
Bacterial Complexity of Traditional Mountain Butter Is Affected by the Malga-Farm of Production

 , and
, and
Abstract
:
1. Introduction
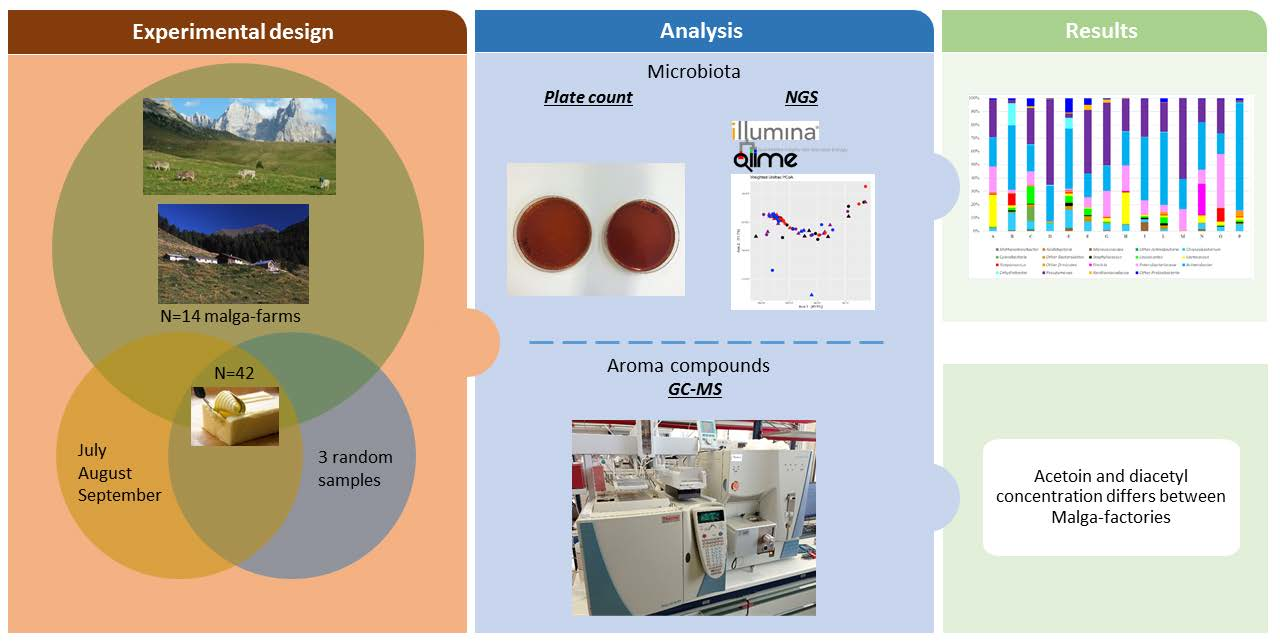
2. Materials and Methods
2.1. Butter Production and Sampling
2.2. Microbiological and Physicochemical Analyses
2.3. DNA Extraction, Miseq Library Preparation, and Illumina Sequencing
2.4. Illumina Data Analysis and Sequences Identification by QIIME2
2.5. GC–MS Analysis of Acetoin and Diacetyl
2.6. Statistical Analysis
3. Results and Discussion
3.1. Microbiological Physicochemical and Aromatic Characteristics of TMB
3.2. Characteristics of Sample Sequence and Alpha Diversity
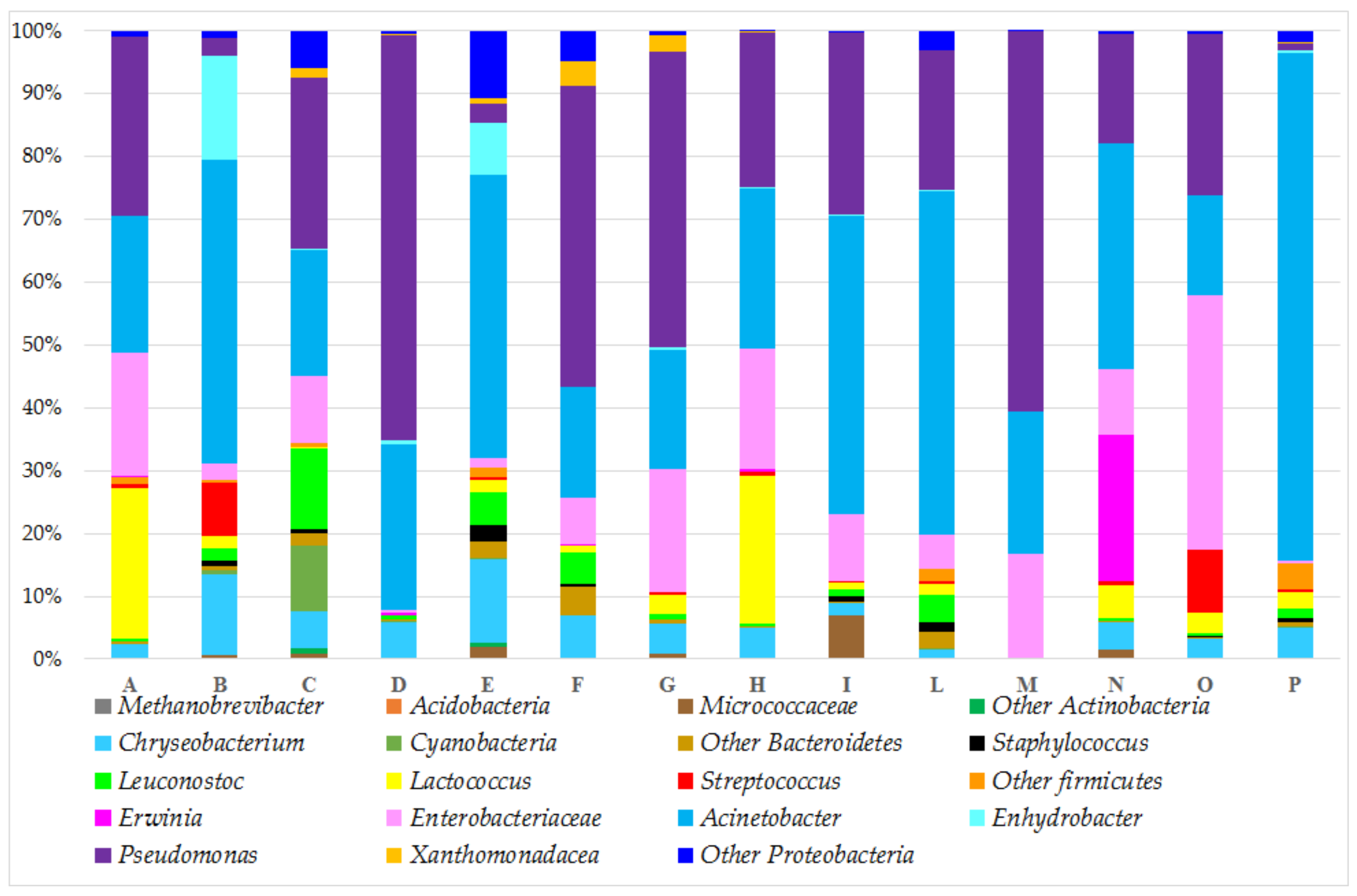
3.3. Taxonomic Composition of the Bacterial Community
3.4. Geographical Discrimination: Val di Sole vs. Valsugana
3.5. Predictive Model of Diacetyl and Acetoin Concentration
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carafa, I.; Nardin, T.; Larcher, R.; Viola, R.; Tuohy, K.; Franciosi, E. Identification and characterization of wild lactobacilli and pediococci from spontaneously fermented Mountain cheese. Food Microbiol. 2015, 48, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Montel, M.C.; Buchin, S.; Mallet, A.; Delbes–Paus, C.; Vuitton, D.A.; Desmasures, N.; Berthier, F. Traditional cheeses: Rich and diverse microbiota with associated benefits. Int. J. Food Microbiol. 2014, 177, 136–154. [Google Scholar] [CrossRef]
- Rousseau, D. Fat crystals and emulsion stability—A review. Food Res. Int. 2000, 33, 3–14. [Google Scholar] [CrossRef]
- Disciplinare di Produzione del Botìro di Primiero di Malga. Available online: https://www.tastetrentino.it/fileadmin/user_upload/download/Disciplinare_Botiro_di_Primiero_di_Malga.pdf (accessed on 17 December 2021).
- Andreo, I.; Doval, M.M.; Romero, A.M.; Judis, M.A. Influence of heating time and oxygen availability on lipid oxidation in meat emulsions. Eur. J. Lipid Sci. Technol. 2003, 105, 207–213. [Google Scholar] [CrossRef]
- Osinchak, J.E.; Hultin, H.O.; Zajicek, O.T.; Kelleher, S.D.; Huang, C.-H. Effect of NaCl on catalysis of lipid oxidation by the soluble fraction of fish muscle. Free Radic. Biol. Med. 1992, 12, 35–41. [Google Scholar] [CrossRef]
- Frankel, E.N. Lipid Oxidation, 2nd ed.; Woodhead Publishing in Food Science, Technology and Nutrition: Cambridge, UK, 2012. [Google Scholar]
- Krause, A.J.; Lopetcharat, K.; Drake, M.A. Identification of the Characteristics That Drive Consumer Liking of Butter. J. Dairy Sci. 2007, 90, 2091–2102. [Google Scholar] [CrossRef]
- Verkerk, G. Pasture-based dairying: Challenges and rewards for New Zealand producers. Theriogenology 2003, 59, 553–561. [Google Scholar] [CrossRef]
- Silva, C.C.G.; Silva, S.P.M.; Prates, J.A.M.; Bessa, R.J.B.; Rosa, H.J.D.; Rego, O.A. Physicochemical traits and sensory quality of commercial butter produced in the Azores. Int. Dairy J. 2019, 88, 10–17. [Google Scholar] [CrossRef]
- O’Callaghan, T.F.; Faulkner, H.; McAuliffe, S.; O’Sullivan, M.G.; Hennessy, D.; Dillon, P.; Kilcawley, K.N.; Stanton, C.; Ross, R.P. Quality characteristics, chemical composition, and sensory properties of butter from cows on pasture versus indoor feeding systems. J. Dairy Sci. 2016, 99, 9441–9460. [Google Scholar] [CrossRef] [Green Version]
- Phumudzo, T.; Ronald, N.; Khayalethu, N.; Fhatuwani, M. Bacterial species identification getting easier. Afr. J. Biotechnol. 2013, 12, 5975–5982. [Google Scholar] [CrossRef]
- Shokralla, S.; Spall, J.L.; Gibson, J.F.; Hajibabaei, M. Next-generation sequencing technologies for environmental DNA research. Mol. Ecol. 2012, 21, 1794–1805. [Google Scholar] [CrossRef] [PubMed]
- El-Hajjaji, S.; Gerard, A.; De Laubier, J.; Lainé, A.; Patz, V.; Sindic, M. Study of the bacterial profile of raw milk butter, made during a challenge test with Listeria monocytogenes, depending on cream maturation temperature. Food Microbiol. 2021, 98, 103778. [Google Scholar] [CrossRef] [PubMed]
- Syromyatnikov, M.Y.; Kokina, A.V.; Solodskikh, S.A.; Panevina, A.V.; Popov, E.S.; Popov, V.N. High-throughput 16S rRNA gene sequencing of butter microbiota reveals a variety of opportunistic pathogens. Foods 2020, 9, 608. [Google Scholar] [CrossRef] [PubMed]
- Curioni, P.M.G.; Bosset, J.O. Key odorants in various cheese types as determined by gas chromatography-olfactometry. Int. Dairy J. 2002, 12, 959–984. [Google Scholar] [CrossRef]
- Cheng, H. Volatile flavor compounds in yogurt: A review. Crit. Rev. Food Sci. Nutr. 2010, 50, 938–950. [Google Scholar] [CrossRef]
- Lanciotti, E.; Patrignani, F.; Bagnolini, F.; Guerzoni, M.E.; Gardini, F. Evaluation of diacetyl antimicrobial activity against Escherichia coli, Listeria monocitogenes and Staphylococcus aureus. Food Microbiol. 2003, 20, 537–543. [Google Scholar] [CrossRef]
- Manafi, M.; Kneifel, W.; Bascomb, S. Fluorogenic and chromogenic substrates used in bacterial diagnostics. Microbiol. Rev. 1991, 55, 335–348. [Google Scholar]
- Feng, P.C.S.; Hartman, P.A. Fluorogenic assays for immediate confirmation of Escherichia coli. Appl. Environ. Microbiol. 1982, 43, 1320–1329. [Google Scholar] [CrossRef] [Green Version]
- IDF. Butter-Determination of Moisture, Non-Fat Solids and Fat Contents (Reference Method); IDF Standard 80; International Dairy Federation: Brussels, Belgium, 2021. [Google Scholar]
- Pirondini, U.; Bonas, E.; Maestri, G.; Visioli, M.; Marmiroli, N. Yield and amplificability of different DNA extraction procedures for traceability in the dairy food chain. Food Control 2010, 21, 663–668. [Google Scholar] [CrossRef]
- Baker, G.C.; Smith, J.J.; Cowan, D.A. Review and re-analysis of domain-specific 16S primers. J. Microbiol. Methods 2003, 55, 541–555. [Google Scholar] [CrossRef] [Green Version]
- Claesson, M.J.; Wang, Q.; O′sullivan, O.; Greene-Diniz, R.; Cole, J.R.; Ross, R.P.; O′Toole, P.W. Comparison of two next-generation sequencing technologies for resolving highly complex microbiota composition using tandem variable 16S rRNA gene regions. Nucleic Acids Res. 2010, 38, e200. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gokce, R.; Akdogan, A.; Divriklib, U.; Elci, L. Simultatenous determination of diacetyl and acetoin in traditional turkish butter stored in sheep’s rumen (Karinyagi). Grasas Aceites 2014, 65, e010. [Google Scholar]
- Idoui, T.; Benhamada, N.; Leghouchi, E. Microbial quality, physicochemical characteristics and fatty acid composition of a traditional butter produced from cows’ milk in East Algeria. Grasas Y Aceites 2010, 61, 232–236. [Google Scholar] [CrossRef] [Green Version]
- Samet-Bali, O.; Ayadi, M.A.; Attia, H. Traditional Tunisian butter: Physicochemical and microbial characteristics and storage stability of the oil fraction. LWT 2009, 42, 899–905. [Google Scholar] [CrossRef]
- Adam, M.R.; Moss, M.O. Food Microbiology, 1st ed.; RSC Publishing: Cambridge, UK, 1995. [Google Scholar]
- Holliday, S.L.; Adler, B.B.; Beuchat, L.R. Viability of Salmonella, Escherichia coli O157: H7, and Listeria monocytogenes in butter, yellow fat spreads, and margarine as affected by temperature and physical abuse. Food Microbiol. 2003, 20, 159–168. [Google Scholar] [CrossRef]
- Méndez-Cid, F.J.; Centeno, J.A.; Martínez, S.; Carballo, J. Changes in the chemical and physical characteristics of cow’s milk butter during storage: Effects of temperature and addition of salt. J. Food Compos. Anal. 2017, 63, 121–132. [Google Scholar] [CrossRef]
- McSweeney, P.L.H.; Fox, P.F. Metabolism of Residual Lactose and of Lactate and Citrate Cheese: Chemistry, Physics and Microbiology, 3rd ed.; Fox, P.F., McSweeney, P.L.H., Cogan, T.M., Guinee, T.P., Eds.; Chapman & Hall: London, UK, 2004; Volume 1, pp. 361–372. [Google Scholar]
- Sarhir, S.T.; Amanpour, A.; Bouseta, A.; Selli, S. Fingerprint of aroma-active compounds and odor activity values in a traditional Moroccan fermented butter “Smen” using GC–MS–Olfactometry. J. Food Compos. Anal. 2021, 96, 103761. [Google Scholar] [CrossRef]
- Liang, T.; Xie, X.; Zhang, J.; Ding, Y.; Wu, Q. Bacterial community and composition of different traditional fermented dairy products in China, South Africa, and Sri Lanka by high-throughput sequencing of 16S rRNA genes. LWT 2021, 144, 111209. [Google Scholar] [CrossRef]
- Carafa, I.; Castro Navarro, I.; Bittante, G.; Tagliapietra, F.; Gallo, L.; Tuohy, K.; Franciosi, E. Shift in the cow milk microbiota during alpine pasture as analyzed by culture dependent and high-throughput sequencing techniques. Food Microbiol. 2020, 91, 103504. [Google Scholar] [CrossRef] [PubMed]
- Gray, M.W. The evolutionary origins of organelles. Trends Genet. 1989, 5, 294–299. [Google Scholar] [CrossRef]
- Franciosi, E.; De Sabbata, G.; Gardini, F.; Cavazza, A.; Poznanski, E. Changes in psychrotrophic microbial populations during milk creaming to produce Grana Trentino cheese. Food Microbiol. 2011, 28, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, G.B.; de Favarin, L.; Luchese, R.H.; McIntosh, D. Psychrotrophic bacteria in milk: How much do we really know? Braz. J. Microbiol. 2015, 46, 313–321. [Google Scholar] [CrossRef]
- Perin, L.M. Intereference of storage temperatures in the development of mesophilic, psychrotrophic, lipolytic and proteolytic microbiota of raw milk. Semin. Ciências Agrárias 2012, 33, 333–342. [Google Scholar] [CrossRef]
- Fricker, M.; Skånseng, B.; Rudi, K.; Stessl, B.; Ehling-Schulz, M. Shift from farm to dairy tank milk microbiota revealed by a polyphasic approach is independent from geographical origin. Int. J. Food Microbiol. 2011, 145, S24–S30. [Google Scholar] [CrossRef]
- Franciosi, E.; Narduzzi, L.; Paradiso, A.; Carlin, S.; Tuohy, K.; Beretta, A.; Mattivi, F. Microbial community dynamics in phyto-thermotherapy baths viewed through next generation sequencing and metabolomics approach. Sci. Rep. 2020, 10, 17931. [Google Scholar] [CrossRef]
- Lafarge, V.; Ogier, J.-C.; Girard, V.; Maladen, V.; Leveau, J.-Y.; Gruss, A.; Delacroix- Buchet, A. Raw cow milk bacterial population shifts attributable to refrigeration. Appl. Environ. Microbiol. 2004, 70, 5644–5650. [Google Scholar] [CrossRef] [Green Version]
- Carafa, I.; Stocco, G.; Franceschi, P.; Summer, A.; Tuohy, K.M.; Bittante, G.; Franciosi, E. Evaluation of autochthonous lactic acid bacteria as starter and non-starter cultures for the production of Traditional Mountain cheese. Food Res. Int. 2019, 115, 209–218. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mfarm | TBC | Lactococci | Lactobacilli | Staphylococci | Coliforms | Enterococci | Moisture |
|---|---|---|---|---|---|---|---|
| A | 7.1 ± 0.59 b | 7.1 ± 0.73 c | 6.9 ± 1.58 bc | 4.2 ± 0.80 b | 6.3 ± 1.69 bc | 4.4 ± 0.15 b | 14.3 ± 0.18 b |
| B | 6.1 ± 0.43 a | 6.2 ± 0.66 b | 5.2 ± 0.25 a | 4.7 ± 0.61 bc | 4.3 ± 1.59 a | 4.4 ± 0.06 b | 16.8 ± 1.30 bc |
| C | 8.2 ± 0.85 c | 7.1 ± 1.08 bc | 5.9 ± 1.26 ab | 3.7 ± 1.83 ab | 5.3 ± 1.94 b | 3.9 ± 1.36 ab | 12.9 ± 0.41 ab |
| D | 6.7 ± 0.01 a | 6.4 ± 0.67 b | 5.6 ± 0.01 b | 2.7 ± 0.00 a | 3.9 ± 0.13 a | 3.5 ± 0.21 a | 17.5 ± 5.66 bc |
| E | 5.8 ± 0.25 a | 4.5 ± 0.28 a | 5.2 ± 0.66 a | 3.5 ± 0.25 a | 3.5 ± 1.67 a | 3.4 ± 0.32 a | 14.3 ± 0.60 bc |
| F | 6.8 ± 1.80 ab | 6.4 ± 1.86 bc | 4.4 ± 1.20 a | 3.8 ± 1.88 ab | 5.1 ± 1.18 b | 3.1 ± 1.38 ab | 13.0 ± 0.44 ab |
| G | 7.0 ± 0.80 b | 7.1 ± 0.95 c | 5.7 ± 0.11 b | 3.9 ± 0.18 b | 4.5 ± 0.63 a | 4.4 ± 0.67 b | 13.4 ± 0.86 ab |
| H | 7.9 ± 0.16 c | 6.8 ± 1.48 bc | 6.9 ± 0.46 c | 5.0 ± 1.64 bc | 6.9 ± 0.66 c | 5.0 ± 0.78 c | 9.3 ± 1.61 a |
| I | 7.0 ± 0.52 b | 6.2 ± 1.95 bc | 5.8 ± 0.20 b | 4.6 ± 1.77 bc | 6.5 ± 1.39 bc | 5.3 ± 0.46 c | 18.3 ± 3.34 c |
| L | 6.6 ± 0.99 ab | 6.6 ± 1.06 bc | 4.5 ± 0.97 a | 4.2 ± 0.85 b | 4.1 ± 1.37 a | 2.6 ± 0.60 a | 13.9 ± 0.93 b |
| M | 7.3 ± 0.39 b | 6.3 ± 1.05 bc | 5.9 ± 0.52 b | 2.9 ± 1.79 ab | 6.4 ± 1.57 bc | 4.7 ± 1.97 bc | 12.0 ± 0.91 ab |
| N | 7.6 ± 0.28 b | 6.9 ± 1.28 bc | 6.8 ± 0.48 c | 3.6 ± 0.58 ab | 6.8 ± 1.00 bc | 4.6 ± 0.38 b | 11.6 ± 0.84 ab |
| O | 7.5 ± 0.23 b | 7.1 ± 0.12 c | 7.0 ± 0.17 c | 5.8 ± 1.08 c | 6.9 ± 0.03 c | 5.5 ± 0.01 c | 12.4 ± 0.06 ab |
| P | 6.5 ± 0.78 ab | 6.6 ± 1.08 bc | 4.9 ± 0.14 a | 3.0 ± 1.73 ab | 4.0 ± 1.16 a | 2.8 ± 0.74 a | 18.6 ± 1.29 c |
| TOT | 6.8 ± 0.88 | 5.6 ± 1.62 | 5.5 ± 1.15 | 2.9 ± 1.79 | 4.5 ± 1.10 | 3.7 ± 1.54 | 14.1 ± 3.1 |
| MFarm | Diacetyl | Acetoin |
|---|---|---|
| A | 0.203 ± 0.122 b | 5.95 ± 4.66 b |
| B | 0.085 ± 0.007 a | 0.25 ± 0.09 a |
| C | 0.090 ± 0.000 a | 0.35 ± 0.22 a |
| D | 0.095 ± 0.007 a | 0.59 ± 0.00 a |
| E | 0.085 ± 0.021 a | 0.20 ± 0.01 a |
| F | 0.090 ± 0.000 a | 0.49 ± 0.47 a |
| G | 0.090 ± 0.014 a | 0.59 ± 0.63 a |
| H | 0.317 ± 0.025 c | 63.88 ± 4.17 c |
| I | 0.190 ± 0.139 bc | 7.09 ± 6.87 b |
| L | 0.173 ± 0.006 b | 5.14 ± 1.60 b |
| M | 0.087 ± 0.015 a | 0.24 ± 0.12 a |
| N | 0.310 ± 0.050 c | 32.84 ± 9.38 c |
| O | 0.175 ± 0.064 b | 7.36 ± 2.27 b |
| P | 0.853 ± 0.399 d | 32.60 ± 8.42 c |
| TOT | 0.226 ± 0.239 | 13.9 ± 24.4 |
| MFarm | ObsOTUs | Shannon’s Index |
|---|---|---|
| A | 403 ± 171 | 7.75 ± 0.808 |
| B | 266 ± 176 | 6.94 ± 0.310 |
| C | 490 ± 197 | 8.17 ± 0.563 |
| D | 395 ± 28 | 7.83 ± 0.053 |
| E | 356 ± 194 | 7.82 ± 0.915 |
| F | 369 ± 118 | 7.58 ± 0.514 |
| G | 449 ± 64 | 7.91 ± 0.021 |
| H | 464 ± 121 | 8.10 ± 0.640 |
| I | 396 ± 24 | 7.98 ± 0.189 |
| L | 475 ± 56 | 8.21 ± 0.178 |
| M | 420 ± 101 | 7.70 ± 0.842 |
| N | 459 ± 120 | 8.07 ± 0.595 |
| O | 445 ± 27 | 7.86 ± 0.390 |
| P | 419 ± 40 | 7.97 ± 0.302 |
| TOT | 418 ± 116 | 7.87 ± 0.613 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schiavon, S.; Paolini, M.; Guzzon, R.; Mancini, A.; Larcher, R.; Villegas, T.R.; Franciosi, E. Bacterial Complexity of Traditional Mountain Butter Is Affected by the Malga-Farm of Production. Microorganisms 2022, 10, 17. https://doi.org/10.3390/microorganisms10010017
Schiavon S, Paolini M, Guzzon R, Mancini A, Larcher R, Villegas TR, Franciosi E. Bacterial Complexity of Traditional Mountain Butter Is Affected by the Malga-Farm of Production. Microorganisms. 2022; 10(1):17. https://doi.org/10.3390/microorganisms10010017
Chicago/Turabian StyleSchiavon, Silvia, Mauro Paolini, Raffaele Guzzon, Andrea Mancini, Roberto Larcher, Tomas Roman Villegas, and Elena Franciosi. 2022. "Bacterial Complexity of Traditional Mountain Butter Is Affected by the Malga-Farm of Production" Microorganisms 10, no. 1: 17. https://doi.org/10.3390/microorganisms10010017