Trypanosomatid Infections among Vertebrates of Chile: A Systematic Review

Abstract
:
1. Introduction
2. Results
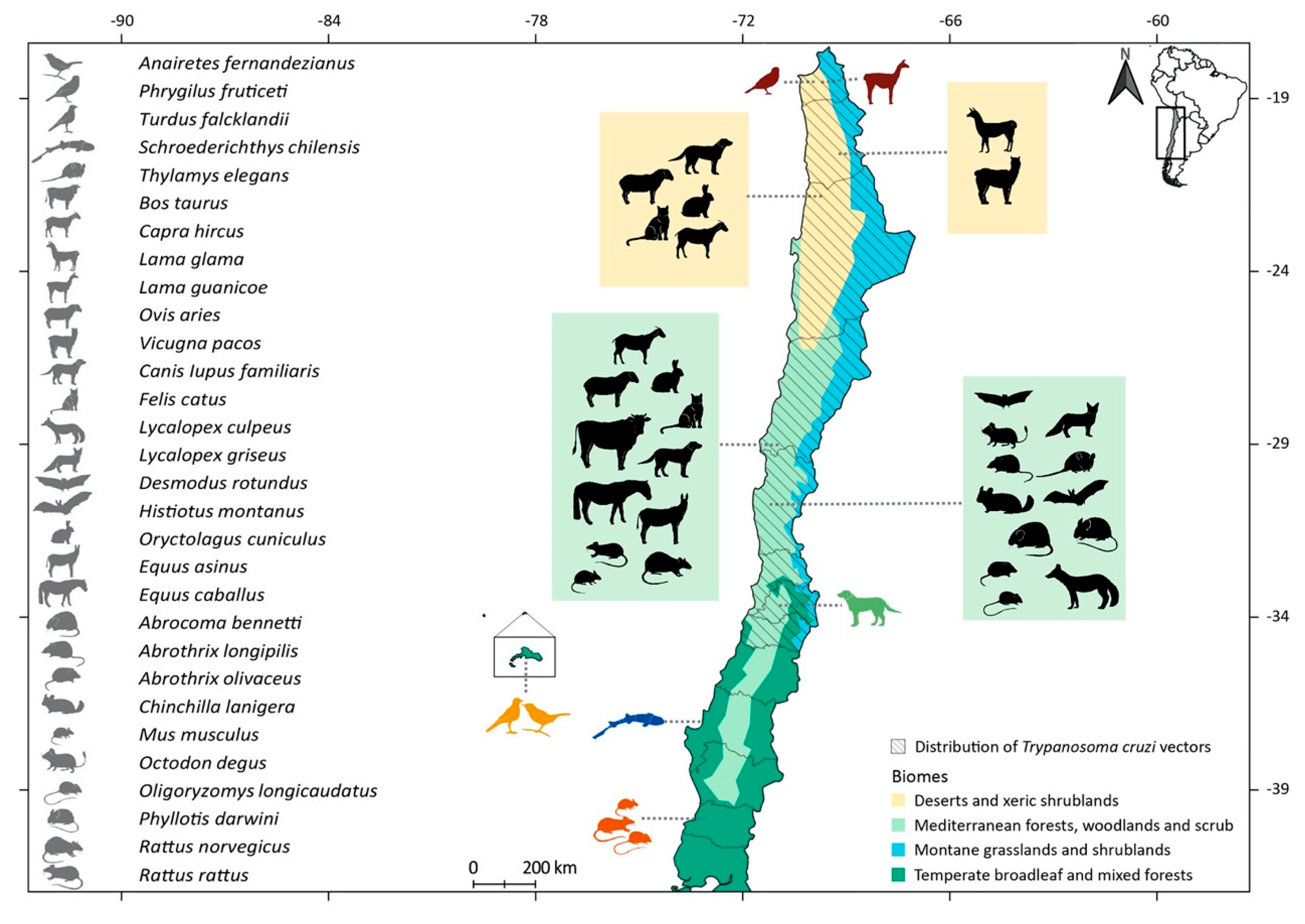
2.1. Native Mammal Hosts
2.1.1. Order Didelphimorphia
2.1.2. Order Artiodactyla
2.1.3. Order Carnivora
2.1.4. Order Chiroptera
2.1.5. Order Rodentia
2.2. Alien Mammal Hosts
2.2.1. Order Artiodactyla
2.2.2. Order Carnivora
2.2.3. Order Lagomorpha
2.2.4. Order Perissodactyla
2.2.5. Order Rodentia
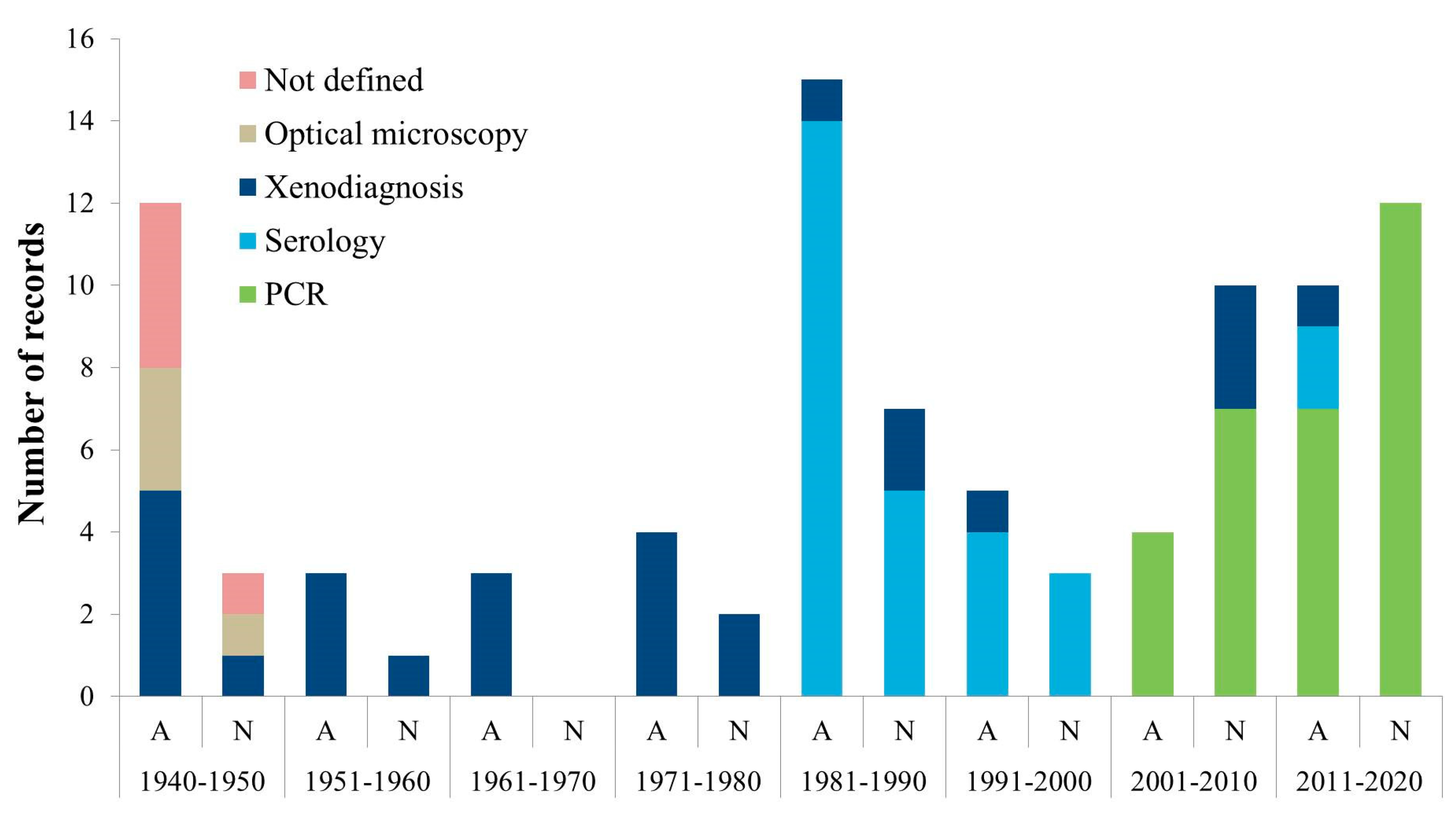
3. Discussion
4. Materials and Methods
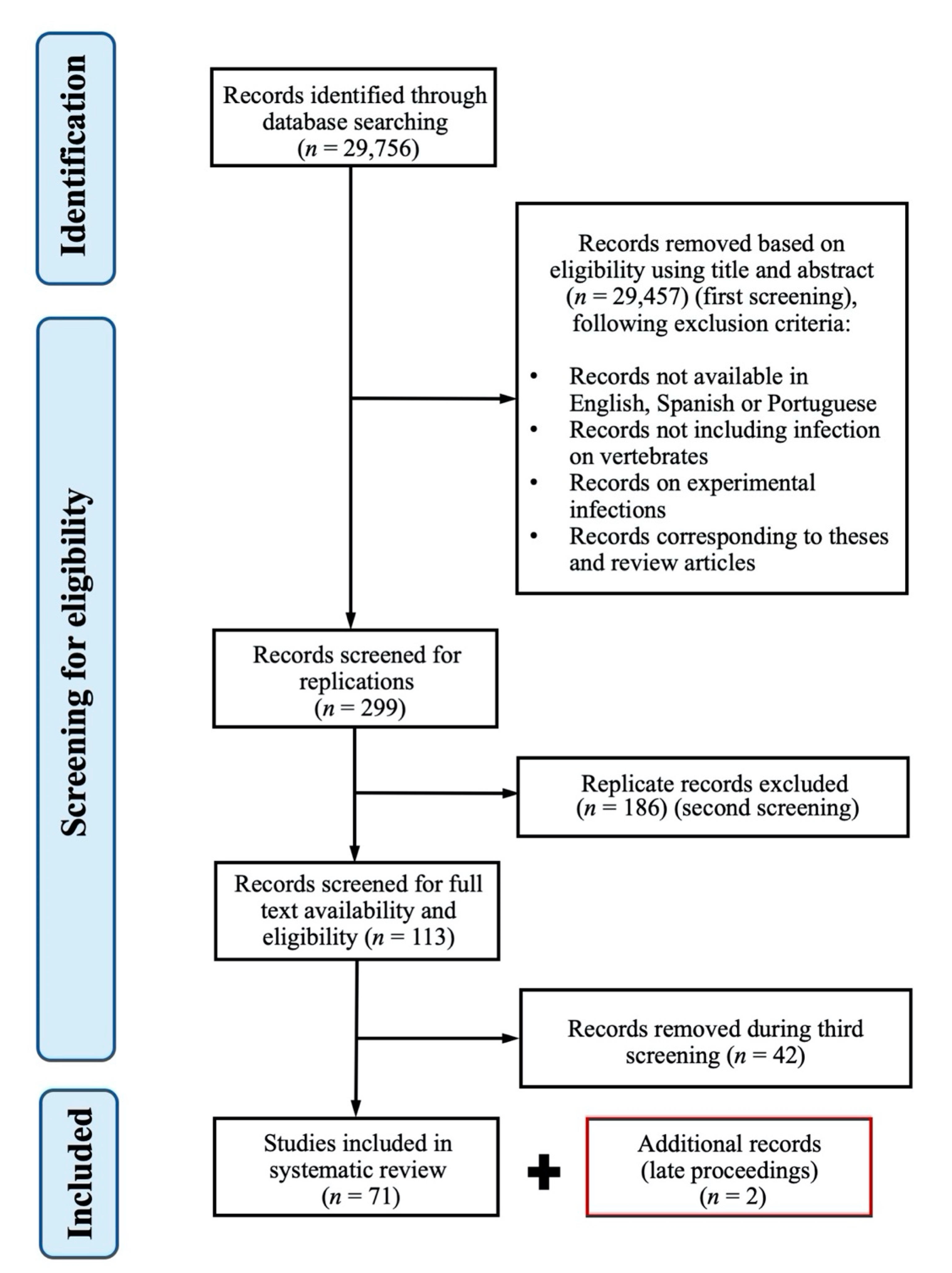
4.1. Systematic Review Protocol, Search Strategy and Data Collection
4.2. Study Selection
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- National Center for Biotechnology Information (NCBI) [Internet]. Bethesda (MD): National Library of Medicine (US), National Center for Biotechnology Information. Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 20 May 2020).
- Kaufer, A.; Ellis, J.; Stark, D.; Barratt, J. The evolution of trypanosomatid taxonomy. Parasit. Vectors 2017, 10, 287. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Research Priorities for Chagas Disease, Human African Trypanosomiasis and Leishmaniasis; WHO Technical Report Series Nº 975: Geneva, Italy, 2012. [Google Scholar]
- Roche, B.; Broutin, H.; Simard, F. Ecology and Evolution of Infectious Diseases: Pathogen Control and Public Health Management in Low-Income Countries; Oxford University Press: Oxford, UK, 2018. [Google Scholar]
- Dinerstein, E.; Olson, D.; Joshi, A.; Vynne, C.; Burgess, N.D.; Wikramanayake, E.; Hahn, N.; Palminteri, S.; Hedao, P.; Noss, R.; et al. An ecoregion-based approach to protecting half the terrestrial realm. Bioscience 2017, 67, 534–545. [Google Scholar] [CrossRef] [PubMed]
- Ministerio del Medio Ambiente (MMA). Biodiversidad de Chile; Patrimonio y Desafíos. Tercera Edición; Tomo I, 430 pages: Santiago, Chile, 2018; Available online: https://mma.gob.cl/wp-content/uploads/2019/04/Tomo-I-libro-Biodiversidad-Chile-MMA-web.pdf (accessed on 15 May 2020).
- Jaksic, F.M. Vertebrate invaders and their ecological impacts in Chile. Biodivers. Conserv. 1998, 7, 1427–1445. [Google Scholar] [CrossRef]
- Taraschewski, H. Hosts and parasites as aliens. J. Helminthol. 2006, 80, 99–128. [Google Scholar] [CrossRef]
- Botto-Mahan, C.; Sepúlveda, M.; Vidal, M.; Acuña-Retamar, M.; Ortiz, S.; Solari, A. Trypanosoma cruzi infection in the wild kissing bug Mepraia gajardoi from the Chilean Southern Pacific Ocean coast. Acta Trop. 2008, 105, 166–169. [Google Scholar] [CrossRef]
- Bacigalupo, A.; Torres-Pérez, F.; Segovia, V.; García, A.; Correa, J.P.; Moreno, L.; Arroyo, P.; Cattan, P.E. Sylvatic foci of the Chagas disease vector Triatoma infestans in Chile: Description of a new focus and challenges for control programs. Mem. Inst. Oswaldo Cruz 2010, 105, 633–641. [Google Scholar] [CrossRef] [Green Version]
- Rives-Blanchard, N.; Torres-Pérez, F.; Ortiz, S.; Solari, A.; Campos-Soto, R. Trypanosoma cruzi over the ocean: Insular zones of Chile with presence of infected vector Mepraia species. Acta Trop. 2017, 172, 229–231. [Google Scholar] [CrossRef]
- Tapia-Garay, V.; Figueroa, D.P.; Maldonado, A.; Frías-Laserre, D.; González, C.R.; Parra, A.; Canals, L.; Apt, W.; Alvarado, S.; Cáceres, D.; et al. Assessing the risk zones of Chagas’ disease in Chile, in a world marked by global climatic change. Mem. Inst. Oswaldo Cruz 2018, 113, 24–29. [Google Scholar] [CrossRef] [Green Version]
- Garrido, R.; Bacigalupo, A.; Peña-Gómez, F.; Bustamante, R.O.; Cattan, P.E.; Gorla, D.; Botto-Mahan, C. Potential impact of climate change on geographical distribution of two wild vectors of Chagas disease in Chile: Mepraia spinolai and Mepraia gajardoi. Parasit. Vectors 2019, 12, 478. [Google Scholar] [CrossRef]
- Ihle-Soto, C.; Costoya, E.; Correa, J.P.; Bacigalupo, A.; Cornejo-Villar, B.; Estadella, V.; Solari, A.; Ortiz, S.; Hernández, H.J.; Botto-Mahan, C.; et al. Spatio-temporal characterization of Trypanosoma cruzi infection and discrete typing units infecting hosts and vectors from non-domestic foci of Central Chile. PLoS Negl. Trop. Dis. 2019, 13, e7170. [Google Scholar] [CrossRef]
- Schenone, H.; Villarroel, F.; Rojas, A.; Alfaro, E. Biological and ecological factors in the epidemiology of Chagas’ disease in Chile. Bol. Chil. Parasitol. 1980, 35, 42–54. [Google Scholar] [PubMed]
- Schenone, H.; Contreras, M.C.; Borgono, J.M.; Maturana, R.; Salinas, P.; Sandoval, L.; Rojas, A.; Tello, P.; Villarroel, F. Overview of the epidemiology of Chagas’ disease in Chile. Bol. Chil. Parasitol. 1991, 46, 19–30. [Google Scholar] [PubMed]
- Rozas, M.; Botto-Mahan, C.; Coronado, X.; Ortiz, S.; Cattan, P.; Solari, A. Trypanosoma cruzi infection in wild mammals from a Chagasic area of Chile. Am. J. Trop. Med. Hyg. 2005, 73, 517–519. [Google Scholar] [CrossRef] [PubMed]
- Cabello, J.; Altet, L.; Napolitano, C.; Sastre, N.; Hidalgo, E.; Davila, J.A.; Millan, J. Survey of infectious agents in the endangered Darwin’s fox (Lycalopex fulvipes): High prevalence and diversity of hemotrophic mycoplasmas. Vet. Microbiol. 2013, 167, 448–454. [Google Scholar] [CrossRef]
- Cevidanes, A.; Cataldo, S.D.; Muñoz-San Martín, C.; Hernández, C.; Latrofa, M.S.; Cattan, P.; Otranto, D.; Millán, J. Canine vector-borne pathogens in rural dogs in Chile: Molecular survey and co-infection patterns. Unpublished (Pre-print available). [CrossRef]
- Forrester, D.J.; Greiner, E.C.; McFarlane, R.W. Blood parasites of some columbiform and passeriform birds from Chile. J. Wildl. Dis. 1977, 13, 94–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortiz, S.; Marín, J.C.; Correa, J.P.; Solari, A. Molecular characterization of a Trypanosomatid circulating in guanacos (Lama guanicoe Müller 1776) from Putre and Magallanes, Chile. Parasitol. Lat. 2020, 69, 79. [Google Scholar]
- Martinez, J.; Vasquez, R.A.; Marques, A.; Diez-Fernandez, A.; Merino, S. The prevalence and molecular characterization of blood parasites infecting the vulnerable Tamarugo Conebill (Conirostrum tamarugense) and other birds in the Pampa del Tamarugal, Chile. Emu-Austral Ornithol. 2016, 116, 310–314. [Google Scholar] [CrossRef] [Green Version]
- Martinez, J.; Vasquez, R.A.; Venegas, C.; Merino, S. Molecular characterisation of haemoparasites in forest birds from Robinson Crusoe Island: Is the Austral Thrush a potential threat to endemic birds? Bird Conserv. Int. 2015, 25, 139–152. [Google Scholar] [CrossRef] [Green Version]
- Morillas, J.; George-Nascimento, M.; Valeria, H.; Khan, R.A. Trypanosoma humboldti sp. from the Chilean Catshark, Schroederichthys chilensis (Guichenot, 1848). J. Protozool. 1987, 34, 342–344. [Google Scholar] [CrossRef]
- Valenzuela, A.; Oyarzún, C.; Silva, V. Blood cells of the Schroederichthys chilensis (Guichenot 1848): The leukocytes (Elasmobranchii, Scyliorhinidae). Gayana 2003, 67, 130–137. [Google Scholar] [CrossRef] [Green Version]
- Silva, V.; Valenzuela, A.; Ruiz, P.; Oyarzún, C. Trypanosoma humboldti in Schroederichthys chilensis (Chondrichthyes, Elasmobranchii, Scyliorhinidae) as non destructive indicator of contamination. Gayana 2005, 69, 160–165. [Google Scholar] [CrossRef] [Green Version]
- Franjola, R.; Soto, G.; Montefusco, A. Prevalence of protozoa infections in synanthropic rodents in Valdivia City, Chile. Bol. Chil. Parasitol. 1995, 50, 66–72. [Google Scholar] [PubMed]
- Iriarte, A. Mamíferos de Chile; Lynx Edicions: Barcelona, Spain, 2008; p. 420. [Google Scholar]
- Whiting, C. Contribución al estudio de las reservas de parásitos de la enfermedad de Chagas en Chile. Primeros hallazgos en Chile de mamíferos silvestres infestados por Trypanosoma cruzi. Rev. Chil. Hig. Med. Prev. 1946, 8, 69–100. [Google Scholar]
- Durán, J.; Videla, M.; Apt, W. Chagas disease in a community of small sympatric mammals from Las Chinchillas National Reserve, IV Region, Chile. Parasitol. Día 1989, 13, 15–20. [Google Scholar]
- Rozas, M.; Botto-Mahan, C.; Coronado, X.; Ortiz, S.; Cattan, P.E.; Solari, A. Co-existence of Trypanosoma cruzi genotypes in wild and peridomestic mammals in Chile. Am. J. Trop. Med. Hyg. 2007, 77, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Botto-Mahan, C.; Campos, R.; Acuña-Retamar, M.; Coronado, X.; Cattan, P.; Solari, A. Temporal variation of Trypanosoma cruzi infection in native mammals in Chile. Vector-Borne Zoonot. Dis. 2010, 10, 317–319. [Google Scholar] [CrossRef] [PubMed]
- Correa, J.P.; Bacigalupo, A.; Fontúrbel, F.; Oda, E.; Cattan, P.E.; Solari, A.; Botto-Mahan, C. Spatial distribution of an infectious disease in a native small mammal community. Sci. Nat. 2015, 102, 51. [Google Scholar] [CrossRef]
- Bacigalupo, A.; Ihle-Soto, C.; Costoya, E.; Correa, J.P.; Cornejo-Villar, B.; Estadella, V.; Solari, A.; Ortiz, S.; Botto-Mahan, C.; Cattan, P.E. Spatio-temporal distribution of Trypanosoma cruzi in foci of the North-Central zone of Chile. Parasitol. Lat. 2018, 67, 18–19. [Google Scholar]
- Zingales, B.; Andrade, S.G.; Briones, M.R.S.; Campbell, D.A.; Chiari, E.; Fernandes, O.; Guhl, F.; Lages-Silva, E.; Macedo, A.M.; Machado, C.R.; et al. A new consensus for Trypanosoma cruzi intraspecific nomenclature: Second revision meeting recommends TcI to TcVI. Mem. Inst. Oswaldo Cruz 2009, 104, 1051–1054. [Google Scholar] [CrossRef]
- Martínez, R.; Ahumada, C.; Contreras, M.C.; Villarroel, F.; Rojas, A.; Schenone, H. Chagas’ disease in Chile. Rural Sectors. Domiciliary triatomid infestation and Trypanosoma cruzi infection of the vector and mammals of Region I (1982–1983). Bol. Chil. Parasitol. 1983, 38, 70–72. [Google Scholar]
- Burchard, L.; Cornejo, J.; Cruz, L.; Contreras, M.C.; Vargas, L.; Villarroel, F.; Rojas, A.; Schenone, H. Epidemiology of Chagas’ disease in Chile. Rural sectors. Domiciliary triatomid infestation and Trypanosoma cruzi infection of the vector and domestic mammals of Region II (1983). Bol. Chil. Parasitol. 1984, 39, 17–19. [Google Scholar] [PubMed]
- Lorca, M.; Campano, S.; Meyer-Kayser, E. Trypanosoma cruzi infection in South American camelids. Parasitol. Día 1991, 15, 52–54. [Google Scholar]
- Villarroel, F.; Schenone, H.; Contreras, M.C.; Rojas, A.; Hernandez, E. Chagas disease in the Chilean altiplano. Epidemiological, parasitological, and clinical aspects. Bol. Chil. Parasitol. 1991, 46, 61–69. [Google Scholar] [PubMed]
- Neghme, A.; Román, J. Present state of Chagas’ disease surveys in Chile. Am. J. Trop. Med. Hyg. 1948, 835–839. [Google Scholar] [CrossRef] [PubMed]
- Neghme, A.; Román, J.; Sotomayor, R. New data on Chagas’ disease in Chile. Bol. Oficina Sanit. Panam. 1949, 28, 808–817. [Google Scholar] [PubMed]
- Neghme, A.R.; Schenone, H. Resumen de veinte años de investigación sobre la enfermedad de Chagas en Chile. Rev. Med. Chile 1960, 88, 82–93. [Google Scholar]
- Schenone, H. The status of epidemiological research in Chile on Chagas’s disease. Bol. Oficina Sanit. Panam. 1971, 70, 250–255. [Google Scholar]
- Jiménez, J.; Lorca, M. American trypanosomiasis in sylvatic vertebrates and its relation to the vector Triatoma spinolai. Arch. Med. Vet. 1990, 22, 179–183. [Google Scholar]
- Correa, J.P.; Quiroga, N.; Campos-Soto, R.; Díaz-Campusano, G.; Yañez-Meza, A.; Allendes, J.L.; Rodríguez-San Pedro, A.; Botto-Mahan, C. Detection of Trypanosoma cruzi DNA in bats from two protected areas of central-northern Chile: Preliminary data. Parasitol. Lat. 2020, 69, 76. [Google Scholar]
- Campos, R.; Botto-Mahan, C.; Ortiz, S.; Acuña, M.; Cattan, P.; Solari, A. Trypanosoma cruzi detection in blood by xenodiagnosis and polymerase chain reaction in the wild rodent Octodon degus. Am. J. Trop. Med. Hyg. 2007, 76, 324–326. [Google Scholar] [CrossRef]
- Campos, R.; Acuña-Retamar, M.; Botto-Mahan, C.; Ortiz, S.; Cattan, P.; Solari, A. Susceptibility of Mepraia spinolai and Triatoma infestans to different Trypanosoma cruzi strains from naturally infected rodent hosts. Acta Trop. 2007, 104, 25–29. [Google Scholar] [CrossRef]
- Galuppo, S.; Bacigalupo, A.; García, A.; Ortiz, S.; Coronado, X.; Cattan, P.E.; Solari, A. Predominance of Trypanosoma cruzi genotypes in two reservoirs infected by sylvatic Triatoma infestans of an endemic area of Chile. Acta Trop. 2009, 111, 90–93. [Google Scholar] [CrossRef] [PubMed]
- Campos, R.; Botto-Mahan, C.; Ortiz, S.; Coronado, X.; Solari, A. Temporal fluctuation of infection with different Trypanosoma cruzi genotypes in the wild rodent Octodon degus. Am. J. Trop. Med. Hyg. 2010, 83, 380–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botto-Mahan, C.; Bacigalupo, A.; Correa, J.P.; Oda, E.; Solari, A. Field assessment of Trypanosoma cruzi infection and host survival in the native rodent Octodon degus. Acta Trop. 2012, 122, 164–167. [Google Scholar] [CrossRef] [PubMed]
- Oda, E.; Solari, A.; Botto-Mahan, C. Effects of mammal host diversity and density in the infection level of a sylvatic kissing bug. Med. Vet. Entomol. 2014, 28, 384–390. [Google Scholar] [CrossRef] [PubMed]
- Botto-Mahan, C.; Rojo, G.; Sandoval-Rodríguez, A.; Peña, F.; Ortiz, S.; Solari, A. Temporal variation in Trypanosoma cruzi lineages in the native rodent Octodon degus in semiarid Chile. Acta Trop. 2015, 151, 178–181. [Google Scholar] [CrossRef]
- Jiménez, C.; Fontúrbel, F.; Oda, E.; Ramírez, P.; Botto-Mahan, C. Parasitic infection alters rodent movement in a semiarid ecosystem. Mamm. Biol. 2015, 80, 255–259. [Google Scholar] [CrossRef]
- Donoso, M.I.; Cares, R.A.; Fontúrbel, F.; Oda, E.; Ramírez, P.A.; Botto-Mahan, C. Temporal fluctuation in shrub species preferences by two native rodents: The effect of infection status on habitat use. Austral. Ecol. 2016, 41, 512–518. [Google Scholar] [CrossRef]
- Rojo, G.; Sandoval-Rodríguez, A.; López, A.; Ortiz, S.; Correa, J.P.; Saavedra, M.; Botto-Mahan, C.; Cattan, P.E.; Solari, A. Within-host temporal fluctuations of Trypanosoma cruzi discrete typing units: The case of the wild reservoir rodent Octodon degus. Parasit. Vectors 2017, 10, 380. [Google Scholar] [CrossRef] [Green Version]
- Yefi-Quinteros, E.; Muñoz-San Martín, C.; Bacigalupo, A.; Correa, J.P.; Cattan, P.E. Trypanosoma cruzi load in synanthropic rodents from rural areas in Chile. Parasit. Vectors 2018, 11, 171. [Google Scholar] [CrossRef] [Green Version]
- Botto-Mahan, C.; Bacigalupo, A.; Correa, J.P.; Fontúrbel, F.E.; Cattan, P.E.; Solari, A. Prevalence, infected density or individual probability of infection? Assessing vector infection risk in the wild transmission of Chagas disease. Proc. R. Soc. B 2020, 287, 20193018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez, C.; Stagno, S.; Welch, E.; Villarroel, F.; Rojas, A.; Schenone, H. Human and animal Chagas’ infection in dwellings previously sprayed with insecticides. Bol. Chil. Parasitol. 1970, 25, 33–36. [Google Scholar] [PubMed]
- Correa, V.; Briceno, J.; Zuniga, J. Trypanosoma cruzi infection in domestic animals in rural sections of the IV Region, Chile. Bol. Chil. Parasitol. 1982, 37, 27–28. [Google Scholar] [PubMed]
- Bertoglia, J.; Rodríguez, J.; Gordillo, N.; Mendoza, J.; Contreras, M.C.; Rojas, J.; Rojas, A.; Villarroel, F.; Schenone, H. Epidemiology of Chagas’ disease in Chile. Rural sectors. Infection of domestic mammals by Trypanosoma cruzi and new contributions to the knowledge of domiciliary triatomid infestation in Region III, Chile (1982–1983). Bol. Chil. Parasitol. 1984, 39, 20–23. [Google Scholar]
- Correa, V.; Zúñiga, J.; Briceño, J.; Contreras, M.C.; Aranda, J.C.; Valdés, J.; Rojas, A.; Villarroel, F.; Schenone, H. Epidemiology of Chagas’ disease in Chile. Rural sections. Domiciliary infestation by Triatominae, rates of Trypanosoma cruzi infection in these and new contributions to the knowledge of Chagas infection in domestic mammals from Region IV (1982–1984). Bol. Chil. Parasitol. 1984, 39, 24–27. [Google Scholar]
- Flores, B.; Hernández, G.; Lepe, A.; Contreras, M.C.; Sandoval, L.; Villarroel, F.; Rojas, A.; González, O.; Schenone, H. Epidemiology of Chagas’ disease in Chile. Rural sectors. Domiciliary triatomid infestation and Trypanosoma cruzi infection of the vector and domestic mammals in Region V 1983. Bol. Chil. Parasitol. 1984, 39, 62–65. [Google Scholar]
- Villarroel, F.; Rojas, A.; Contreras, M.C.; Schenone, H. Epidemiology of Chagas’ disease in Chile. Rural sectors. Domiciliary triatomid infestation and Trypanosoma cruzi infection of vectors and domestic mammals of the Metropolitan Region. Bol. Chil. Parasitol. 1984, 39, 65–68. [Google Scholar]
- Venegas, L.; Rojas, A.; Villarroel, F.; Contreras, M.C.; Sandoval, L.; Schenone, H. Epidemiology of Chagas’ disease in Chile. Rural sectors. Domiciliary triatomid infestation and Trypanosoma cruzi infection of the vector and domestic mammals of the Bernardo O’Higgins Region VI, 1983. Bol. Chil. Parasitol. 1984, 39, 69–72. [Google Scholar]
- Schenone, H.; Contreras, M.C.; Borgoño, J.M.; Rojas, A.; Villarroel, F.; Valdés, J. Chagas’ disease in Chile. Rural and periurban sectors of the endemo-enzootic area. Relationship between housing conditions, domiciliary triatomid infestation and infection by Trypanosoma cruzi of the vector, humans and domestic mammals. 1982–1985. Bol. Chil. Parasitol. 1985, 40, 58–67. [Google Scholar]
- Ríos, A.; Alcaíno, H.; Apt, W. Chagas’s disease in synanthropic dogs, cattle and equines of the Limarí Province, Chile. Parasitol. Día 1986, 10, 40–45. [Google Scholar]
- Donckaster, R.; Neumann, G.; Gajardo, G.; Díaz, J. Evaluación de un programa de erradicación del triatomino domiciliario en el Valle de Elqui. Bol. Chil. Parasitol. 1962, 17, 2–6. [Google Scholar] [PubMed]
- Schenone, H.; Villarroel, F.; Alfaro, E. Epidemiology of Chagas disease in Chile. Housing conditions related to the presence of Triatoma infestans and the rate of humans and animals infected by Trypanosoma cruzi. Bol. Chil. Parasitol. 1978, 33, 2–7. [Google Scholar] [PubMed]
- Alcaino, H.A.; Arrau, S.A.; Apt, W.; Ríos, A. Chagas’ disease in synanthropic goats of the Limari Province, Chile. Rev. Med. Chile 1982, 110, 328–332. [Google Scholar] [PubMed]
- Ulloa, M.; Traslaviña, M.; Alcaino, H.; Apt, W.; Sandoval, J. Enfermedad de Chagas en caninos y caprinos sinantrópicos de la provincia del Choapa (IV Región), Chile. Parasitol. Día 1989, 13, 120–124. [Google Scholar]
- Alcaíno, T.V.; Lorca Herrera, M.; Nuñez, F.; Issotta, A.; Gorman, T. Chagas’ disease in goats from the Metropolitan Region, Chile: Seroepidemiological survey and experimental infection. Parasitol. Día 1995, 19, 30–36. [Google Scholar]
- Aguilera, C.; Zulantay, I.; Saavedra, M.; Apt, W.; Martinez, G.; Rodriguez, J. Economic loss by goats infected with Trypanosoma cruzi. IV Region, Coquimbo, Chile. Parasitol. Lat. 2015, 64, 23–31. [Google Scholar]
- Gasic, G.; Bertin, V. Animal reservoirs of the virus of American Trypanosomiasis in Chile. Rev. Chil. Hig. Med. Prev. 1940, 2, 247–261. [Google Scholar]
- Gasic, G.; Bertin, V. The epidemiology of Chagas’ disease in Chile. Rev. Chil. Pediatr. 1940, 11, 561–583. [Google Scholar]
- Gasic, G.; Carvajal, V. Chagas’s disease in Chile. Symptomatology and epidemiology. Rev. Med. Chile 1941, 69, 818–833. [Google Scholar]
- Gasic, G. Contribution to the study of Chagas’ disease in Chile. Proc. 8th Amer. Sci. Congr. 1942, 6, 239. [Google Scholar]
- Gasic, G. Facts regarding Chagas’s disease in Chile. Bol. Oficina Sanit. Panam. 1943, 22, 327–335. [Google Scholar]
- Badínez, O. Encuestas rurales sobre enfermedad de Chagas en la Provincia de Santiago. Rev. Hig. Med. Prev. 1943, 6, 1–9. [Google Scholar]
- Pino, F.; Whiting, C.; Alee, K.; Román, J. Una experiencia sanitaria rural. II. Encuesta epidemiológica de enfermedad de Chagas en el distrito sanitario experimental de Pirque (Provincia de Santiago). Rev. Chil. Hig. Med. Prev. 1953, 15, 65–71. [Google Scholar] [PubMed]
- Forgacs, E.; Schenone, H.; Niedmann, G. Encuesta sobre enfermedad de Chagas en el pueblo minero Inca de Oro. Bol. Chil. Parasitol. 1957, 12, 51–52. [Google Scholar] [PubMed]
- Schenone, H.; Ramírez, M.; Reyes, H.; Rojas, A.; Díaz, L. Contribution to the epidemiology of Chagas disease in Chile. Epidemiological survey in Colchagua Province. Bol. Chil. Parasitol. 1966, 21, 66–69. [Google Scholar]
- Rojas, A.; Sotelo, J.M.; Villarroel, F.; Contreras, M.C. The importance of dogs and cats in the epidemiology of Chagas disease. Bol. Chil. Parasitol. 1973, 28, 42–43. [Google Scholar]
- Rodríguez, J.; Bertoglia, J.; Gordillo, N.; Mendoza, J.; Rojas, J.; Contreras, M.C.; Schenone, H.; Villarroel, F.; Rojas, A. Domiciliary Triatomidae infestation and Trypanosoma cruzi infection in Region III of Atacama, Chile. Bol. Chil. Parasitol. 1982, 37, 29–30. [Google Scholar]
- Burchard, L.; Cáceres, J.; Sagua, H.; Bahamonde, M.I.; Neira, I.; Araya, J.; Goycolea, M. Present status of seroprevalence of canine and human chagasic infection in San Pedro de Atacama, II Región, Antofagasta, Chile 1995. Bol. Chil. Parasitol. 1996, 51, 76–79. [Google Scholar]
- González, C.R.; Reyes, C.; Canals, A.; Parra, A.; Munoz, X.; Rodriguez, K. An entomological and seroepidemiological study of the vectorial-transmission risk of Chagas disease in the coast of northern Chile. Med. Vet. Entomol. 2015, 29, 387–392. [Google Scholar] [CrossRef]
- Opazo, A.; Urrutia, S.; Bacigalupo, A. Determination of Trypanosoma cruzi infection in dogs and their ticks using polymerase chain reaction in Chile. In Proceedings of the 41st World Small Animal Veterinary Association Congress, Cartagena, Colombia, 27–30 September 2016; p. 816. [Google Scholar]
- Ortiz, S.; Ceballos, M.J.; González, C.R.; Reyes, C.; Gómez, V.; García, A.; Solari, A. Trypanosoma cruzi diversity in infected dogs from areas of the north coast of Chile. Vet. Parasitol. Reg. Stud. Rep. 2016, 5, 42–47. [Google Scholar] [CrossRef]
- Botto-Mahan, C.; Acuña-Retamar, M.; Campos, R.; Cattan, P.; Solari, A. European rabbits (Oryctolagus cuniculus) are naturally infected with different Trypanosoma cruzi genotypes. Am. J. Trop. Med. Hyg. 2009, 80, 944–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarkson, M.J. Trypanosomosis of domesticated animals of South America. Trans. Roy. Soc. Trop. Med. Hyg. 1976, 70, 125–126. [Google Scholar] [CrossRef]
- Desquesnes, M. Livestock Trypanosomoses and Their Vectors in Latin America; CIRAD-EMVT Publication: OIE, Paris, France, 2004; Available online: http://www.oie.int/doc/ged/D9818 (accessed on 1 March 2020).
- Gajardo-Tobar, R.; Thiermann, E. Discovery of a new Trypanosoma in Chile. Bol. Inf. Parasit. Chil. 1950, 5, 23–25. [Google Scholar]
- Ministerio del Medio Ambiente (MMA). Inventario Nacional de Especies de Chile. Available online: http://especies.mma.gob.cl/CNMWeb/Web/WebCiudadana/Default.aspx (accessed on 1 March 2020).
- Jones, T.W.; Dávila, A.M.R. Trypanosoma vivax–out of Africa. Trends Parasitol. 2001, 17, 99–101. [Google Scholar] [CrossRef]
- Aregawi, W.G.; Agga, G.E.; Abdi, R.D.; Büscher, P. Systematic review and meta-analysis on the global distribution, host range, and prevalence of Trypanosoma evansi. Parasit. Vectors 2019, 12, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez, M.F.; Kowalewski, M.M.; Salomón, O.D.; Schijman, A.G. Molecular characterization of trypanosomatid infections in wild howler monkeys (Alouatta caraya) in northeastern Argentina. Int. J. Parasitol. Parasites Wildl. 2016, 5, 198–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carhuallanqui, A.; Chávez, A.; Pinedo, R. Trypanosoma spp. in rats (Rattus rattus and Rattus norvegicus) from two environments: Food markets and pig farms. Rev. Inv. Vet. Perú 2017, 28, 958–968. [Google Scholar] [CrossRef] [Green Version]
- Orozco, M.M.; Argibay, H.D.; Minatel, L.; Guillemi, E.C.; Berra, Y.; Schapira, A.; Di Nucci, D.; Marcos, A.; Lois, F.; Falzone, M.; et al. A participatory approach to marsh deer (Blastocerus dichotomus) morbidity and mortality passive surveillance in Argentina: First results. Unpublished (pre-print available). [CrossRef]
- Davies, C.R.; Reithinger, R.; Campbell-Lendrum, D.; Feliciangeli, D.; Borges, R.; Rodríguez, N. The epidemiology and control of leishmaniasis in Andean countries. Cadernos de Saúde Pública 2000, 16, 925–950. [Google Scholar] [CrossRef] [Green Version]
- Roque, A.L.R.; Jansen, A.M. Wild and synanthropic reservoirs of Leishmania species in the Americas. Int. J. Parasitol. Parasites Wildl. 2014, 3, 251–262. [Google Scholar] [CrossRef] [Green Version]
- Hemmingsen, W.; Jansen, P.A.; MacKenzie, K. Crabs, leeches and trypanosomes: An unholy trinity? Mar. Pollut. Bull. 2005, 50, 336–339. [Google Scholar] [CrossRef]
- Latif, A.A.; Bakheit, M.A.; Mohamed, A.E.; Zweygarth, E. High infection rates of the tick Hyalomma anatolicum anatolicum with Trypanosoma theileri. Onderstepoort J. Vet. Res. 2004, 71, 251–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Austen, J.M.; Ryan, U.M.; Friend, J.A.; Ditcham, W.G.F.; Reid, S.A. Vector of Trypanosoma copemani identified as Ixodes sp. Parasitology 2011, 138, 866–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krige, A.-S.; Thompson, R.C.A.; Clode, P.L. ‘Hang on a tick’–Are ticks really the vectors for Australian trypanosomes? Trends Parasitol. 2019, 35, 596–606. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, H.D.; Molyneux, D.H.; Wallbanks, K.R. On Trypanosoma (Megatrypanum) talpe from Talpa europaea: Method of division and evidence of Haemogamasinae as vectors. J. Parasitol. 1987, 73, 1050–1052. [Google Scholar] [CrossRef] [PubMed]
- Bastin, P. The trypanosome journey in the tsetse fly. C. R. Biol. 2019, 342, 273–275. [Google Scholar] [CrossRef]
- Mulandane, F.C.; Snyman, L.P.; Brito, D.R.A.; Bouyer, J.; Fafetine, J.; Van Den Abbeele, J.; Oosthuizen, M.; Delespaux, V.; Neves, L. Evaluation of the relative roles of the Tabanidae and Glossinidae in the transmission of trypanosomosis in drug resistance hotspots in Mozambique. Parasit. Vectors 2020, 13, 219. [Google Scholar] [CrossRef]
- Votýpka, J.; Szabová, J.; Rádrová, J.; Zídková, L.; Svobodová, M. Trypanosoma culicavium sp. nov., an avian trypanosome transmitted by Culex mosquitoes. Int. J. Syst. Evol. Microbiol. 2012, 62, 745–754. [Google Scholar] [CrossRef] [Green Version]
- Akhoundi, M.; Kuhls, K.; Cannet, A.; Votýpka, J.; Marty, P.; Delaunay, P.; Sereno, D. A Historical overview of the classification, evolution, and dispersion of Leishmania parasites and sandflies. PLoS Negl. Trop. Dis. 2016, 10, e0004349. [Google Scholar] [CrossRef]
- Millán, J.; Travaini, A.; Zanet, S.; López-Bao, J.V.; Trisciuoglio, A.; Ferroglio, E.; Rodríguez, A. Detection of Leishmania DNA in wild foxes and associated ticks in Patagonia, Argentina, 2000 km south of its known distribution area. Parasit. Vectors 2016, 9, 241. [Google Scholar] [CrossRef] [Green Version]
- González, C.R. Reporte de Lutzomyia (Diptera: Psychodidae) en Chile. Parasitol. Día 2013, 4, 3. [Google Scholar]
- Georgieva, A.Y.; Gordon, E.R.L.; Weirauch, C. Sylvatic host associations of Triatominae and implications for Chagas disease reservoirs: A review and new host records based on archival specimens. PeerJ 2017, 5, e3826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canals, M.; González, C.; Canals, L.; Canals, A.; Cáceres, D.; Alvarado, S.; Cattan, P.; Saavedra, M.; Zulantay, I.; Apt, W. What do the numbers tell us about the temporal evolution of Chagas’ disease? Rev. Chile. Infectol. 2017, 34, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Frías-Lasserre, D.; González, C.R.; Reyes, C.; Blanco de Carvalho, D.; Oliveira, J.; Canals, M.; da Rosa, J.A. Wing polymorphism and Trypanosoma cruzi infection in wild, peridomestic and domestic collections of Mepraia spinolai (Hemiptera: Reduviidae) from Chile. J. Med. Entomol. 2017, 54, 1061–1066. [Google Scholar] [CrossRef] [PubMed]
- Jansen, A.M.; Roque, A.L.R. Domestic and wild mammalian reservoir. In American Trypanosomiasis Chagas Disease—100 Years of Research; Telleria, J., Tibayrenc, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2010. [Google Scholar]
- Añez, N.; Crisantea, G.; Soriano, P.J. Trypanosoma cruzi congenital transmission in wild bats. Acta Trop. 2009, 109, 78–80. [Google Scholar] [CrossRef]
- Daszak, P.; Cunningham, A.A.; Hyatt, A.D. Anthropogenic environmental change and the emergence of infectious diseases in wildlife. Acta Trop. 2001, 78, 103–116. [Google Scholar] [CrossRef]
- Newbold, T. Future effects of climate and land-use change on terrestrial vertebrate community diversity under different scenarios. Proc. R. Soc. B 2018, 285, 20180792. [Google Scholar] [CrossRef]
- Acosta-Jamett, G.; Cleaveland, S.; Cunningham, A.A.; Bronsvoort, B.M.D. Demography of domestic dogs in rural and urban areas of the Coquimbo region of Chile and implications for disease transmission. Prev. Vet. Med. 2010, 94, 272–281. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
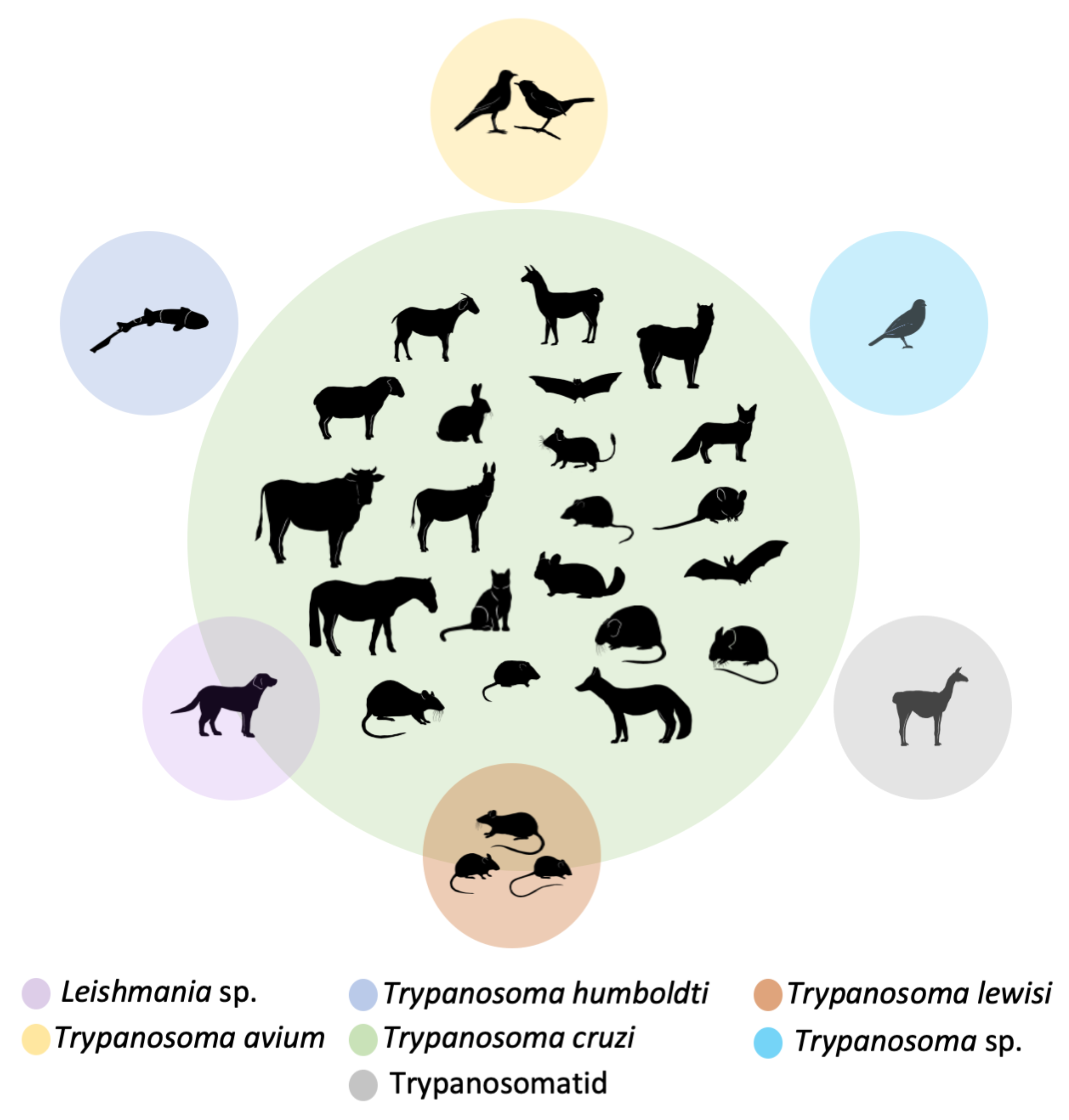
| Trypanosomatid Species | Host Class | Species |
|---|---|---|
| Leishmania sp. | Mammalia | Canis lupus familiaris |
| Trypanosoma avium | Aves | Anairetes fernandezianus, Turdus falcklandii |
| Trypanosoma cruzi | Mammalia | Abrocoma bennetti, Abrothrix longipilis, Abrothrix olivaceus, Bos taurus, Canis lupus familiaris, Capra hircus, Chinchilla lanigera, Desmodus rotundus, Equus asinus, Equus caballus, Felis catus, Histiotus montanus, Lama glama, Lycalopes culpaeus, Lycalopex griseus, Mus musculus, Octodon degus, Oligoryzomys longicaudatus, Oryctolagus cuniculus, Ovis aries, Phyllotis darwini, Rattus norvegicus, Rattus rattus, Thylamys elegans, Vicugna pacos |
| Trypanosoma humboldti | Carcharhiniformes | Schroederichthys chilensis |
| Trypanosoma lewisi | Mammalia | Mus musculus, Oligoryzomys longicaudatus, Rattus rattus |
| Trypanosoma sp. | Aves | Phrygilus fruticeti |
| Trypanosomatid | Mammalia | Lama guanicoe |
| ORDER/Species | Region(s) | Positive/ Total Tested (%) | Assay Type (Sample Type) | Reference |
|---|---|---|---|---|
| DIDELPHIMORPHIA | ||||
| Elegant fat-tailed opossum | AT/CO/VA/ME | 0/15 | OM, XD | [29] |
| (Thylamys elegans) | CO | 0/4 | XD, IHA | [30] |
| CO | 6/13 (46.2) | cPCR+SB (blood) | [17,31] | |
| CO | 8/28 (28.6) a | cPCR (blood) | [32] | |
| CO | 2/4 (50.0) a | cPCR (blood) | [33] | |
| CO/VA/ME | 6/14 (42.9) b | cPCR (blood) | [14,34] | |
| ARTIODACTYLA | ||||
| Llama | AP/TA | 0/182 | IHA | [36,49] |
| (Lama glama)c | AN | 0/37 | IHA | [37,39] |
| AP | 1/136 (0.7) | IHA | [38] | |
| Alpaca | AP | 49/439 (11.2) | IHA | [16] |
| (Vicugna pacos)c | AP | 29/2011 (1.4) | IHA | [38] |
| CARNIVORA | ||||
| Culpeo fox | AT/VA/ME | 7/533 (1.3) b | OM, XD | [15,29,40,41,42,43] |
| (Lycalopex culpaeus) | CO | 0/1 | IHA | [44] |
| South American gray fox | AP/TA/AN/AT/VA/ME/OH | 3/78 (3.8) b | XD | [15,29,40,41,42,43] |
| (Lycalopex griseus) | CO | 0/2 | IHA | [44] |
| CHIROPTERA | ||||
| Common vampire bat (Desmodus rotundus) | AT | 6/17 (35.3) | Real-time PCR (tissue) | [45] |
| Small big-eared brown bat (Histiotus montanus) | CO | 4/8 (50.0) | Real-time PCR (anal swab/feces) | [45] |
| RODENTIA | ||||
| Bennett’s chinchilla-rat | AT/VA/ME | 0/43 | OM, XD | [29] |
| (Abrocoma bennetti) | CO | 4/11 (36.4) | XD, IHA | [30] |
| ME | 0/2 | hnPCR (blood) | [48] | |
| CO | 3/7 (42.9) | cPCR (blood) | [51] | |
| CO | 4/12 (33.3)a | cPCR (blood) | [33] | |
| CO | 2/9 (22.2) | cPCR (blood) | [14,34] | |
| Long-haired grass mouse | NK | 0/2 | OM | [29] |
| (Abrothrix longipilis) | CO | 0/1 | XD, IHA | [30] |
| CO | 0/1 | IHA | [44] | |
| CO/ME | 2/21 (9.5) | cPCR (blood) | [14,34] | |
| Olive grass mouse | AT | 0/5 | OM | [29] |
| (Abrothrix olivaceus) | CO | 0/4 | IHA | [30] |
| CO | 31/44 (71.0) | cPCR+SB (blood) | [17,31] | |
| ME | 0/2 | hnPCR (blood) | [48] | |
| CO | 20/32 (62.5) | cPCR (blood) | [51] | |
| CO | 36/89 (40.5) a | cPCR (blood) | [33] | |
| CO/VA/ME | 16/41 (39.0) | cPCR (blood) | [14,34] | |
| CO | 15/45 (33.3) a | cPCR (blood) | [57] | |
| Long-tailed chinchilla | CO | 7/35 (20.0) | XD, IHA | [30] |
| (Chinchilla lanigera) | CO | 8/20 (40.0) | XD, IHA | [44] |
| Degu | AT/CO/ME | 9/412 (2.2) b | OM, XD, IHA | [15,29,40,41,42,43] |
| (Octodon degus) | CO | 5/60 (8.3) | XD, IHA | [30] |
| CO | 3/14 (21.4) | IHA | [44] | |
| CO | 28/46 (61.0) | cPCR+SB (blood) | [17,31] | |
| CO | 8/35 (22.9) a | XD-cPCR | [46,47,49] | |
| ME | 8/60 (13.3) | hnPCR (blood) | [48] | |
| CO | 38/96 (39.6) | cPCR (blood) | [32] | |
| CO | 68/140 (48.6) | cPCR (blood) | [50] | |
| CO | 69/98 (70.4) | cPCR (blood) | [51] | |
| CO | 106/262 (40.5) | cPCR (blood) | [53,54] | |
| CO | 170/460 (37.0) ab | cPCR (blood) | [33,52] | |
| CO | 40/57 (70.2) | cPCR (blood) | [55] | |
| CO | 2/4 (50.0) | qPCR (blood) | [56] | |
| CO | 107/273 (40.2) a | cPCR (blood) | [57] | |
| Degu and moon-toothed degu (O. degus and O. lunatus) | CO/VA/ME | 89/356 (25.0) | cPCR (blood) | [14,34] |
| Long-tailed rice mouse | ME | 0/11 | OM | [29] |
| (Oligoryzomys longicaudatus) | CO | 0/1 | XD | [30] |
| CO | 1/2 (50) a | cPCR (blood) | [33] | |
| CO/VA/ME | 8/45 (17.8) | cPCR (blood) | [14,34] | |
| Darwin’s leaf-eared mouse | AP/CO/VA/ME | 0/59 | OM, XD | [29] |
| (Phyllotis darwini) | CO | 1/10 (10.0) | XD, IHA | [30] |
| CO | 5/62 (8.1) | IHA | [44] | |
| CO | 31/55 (56.0) | cPCR+SB (blood) | [17,31] | |
| ME | 1/4 (25.0) | hnPCR (blood) | [48] | |
| CO | 38/117 (32.5) | cPCR (blood) | [32] | |
| CO | 63/103 (61.2) | cPCR (blood) | [51] | |
| CO | 76/210 (36.2) | cPCR (blood) | [53,54] | |
| CO | 129/379 (34.0) a | cPCR (blood) | [33] | |
| CO | 6/6 (100) | qPCR (blood) | [56] | |
| CO/VA/ME | 73/187 (39.0) | cPCR (blood) | [14,34] | |
| CO | 81/221 (36.7) a | cPCR (blood) | [57] |
| ORDER/Species | Region(s) | Positive/ Total Tested (%) | Assay Type (Sample Type) | Reference |
|---|---|---|---|---|
| ARTIODACTYLA | ||||
| Cattle | NK | 0/2 | XD | [42] |
| (Bos taurus) | ME | 0/2 | XD | [58] |
| CO | 0/1 | IHA | [16,61,65] | |
| CO | 27/202 (13.4) | IHA | [66] | |
| Goat | AT | 0/82 | OM, XD | [29] |
| (Capra hircus) | AP/TA/AN/AT/CO/VA/ME/OH | 1/233 (0.4) a | XD | [15,42,43] |
| CO | 0/2 | XD | [67] | |
| NK | 0/3 | XD | [68] | |
| CO | 32/180 (17.8) | IHA | [69] | |
| CO | 25/265 (9.4) a | IHA | [16,59,61,65] | |
| AP/TA | 0/45a | IHA | [16,36,39,65] | |
| AN | 7/98 (7.1) a | IHA | [16,37,39,65] | |
| AT | 7/100 (7.0) a | IHA | [16,60,65] | |
| VA | 0/52 a | IHA | [16,62,65] | |
| ME | 2/11 (18.2) a | IHA | [16,63,65] | |
| OH | 1/26 (3.9) a | IHA | [16,64,65] | |
| CO | 11/316 (3.5) | IHA | [70] | |
| ME | 55/841 (6.5) | IIF | [71] | |
| CO | 21/42 (50.0) | cPCR+SB (blood) | [17,31] | |
| CO | 35/100 (35.0) | IIF, XD-cPCR | [72] | |
| Sheep | AP/TA/AN/AT/CO/VA/ME/OH | 0/99 | XD | [42] |
| (Ovis aries) | CO | 2/42 (4.8) a | IHA | [16,59,61,65] |
| AP/TA | 7/161 (4.4) a | IHA | [16,36,39,65] | |
| AN | 4/147 (2.7) a | IHA | [16,37,39,65] | |
| AT | 3/16 (18.8) a | IHA | [16,60,65] | |
| VA | 1/33 (3.0) a | IHA | [16,62,65] | |
| ME | 0/1 | IHA | [16,63,65] | |
| OH | 4/25 (16.0) a | IHA | [16,64,65] | |
| CARNIVORA | ||||
| Dog | AT | 16/46 (34.8) a | OM, XD | [73,74,76] |
| (Canis lupus familiaris) | AN/AT/ME | 37/184 (20.1) | OM, XD | [75,77] |
| ME | 2/29 (6.9) | XD | [78] | |
| AT | 2/13 (15.4) | XD | [29] | |
| ME | 24/1026 (2.3) | XD | [79] | |
| AT | 23/104 (22.1) | XD | [80] | |
| AP/TA/AN/AT/CO/VA/ME/OH | 318/3591 (8.9) a | XD | [15,40,41,42,43] | |
| CO | 1/15 (6.7) | XD | [67] | |
| OH | 0/25 | XD | [81] | |
| ME | 2/98 (2.0) | XD | [58] | |
| ME | 8/86 (9.3) | XD | [82] | |
| AN/AT/VA/ME | 45/1101 (4.1) | XD | [68] | |
| CO | 44/304 (14.5) a | IHA | [16,59,61,65] | |
| AP/TA | 4/203 (2.0) a | IHA | [16,36,39,65] | |
| AN | 4/65 (6.2) a | IHA, IIF | [16,37,39,65,84] | |
| AT | 8/73 (11.0) a | IHA | [16,60,65,83] | |
| VA | 7/374 (1.9) a | IHA | [16,62,65] | |
| ME | 71/617 (11.5) a | IHA | [16,63,65] | |
| OH | 14/540 (2.6) a | IHA | [16,64,65] | |
| CO | 40/202 (19.8) | IHA | [66] | |
| CO | 20/288 (6.9) | IHA | [70] | |
| NK | 4/36 (11.1) | XD | [16] | |
| TA | 3/29 (10.4) | ELISA | [85] | |
| VA | 8/28 (28.6) | cPCR | [86] | |
| TA/CO | 38/108 (35.2) | nPCR | [87] | |
| ME | 19/111 (17.1) b | Real-time PCR (blood) | [19] | |
| Domestic cat | AT | 4/22 (18.2) a | OM, XD | [73,74,76] |
| (Felis catus) | AN/AT/ME | 6/136 (4.5) a | OM, XD | [75,77] |
| ME | 1/8 (12.5) | XD | [78] | |
| AT | 2/8 (25.0) | XD | [29] | |
| ME | 8/595 (1.4) | XD | [79] | |
| AT | 11/47 (23.4) | XD | [80] | |
| AP/TA/AN/AT/VA/ME/OH | 217/1892 (11.5) a | XD | [15,40,41,42,43] | |
| CO | 0/9 | XD | [67] | |
| OH | 0/10 | XD | [81] | |
| ME | 0/48 | XD | [58] | |
| ME | 1/27 (3.3) | XD | [82] | |
| AN/AT/VA/ME | 11/522 (2.1) | XD | [68] | |
| CO | 23/214 (10.8) a | IHA | [16,59,61,65] | |
| AP/TA | 15/140 (10.7) a | IHA | [16,36,39,65] | |
| AN | 1/32 (3.1) a | IHA | [16,37,39,65] | |
| AT | 15/165 (9.1) a | IHA | [16,60,65,83] | |
| VA | 2/197 (1.0) a | IHA | [16,62,65] | |
| ME | 33/304 (10.9) a | IHA | [16,63,65] | |
| OH | 2/93 (2.2) a | IHA | [16,64,65] | |
| NK | 4/19 (21.1) | XD | [16] | |
| LAGOMORPHA | ||||
| Rabbit | AP/AT/ME | 0/27 | OM, XD | [29] |
| (Oryctolagus cuniculus)c | AP/TA/AN/AT/CO/VA/ME | 2/209 (1.0) a | XD | [15,40,41,42,43] |
| CO | 0/2 | XD | [65] | |
| CO | 18/149 (12.1) a | IHA | [16,59,61,65] | |
| AP/TA | 1/182 (0.6) a | IHA | [16,36,39,65] | |
| AN | 7/145 (4.8) a | IHA | [16,37,39,65] | |
| AT | 0/158 a | IHA | [16,60,65,83] | |
| VA | 0/15 a | IHA | [16,62,65] | |
| ME | 3/47 (6.4) a | IHA | [16,63,65] | |
| OH | 0/8 | IHA | [16,64,65] | |
| CO | 22/58 (37.9) a | cPCR, hnPCR | [88] | |
| PERISSODACTYLA | ||||
| Ass | AT | 0/17 | XD | [29] |
| (Equus asinus) | NK | 0/74 | XD | [42] |
| CO | 0/1 | XD | [67] | |
| AT | 0/21 | IHA | [60] | |
| AP/TA/AN | 0/12 | IHA | [37,39] | |
| CO | 0/2 | IHA | [61] | |
| VA | 0/3 | IHA | [62] | |
| ME | 0/1 a | IHA | [16,63,65] | |
| CO | 18/101 (17.8) | IHA | [66] | |
| Horse | NK | 0/13 | XD | [42] |
| (Equus caballus) | AT | 0/10 a | IHA | [16,60,65] |
| CO | 16/101 (15.8) | IHA | [66] | |
| RODENTIA | ||||
| House mouse | AT/ME | 0/37 | OM, XD | [29] |
| (Mus musculus) c | CO | 5/6 (83.3) | qPCR (blood) | [56] |
| Norway rat | AT/ME | 0/4 | OM | [29] |
| (Rattus norvegicus) c | CO/VA/ME | 5/7 (71.4) b | cPCR (blood) | [14,34] |
| Black rat | AP/AT/ME | 0/11 | OM, XD | [29] |
| (Rattus rattus) c | ME | 10/44 (22.7) | hnPCR (blood) | [48] |
| CO | 46/55 (83.6) | qPCR (blood) | [56] | |
| CO/VA/ME | 14/30 (46.7) | cPCR (blood) | [14,34] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Correa, J.P.; Bacigalupo, A.; Yefi-Quinteros, E.; Rojo, G.; Solari, A.; Cattan, P.E.; Botto-Mahan, C. Trypanosomatid Infections among Vertebrates of Chile: A Systematic Review. Pathogens 2020, 9, 661. https://doi.org/10.3390/pathogens9080661
Correa JP, Bacigalupo A, Yefi-Quinteros E, Rojo G, Solari A, Cattan PE, Botto-Mahan C. Trypanosomatid Infections among Vertebrates of Chile: A Systematic Review. Pathogens. 2020; 9(8):661. https://doi.org/10.3390/pathogens9080661
Chicago/Turabian StyleCorrea, Juana P., Antonella Bacigalupo, Esteban Yefi-Quinteros, Gemma Rojo, Aldo Solari, Pedro E. Cattan, and Carezza Botto-Mahan. 2020. "Trypanosomatid Infections among Vertebrates of Chile: A Systematic Review" Pathogens 9, no. 8: 661. https://doi.org/10.3390/pathogens9080661