytic fungus Talaromyces pinophilus. Nat. Prod. Res. 2017, 31, 1778–1785. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Li, C.; Zhu, J.; Zhang, L.; Wang, Y.; Chen, Q.; Xu, L.; Zhang, S.; Fang, Y.; Liu, T. A new meroterpenoid from endophytic fungus Talaromyces amestolkiae CS-O-1. Biochem. Syst. Ecol. 2020, 93, 104186. [Google Scholar] [CrossRef]
- Guo, J.; Ran, H.; Zeng, J.; Liu, D.; Xin, Z. Tafuketide, a phylogeny-guided discovery of a new polyketide from Talaromyces funiculosus Salicorn 58. Appl. Microbiol. Biotechnol. 2016, 100, 5323–5338. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, H.; Oka, Y.; Kai, K.; Akiyama, K. A new meroterpenoid, chrodrimanin C, from YO-2 of Talaromyces sp. Biosci. Biotechnol. Biochem. 2012, 76, 745–748. [Google Scholar] [CrossRef] [Green Version]
- Kong, F.-D.; Ma, Q.-Y.; Huang, S.-Z.; Wang, P.; Wang, J.-F.; Zhou, L.-M.; Yuan, J.-Z.; Dai, H.-F.; Zhao, Y.-X. Chrodrimanins K–N and related meroterpenoids from the fungus Penicillium sp. SCS-KFD09 isolated from a marine worm, Sipunculus nudus. J. Nat. Prod. 2017, 80, 1039–1047. [Google Scholar] [CrossRef]
- Zhou, H.; Li, L.; Wang, W.; Che, Q.; Li, D.; Gu, Q.; Zhu, T. Chrodrimanins I and J from the Antarctic moss-derived fungus Penicillium funiculosum GWT2-24. J. Nat. Prod. 2015, 78, 1442–1445. [Google Scholar] [CrossRef]
- Hayashi, H.; Oka, Y.; Kai, K.; Akiyama, K. New chrodrimanin congeners, chrodrimanins D–H, from YO-2 of Talaromyces sp. Biosci. Biotechnol. Biochem. 2012, 76, 1765–1768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Furutani, S.; Ihara, M.; Ling, Y.; Yang, X.; Kai, K.; Hayashi, H.; Matsuda, K. Meroterpenoid chrodrimanins are selective and potent blockers of insect GABA-gated chloride channels. PLoS ONE 2015, 10, e0122629. [Google Scholar] [CrossRef] [PubMed]
- Chaiyosang, B.; Kanokmedhakul, K.; Soytong, K.; Poeaim, S.; Soytong, M.; Hadsadee, S.; Jungsuttiwong, S.; Yahuafai, J.; Siripong, P.; Kanokmedhakul, S. New pyrrolobenzoxazine sesquiterpenoid derivatives from the fungus Talaromyces trachyspermus. Planta Med. 2021, 87, 600–610. [Google Scholar] [CrossRef]
- Horikoshi, R.; Goto, K.; Mitomi, M.; Oyama, K.; Sunazuka, T.; Ōmura, S. Identification of pyripyropene A as a promising insecticidal compound in a microbial metabolite screening. J. Antibiot. 2017, 70, 272–276. [Google Scholar] [CrossRef]
- Horikoshi, R.; Goto, K.; Mitomi, M.; Oyama, K.; Sunazuka, T.; Ōmura, S. Insecticidal properties of pyripyropene A, a microbial secondary metabolite, against agricultural pests. J. Pesticide Sci. 2018, 43, 266–271. [Google Scholar] [CrossRef]
- Fuse, S.; Matsumura, K.; Johmoto, K.; Uekusa, H.; Tanaka, H.; Hirose, T.; Sunazuka, T.; Ōmura, S.; Takahashi, T. The design, synthesis, and evaluation of 1,5,7-trisubstituted-3-pyridyl-xanthones for use as insecticides starting from pyripyropene A. Chemistry 2016, 22, 18450–18455. [Google Scholar] [CrossRef]
- Goto, K.; Horikoshi, R.; Mitomi, M.; Oyama, K.; Hirose, T.; Sunazuka, T.; Ōmura, S. Synthesis and insecticidal efficacy of pyripyropene derivatives focusing on the C-1, C-7, and C-11 positions’ substituent groups. J. Antibiot. 2018, 71, 785–797. [Google Scholar] [CrossRef]
- Goto, K.; Horikoshi, R.; Mitomi, M.; Oyama, K.; Hirose, T.; Sunazuka, T.; Ōmura, S. Synthesis and insecticidal efficacy of pyripyropene derivatives. Part II—Invention of afidopyropen. J. Antibiot. 2019, 72, 661–681. [Google Scholar] [CrossRef]
- Tomoda, H.; Kim, Y.K.; Nishida, H.; Masuma, R.; Omura, S. Pyripyropenes, novel inhibitors of acyl-coa: Cholesterol acyltransferase produced by Aspergillus fumigatus I. Production, isolation, and biological properties. J. Antibiot. 1994, 47, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Thapa, S.; Lv, M.; Xu, H. Acetylcholinesterase: A primary target for drugs and insecticides. Mini Rev. Med. Chem. 2017, 17, 1665–1676. [Google Scholar] [CrossRef]
- Lima, M.T.N.S.; dos Santos, L.B.; Bastos, R.W.; Nicoli, J.R.; Takahashi, J.A. Antimicrobial activity and acetylcholinesterase inhibition by extracts from chromatin modulated fungi. Braz. J. Microbiol. 2018, 49, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Ohlendorf, B.; Oesker, V.; Wiese, J.; Malien, S.; Schmaljohann, R.; Imhoff, J.F. Acetylcholinesterase inhibitors from a marine fungus Talaromyces sp. strain LF458. Mar. Biotechnol. 2015, 17, 110–119. [Google Scholar] [CrossRef]
- Breen, J.; Dacre, J.C.; Raistrick, H.; Smith, G. Studies in the biochemistry of micro-organisms. 95. Rugulosin, a crystalline colouring matter of Penicillium rugulosum Thom. Biochem. J. 1955, 60, 618–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobias, J.; Betina, V.; Nemec, P. Insecticidal activity of ramihyfin-A, citrinin and rugulosin. Biologia 1980, 35, 431–434. [Google Scholar]
- Watts, P.; Kittakoop, P.; Veeranondha, S.; Wanasith, S.; Thongwichian, R.; Saisaha, P.; Intamas, S.; Hywel-Jones, N.L. Cytotoxicity against insect cells of entomopathogenic fungi of the genera Hypocrella (anamorph Aschersonia): Possible agents for biological control. Mycol. Res. 2003, 107, 581–586. [Google Scholar] [CrossRef]
- Calhoun, L.A.; Findlay, J.A.; David Miller, J.; Whitney, N.J. Metabolites toxic to spruce budworm from balsam fir needle endophytes. Mycol. Res. 1992, 96, 281–286. [Google Scholar] [CrossRef]
- Sumarah, M.W.; Adams, G.W.; Berghout, J.; Slack, G.J.; Wilson, A.M.; Miller, J.D. Spread and persistence of a rugulosin-producing endophyte in Picea glauca seedlings. Mycol. Res. 2008, 112, 731–736. [Google Scholar] [CrossRef]
- Miller, J.D.; Mackenzie, S.; Foto, M.; Adams, G.W.; Findlay, J.A. Needles of white spruce inoculated with rugulosin-producing endophytes contain rugulosin reducing spruce budworm growth rate. Mycol. Res. 2002, 106, 471–479. [Google Scholar] [CrossRef]
- Sumarah, M.W.; Miller, J.D.; Adams, G.W. Measurement of a rugulosin-producing endophyte in white spruce seedlings. Mycologia 2005, 97, 770–776. [Google Scholar] [CrossRef] [PubMed]
- Gessler, N.N.; Egorova, A.S.; Belozerskaya, T.A. Fungal anthraquinones. Appl. Biochem. Microbiol. 2013, 49, 85–99. [Google Scholar] [CrossRef]
- Naseem, F.; Kayang, H.; Naseem, F.; Kayang, H. Endophytic fungal diversity of endemic carnivorous plant Nepenthes khasiana in Meghalaya, India. Stud. Fungi 2021, 6, 138–150. [Google Scholar] [CrossRef]
- Urooj, F.; Farhat, H.; Ali, S.A.; Ahmed, M.; Sultana, V.; Shams, Z.I.; Ara, J.; Ehteshamul-Haque, S. Role of endophytic Penicillium species in suppressing the root rotting fungi of sunflower. Pak. J. Bot. 2018, 50, 1621–1628. [Google Scholar]
- Yilmaz, N.; Visagie, C.M.; Frisvad, J.C.; Houbraken, J.; Jacobs, K.; Samson, R.A. Taxonomic re-evaluation of species in Talaromyces section Islandici, using a polyphasic approach. Persoonia 2016, 36, 37–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proksa, B. Talaromyces flavus and its metabolites. Chem. Papers 2010, 64, 696–714. [Google Scholar] [CrossRef]
- Patel, D.; Patel, A.; Patel, M.; Goswami, D. Talaromyces pinophilus strain M13: A portrayal of novel groundbreaking fungal strain for phytointensification. Environ. Sci. Pollut. Res. 2021, 28, 8758–8769. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Molecular structure of anti-insectan products of Talaromyces spp.


{kind=link}
Table 1.
Occurrence of Talaromyces species found in association with insects.
| Species | Insect Source * | Location | Year | Reference |
|---|---|---|---|---|
| T. amestolkiae | corpses of Capnodis tenebrionis, Culex sp., unidentified Pyralidae, unidentified Pyrrhocoridae | Lebanon | 2016 | [15] |
| T. atroroseus | living adults of Chaetoptelius vestitus | Tunisia | 2020 | [16] |
| T. brasiliensis | nests of Melipona scutellaris | Brazil | 2018 | [17] |
| T. cecidicola | galls of unidentified cynipid on Quercus pacifica | Western USA | 2004 | [18] |
| T. dendriticus | galls of unidentified insect on Eucalyptus leaves | Australia | 2004 | [18] |
| T. diversus | meconia of Polistes hebraeus | La Reunion | 1995 | [12] |
| larva of Tenebrio molitor (bait) | Sumatra | 2020 | [19] | |
| T. duclauxii | dead larvae and adults of Troglophilus neglectus | Slovenia | 1998 | [14] |
| T. erythromellis | galls of Diplolepis rosae on Rosa sitchensis | Canada | 2004 | [18] |
| T. flavus | mines of Thrypticus truncatus, Thrypticus sagittatus on Eichhornia crassipes | Argentina | 2007 | [20] |
| larvae of Galleria mellonella (bait) | China | 2008 | [21,22] | |
| larvae of Prays oleae | Portugal | 2012 | [23] | |
| coarse wood debris with termites (Reticulitermes sp.) | Mississippi | 2012 | [24] | |
| frass of Busseola fusca | South Africa | 2020 | [25] | |
| T. funiculosus | larvae of Aedes cantans and Aedes communis | Ukraine | 1990 | [11] |
| dead larvae and pupae of Liriomyza sativae | Egypt | 2006 | [26] | |
| T. aff. helicus | gut of Triatoma infestans | Argentina | 2007 | [27] |
| T. minioluteus | adults of Ips acuminatus | Ukraine | 2017 | [28] |
| gut of Ips sexdentatus | Czechia | 2020 | [29] | |
| T. mycothecae | nests of M. scutellaris | Brazil | 2018 | [17] |
| T. pigmentosus | nests of M. scutellaris | Brazil | 2018 | [17] |
| T. pinophilus | larvae of P. oleae | Portugal | 2012 | [23] |
| Rhynchophorus ferrugineus | Egypt | 2019 | [30] | |
| larvae of Bactrocera oleae, Euphyllura olivina | Tunisia | 2020 | [31] | |
| dead adults of C. vestitus | Tunisia | 2020 | [16] | |
| T. pseudostromaticus | galls of D. rosae on R. sitchensis | Canada | 2004 | [18] |
| T. pulveris | bore dust of Xestobium rufovillosum | France | 2020 | [32] |
| T. purpureogenus | mosquitoes (Aedes sp., Anopheles sp., Mansonia sp.) | Brazil | 1998 | [13] |
| gut of T. infestans | Argentina | 2007 | [27] | |
| nest of Trachymyrmex septentrionalis | Texas | 2011 | [33] | |
| larvae of Dendroctonus punctatus | France | 2016 | [34] | |
| adults of I. acuminatus | Ukraine | 2017 | [28] | |
| adults of Xyleborus affinis, Xyleborus bispinatus | Mexico | 2018 | [35] | |
| gut of larva of Triplectides sp. | Brazil | 2020 | [36] | |
| T. radicus | larvae of D. punctatus | France | 2016 | [34] |
| T. ruber | gut of T. infestans | Argentina | 2007 | [27] |
| T. rugulosus | nests of Acromyrmex spp. | Argentina | 2017 | [37] |
| gut of Ips duplicatus, Ips typographus, I. acuminatus, I. sexdentatus, Polygraphus poligraphus | Czechia | 2020 | [29] | |
| gardens of Xyleborinus saxesenii in laboratory rearing | Switzerland | 2021 | [38] | |
| T. scorteus | nests of M. scutellaris | Brazil | 2018 | [17] |
| Talaromyces sp. | fungal comb in nest of Macrotermes carbonarius | Vietnam | 2019 | [39] |
| nest of Coptotermes formosanus | Florida | 2013 | [40] | |
| gut of Aedes aegypti | Puerto Rico | 2017 | [41] | |
| unidentified mud dauber wasp | Australia | 2017 | [42] | |
| gut of larvae of Rhagium inquisitor | Russia | 2018 | [43] | |
| adults of Tomicus brevipilosus, Tomicus minor, Tomicus yunnanensis | China | 2020 | [44] | |
| adult of Platypus cylindrus | Italy | 2020 | This paper | |
| abdomen fat body of Nilaparvata lugens | China | 2021 | [45] | |
| gut of N. lugens | China | 2021 | [46] | |
| T. stollii | fungal comb in nest of Macrotermes barneyi | China | 2021 | [47] |
| T. trachyspermus | meconia of P. hebraeus | La Reunion | 1995 | [12] |
| larvae of G. mellonella (bait) | China | 2008 | [21] | |
| adults of I. typographus, | Italy | 2013 | [48] | |
| T. verruculosus | unidentified thrips | Thailand | 2007 | [49] |
| nests of Atta texana, Cyphomyrmex wheeleri, T. septentrionalis | Texas | 2011 | [33] | |
| nests of Atta cephalotes | Brazil | 2015 | [50] | |
| mycangia of Xylosandrus crassiusculus | China, Florida | 2019 | [51] | |
| larvae of B. oleae, E. olivina | Tunisia | 2020 | [31] | |
| T. versatilis | dead adult of Apis dorsata | Sri Lanka | 2020 | [52] |
| T. wortmannii(T. variabilis) | larvae of Simulium goeldii | Brazil | 2008 | [53] |
| nest of Ropalidia marginata | India | 2010 | [54] | |
| larvae and adults of Dendroctonus micans, Dendroctonus valens | France | 2016 | [34] | |
| nests and honey of M. scutellaris | Brazil | 2018 | [17] | |
| gut of Ips cembrae, I. acuminatus, I. duplicatus, I. sexdentatus, I. typographus | Czechia | 2020 | [29] | |
| adults of Diabrotica sp. and Neomyopites sp. | Colombia | 2020 | [55] |
* Colors are indicative of the order to which the species belong, as follows: Blattodea; Coleoptera; Diptera; Hemiptera; Hymenoptera; Lepidoptera; Orthoptera; Thysanoptera; Trichoptera.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Nicoletti, R.; Becchimanzi, A. Talaromyces–Insect Relationships. Microorganisms 2022, 10, 45. https://doi.org/10.3390/microorganisms10010045
AMA Style
Nicoletti R, Becchimanzi A. Talaromyces–Insect Relationships. Microorganisms. 2022; 10(1):45. https://doi.org/10.3390/microorganisms10010045
Chicago/Turabian StyleNicoletti, Rosario, and Andrea Becchimanzi. 2022. "Talaromyces–Insect Relationships" Microorganisms 10, no. 1: 45. https://doi.org/10.3390/microorganisms10010045
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.