Development and Validation of a New TaqMan Real-Time PCR for Detection of ‘Candidatus Phytoplasma pruni’


Abstract
:1. Introduction
2. Results
2.1. In Silico Analysis of the Designed Real-Time PCRs
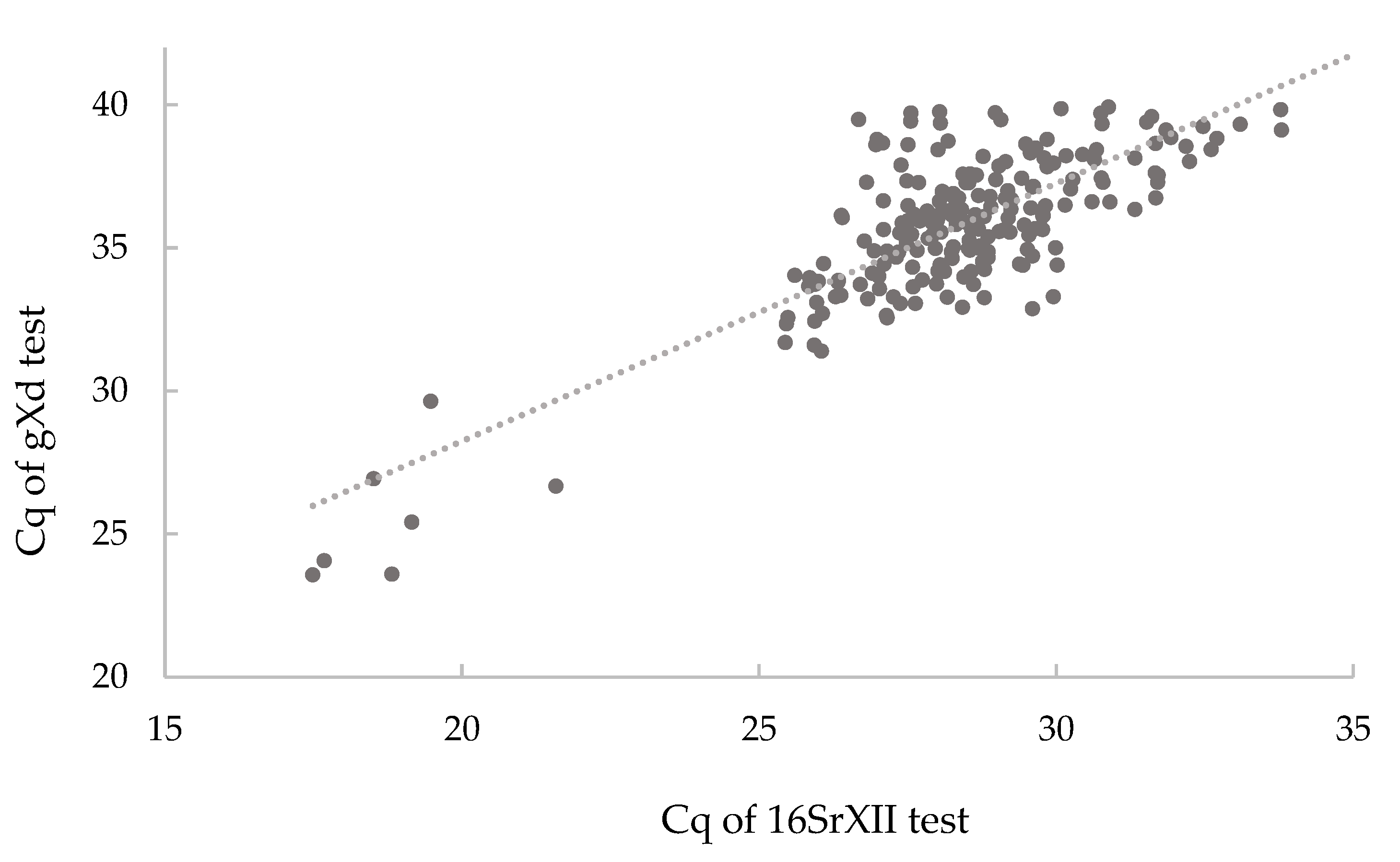
2.2. Experimental Testing of the Specificities of the sXd and gXd Tests
2.3. Analytical Sensitivity of the sXd Test
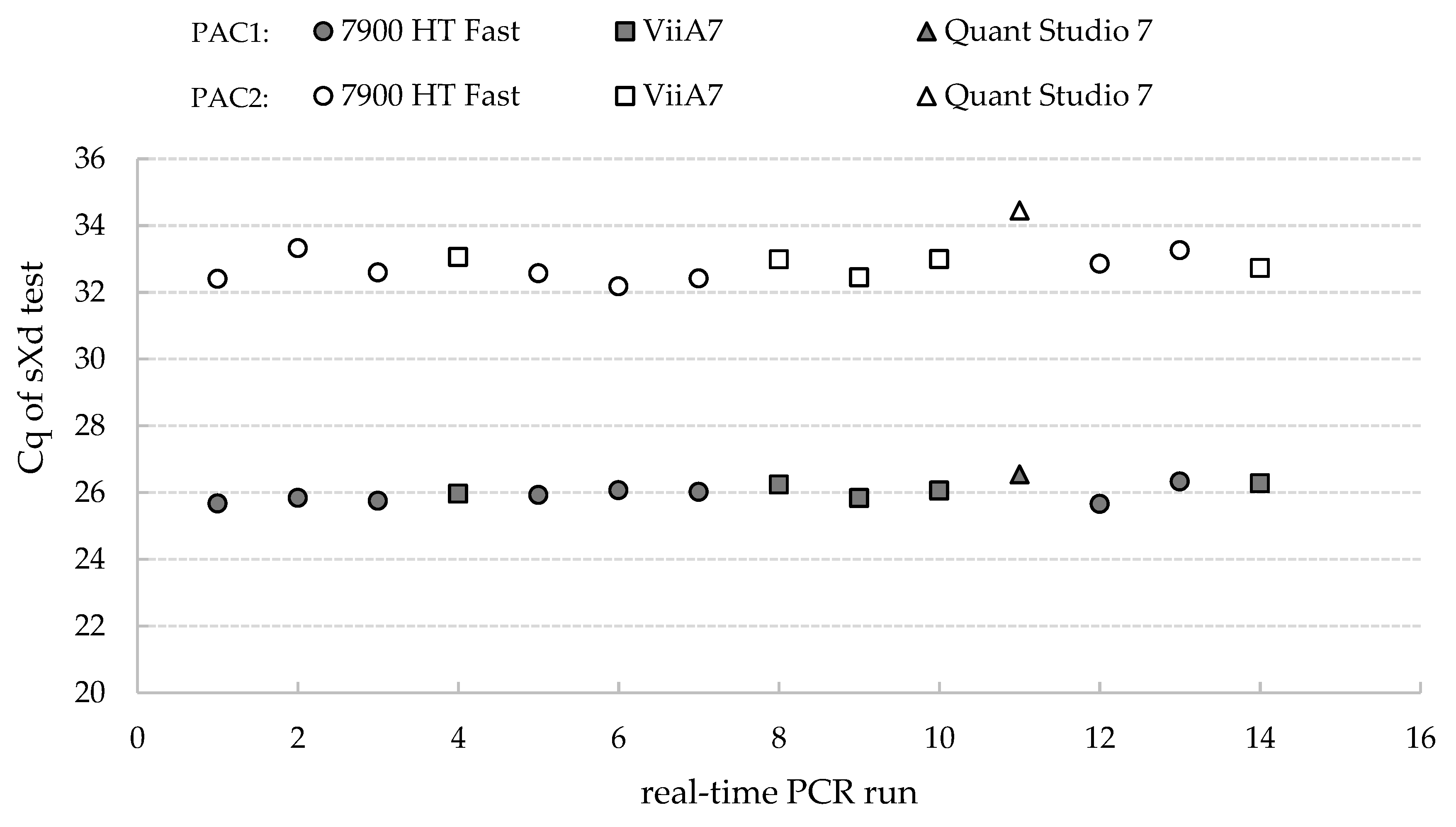
2.4. Repeatability and Reproducibility of the sXd Test
3. Discussion
4. Materials and Methods
4.1. Plant Aaterials
4.2. DNA Extraction and Detection of Phytoplasmas
4.3. Design and Set-up of the sXd and gXd Tests
4.4. Validation of the sXd and gXd Tests
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bertaccini, A.; Duduk, B.; Paltrinieri, S.; Contaldo, N. Phytoplasmas and Phytoplasma Diseases: A Severe Threat to Agriculture. Am. J. Plant Sci. 2014, 5, 1763–1788. [Google Scholar] [CrossRef] [Green Version]
- Bertaccini, A.; Lee, I.-M. Phytoplasmas: An Update. In Phytoplasmas: Plant Pathogenic Bacteria—I: Characterisation and Epidemiology of Phytoplasma—Associated Diseases; Rao, G.P., Bertaccini, A., Fiore, N., Liefting, L.W., Eds.; Springer: Singapore, 2018; pp. 1–29. [Google Scholar]
- Murray, R.G.E.; Stackebrandt, E. Taxonomic Note: Implementation of the Provisional Status Candidatus for Incompletely Described Procaryotes. Int. J. Syst. Bacteriol. 1995, 45, 186–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IRPCM. “Candidatus Phytoplasma”, a Taxon for the Wall-Less, Non-Helical Prokaryotes That Colonize Plant Phloem and Insects. Int. J. Syst. Evol. Microbiol. 2004, 54, 1243–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gundersen, D.E.; Lee, I.M.; Schaff, D.A.; Harrison, N.A.; Chang, C.J.; Davis, R.E.; Kingsbury, D.T. Genomic Diversity and Differentiation among Phytoplasma Strains in 16S rRNA Groups I (Aster Yellows and Related Phytoplasmas) and III (X-Disease and Related Phytoplasmas). Int. J. Syst. Bacteriol. 1996, 46, 64–75. [Google Scholar] [CrossRef]
- Lee, I.; Davis, R.E.; Gundersen-Rindal, D.E. Phytoplasma: Phytopathogenic Mollicutes. Annu. Rev. Microbiol. 2000, 54, 221–255. [Google Scholar] [CrossRef] [Green Version]
- Bertaccini, A. Phytoplasmas: Diversity, Taxonomy, and Epidemiology. Front. Biosci. 2007, 12, 673–689. [Google Scholar] [CrossRef] [Green Version]
- Olivier, C.Y.; Lowery, D.T.; Stobbs, L.W. Phytoplasma Diseases and their Relationships with Insect and Plant Hosts in Canadian Horticultural and Field Crops. Can. Entomol. 2009, 141, 425–462. [Google Scholar] [CrossRef]
- Davis, R.E.; Zhao, Y.; Dally, E.L.; Lee, I.M.; Jomantiene, R.; Douglas, S.M. ‘Candidatus Phytoplasma Pruni’, a Novel Taxon Associated with X-Disease of Stone Fruits, Prunus spp.: Multilocus Characterization Based on 16S rRNA, secY, and Ribosomal Protein Genes. Int. J. Syst. Evol. Microbiol. 2013, 63, 766–776. [Google Scholar] [CrossRef]
- Sinclair, W.A.; Lyon, H.H. Diseases of Trees and Shrubs, 2nd ed.; Comstock Publishing Associates: Ithaca, NY, USA, 2005. [Google Scholar]
- Scott, S.W.; Zimmerman, M. Peach Rosette, Little Peach, and Red Suture Are Diseases Induced by a Phytoplasma Closely Related to Western X-Disease. ISHS Acta Hortic. 2001, 550, 351–354. [Google Scholar] [CrossRef]
- Stoddard, E.M.; Hildebrand, E.M.; Palmiter, D.H.; Parker, H.G. X Disease. In Virus Diseases and Other Disorders with Virus-Like Symptoms of Stone Fruits in North America—Agriculture Handbook; Gilmer, R.M., Duain Moore, J., Nyland, G., Welsh, M.F., Pine, T.S., Eds.; United States Government Printing Office: Washington, DC, USA, 1951; pp. 37–42. [Google Scholar]
- Lee, I.M.; Gundersen-Rindal, D.E.; Bertaccini, A. Phytoplasma: Ecology and Genomic Diversity. Am. Phytopathol. Soc. 1998, 88, 1359–1366. [Google Scholar] [CrossRef] [Green Version]
- Van Steenwyk, R.A.; Kirkpatrick, B.C.; Purcell, A.H.; Fouche, C.F.; Grant, J.A.; Uyemoto, J.L. Evaluation of an Abatement Program for Western X-Disease in Sweet Cherry. Plant Dis. 1995, 79, 1025–1028. [Google Scholar] [CrossRef]
- Peterson, G.W. Spread and Damage of Western X-Disease of Chokecherry in Eastern Nebraska Plantings. Plant Dis. 1984, 68, 103–104. [Google Scholar] [CrossRef]
- Douglas, S.M. Detection of Mycoplasmalike Organsms in Peach and Chokecherry with X-Disease by Fluorescence Microscopy. Phytopathology 1986, 76, 784–787. [Google Scholar] [CrossRef]
- Nikolaeva, E.V.; Welliver, R.; Rosa, C.; Jones, T.; Peter, K.; Costanzo, S.; Davis, R.E. First Report of Apple (Malus Domestica) as a Host of ‘Candidatus Phytoplasma Pruni’ in the United States. Plant Dis. 2017, 101, 378. [Google Scholar] [CrossRef]
- Arocha, Y.; Zerfy, T.; Abebe, G.; Proud, J.; Hanson, J.; Wilson, M.; Jones, P.; Lucas, J. Identification of Potential Vectors and Alternative Plant Hosts for the Phytoplasma Associated with Napier Grass Stunt Disease in Ethiopia. J. Phytopathol. 2009, 157, 126–132. [Google Scholar] [CrossRef]
- De Souza, A.N.; Da Silva, F.N.; Bedendo, I.P.; Carvalho, C.M. A Phytoplasma Belonging to a 16SrIII-A Subgroup and dsRNA Virus Associated with Cassava Frogskin Disease in Brazil. Plant Dis. 2014, 98, 771–779. [Google Scholar] [CrossRef] [Green Version]
- Koinuma, H.; Miyazaki, A.; Wakaki, R.; Fujimoto, Y.; Nozomu, I.; Nijo, T.; Kitazawa, Y.; Shigaki, T.; Maejima, K.; Yamaji, Y.; et al. First Report of ‘Candidatus Phytoplasma Pruni’ Infecting Cassava in Japan. J. Gen. Plant Pathol. 2018, 84, 300–304. [Google Scholar] [CrossRef]
- Landi, F.; Prandini, A.; Paltrinieri, S.; Mori, N.; Bertaccini, A. Detection of Different Types of Phytoplasmas in Stone Fruit Orchards in Northern Italy. Bull. Insectology 2007, 60, 163–164. [Google Scholar]
- Davis, R.E.; Dally, E.L.; Zhao, Y.; Lee, I.; Wei, W.; Plant, M.; Wolf, T.K.; Beanland, L.; Smith, A.H.; LeDoux, D.G.; et al. Unraveling the Etiology of North American Grapevine Yellows (NAGY): Novel NAGY Phytoplasma Sequevars Related to ‘Candidatus Phytoplasma Pruni’. Plant Dis. 2015, 99, 1087–1097. [Google Scholar] [CrossRef] [Green Version]
- Prezelj, N.; Covington, E.; Roitsch, T.; Gruden, K.; Fragner, L.; Weckwerth, W.; Chersicola, M.; Vodopivec, M.; Dermastia, M. Metabolic Consequences of Infection of Grapevine (Vitis vinifera L.) Cv. “Modra Frankinja” with Flavescence Dorée Phytoplasma. Front. Plant Sci. 2016, 7, 711. [Google Scholar] [CrossRef]
- Jarausch, W.; Lansac, M.; Dosba, F. Seasonal Colonization Pattern of European Stone Fruit Yellows Phytoplasmas in Different Prunus Species Detected by Specific PCR. J. Phytopathol. 1999, 147, 47–54. [Google Scholar] [CrossRef]
- Dermastia, M.; Bertaccini, A.; Constable, F.; Mehle, N. Grapevine Yellows Diseases and Their Phytoplasma Agents; Springer International Publishing: Cham, Switzerland, 2017. [Google Scholar] [CrossRef]
- Mitrović, J.; Kakizawa, S.; Duduk, B.; Oshima, K.; Namba, S.; Bertaccini, A. The GroEL Gene as an Additional Marker for Finer Differentiation of ‘Candidatus Phytoplasma Asteris’-Related Strains. Ann. Appl. Biol. 2011, 159, 41–48. [Google Scholar] [CrossRef]
- Huang, D.; Walla, J.A.; Dai, W. Quantitative Phenotyping of X-Disease Resistance in Chokecherry Using Real-Time PCR. J. Microbiol. Methods 2014, 98, 1–7. [Google Scholar] [CrossRef]
- EPPO. PM 7/98 (2) Specific Requirements for Laboratories Preparing Accreditation for a Plant Pest Diagnostic Activity. EPPO Bull. 2014, 44, 117–147. [Google Scholar] [CrossRef] [Green Version]
- Valiunas, D.; Jomantiene, R.; Ivanauskas, A.; Abraitis, R.; Staniene, G.; Zhao, Y.; Davis, R.E. First Report of a New Phytoplasma Subgroup, 16SrIII-T, Associated with Decline Disease Affecting Sweet and Sour Cherry Trees in Lithuania. Plant Dis. 2009, 93, 550. [Google Scholar] [CrossRef] [PubMed]
- Fránová, J.; Špak, J.; Šimková, M. First Report of a 16SrIII-B Subgroup Phytoplasma Associated with Leaf Reddening, Virescence and Phyllody of Purple Coneflower. Eur. J. Plant Pathol. 2013, 136, 7–12. [Google Scholar] [CrossRef]
- Fernandez, F.D.; Meneguzzi, N.G.; Conci, L.R. Identification of Three Novel Subgroups within the X-Disease Group Phytoplasma Associated with Strawberry Redness Disease. Int. J. Syst. Evol. Microbiol. 2017, 67, 753–758. [Google Scholar] [CrossRef]
- Rappussi, M.C.C.; Eckstein, B.; Flôres, D.; Haas, I.C.R.; Amorim, L.; Bedendo, I.P. Cauliflower Stunt Associated with a Phytoplasma of Subgroup 16SrIII-J and the Spatial Pattern of Disease. Eur. J. Plant Pathol. 2012, 133, 829–840. [Google Scholar] [CrossRef]
- Lee, I.-M.; Gundersen-Rindal, D.E.; Davis, R.E.; Bottner, K.D.; Marcone, C.; Seemüller, E. ‘Candidatus Phytoplasma Asteris’, a Novel Phytoplasma Taxon Associated with Aster Yellows and Related Diseases. Int. J. Syst. Evol. Microbiol. 2004, 54, 1037–1048. [Google Scholar] [CrossRef] [Green Version]
- Quaglino, F.; Zhao, Y.; Casati, P.; Bulgari, D.; Bianco, P.A.; Wei, W.; Davis, R.E. ‘Candidatus Phytoplasma Solani’, a Novel Taxon Associated with Stolbur- and Bois Noir-Related Diseases of Plants. Int. J. Syst. Evol. Microbiol. 2013, 63, 2879–2894. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.M.; Gundersen, D.E.; Hammond, R.W.; Davis, R.E. Use of Mycoplasmalike Organism (MLO) Group-Specific Oligonucleotide Primers for Nested-PCR Assays to Detect Mixed-MLO Infections in a Single Host Plant. Phytopathology 1994, 84, 559–566. [Google Scholar] [CrossRef]
- Alma, A.; Davis, R.E.; Vibio, M.; Danielli, A.; Bosco, D.; Arzone, A.; Bertaccini, A. Mixed Infection of Grapevines in Northern Italy by Phytoplasmas Including 16S rRNA RFLP Subgroup 16SrI-B Strains Previously Unreported in this Host. Plant Dis. 1996, 80, 418–421; [Google Scholar] [CrossRef]
- Bertaccini, A.; Botti, S.; Martini, M.; Kaminska, M. Molecular Evidence for Mixed Phytoplasma Infection in Lily Plants. Acta Hortic. 2002, 568, 35–41. [Google Scholar] [CrossRef]
- Kamińska, M.; Śliwa, H. Mixed Infection of Dahlia Plants in Poland with Apple Proliferation and Aster Yellows Phytoplasmas. Plant Pathol. 2008, 57, 363. [Google Scholar] [CrossRef]
- Mehle, N.; Nikolić, P.; Rupar, M.; Boben, J.; Ravnikar, M.; Dermastia, M. Automated DNA Extraction for Large Numbers of Plant Samples. In Phytoplasma. Methods in Molecular Biology (Methods and Protocols); Dickinson, M., Hodgetts, J., Eds.; Humana Press: Totowa, NJ, USA, 2013; pp. 139–145. [Google Scholar]
- Christensen, N.M.; Nicolaisen, M.; Hansen, M.; Schulz, A. Distribution of Phytoplasmas in Infected Plants as Revealed by Real-Time PCR and Bioimaging. Mol. Plant-Microbe Interact. 2004, 17, 1175–1184. [Google Scholar] [CrossRef] [Green Version]
- Hren, M.; Boben, J.; Rotter, A.; Kralj, P.; Gruden, K.; Ravnikar, M. Real-Time PCR Detection Systems for Flavescence Dorée and Bois Noir Phytoplasmas in Grapevine: Comparison with Conventional PCR Detection and Application in Diagnostics. Plant Pathol. 2007, 56, 785–796. [Google Scholar] [CrossRef]
- Mehle, N.; Prezelj, N.; Hren, M.; Boben, J.; Gruden, K.; Ravnikar, M.; Dermastia, M. A Real-Time PCR Detection System for the Bois Noir and Flavescence Dorée Phytoplasmas and Quantification of the Target DNA. In Phytoplasma: Methods and Protocols, Methods in Molecular Biology; Dickinson, M., Hodgetts, J., Eds.; Humana Press: Totowa, NJ, USA, 2013; pp. 269–281. [Google Scholar]
- Nikolić, P.; Boben, J.; Hren, M.; Ravnikar, M.; Dermastia, M. Development of Diagnostic Method for Aster Yellows Phytoplasma Detection on Grapevine with Real-Time PCR. In Zbornik Predavanj in Referatov 9. Slovenskega Posvetovanja o Varstvu Rastlin; Maček, J., Ed.; Društvo za Varstvo Rastlin Slovenije: Ljubljana, Slovenia, 2009; pp. 237–241. [Google Scholar]
- Mehle, N.; Nikolić, P.; Gruden, K.; Ravnikar, M.; Dermastia, M. Real-Time PCR for Specific Detection of Three Phytoplasmas from the Apple Proliferation Group. In Phytoplasma: Methods and Protocols, Methods in Molecular Biology; Dickinson, M., Hodgetts, J., Eds.; Humana Press: Totowa, NJ, USA, 2013; pp. 269–281. [Google Scholar]
- Nikolić, P.; Mehle, N.; Gruden, K.; Ravnikar, M.; Dermastia, M. A Panel of Real-Time PCR Assays for Specific Detection of Three Phytoplasmas from the Apple Proliferation Group. Mol. Cell. Probes 2010, 24, 303–309. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT–PCR. Nucleic Acids Res. 2001, 29, 2002–2007. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Host Plant/Insect | Phytoplasma | Disease | 16Sr Group | DNA Source | No. of Samples * | Universal Phytoplasma Real-Time PCR (Cq) | sXd Test (Cq) | gXd Test (Cq) |
|---|---|---|---|---|---|---|---|---|
| Catharanthus roseus | ‘Ca. P. pruni’ | Peach Western X | III-A | DNA, Foissac | 1 | 17 | 18 | 17 |
| Unknown | Green Valley X-disease | III-A | DNA, Bertaccini | 1 | 18 | 20 | 18 | |
| Unknown | Peach yellow leaf roll | III-A | DNA, Bertaccini | 1 | 19 | 20 | 19 | |
| Unknown | Western X | III-A | DNA, Bertaccini | 1 | 18 | 20 | 19 | |
| Solidago rugosa | Golden rod yellows | III-D | DNA, Foissac | 1 | 19 | 28 | 19 | |
| Spiraea tomentosa | Spirea stunt | III-E | DNA, Foissac | 1 | 17 | 20 | 17 | |
| Asclepias syriaca | Milkweed yellows | III-F | DNA, Foissac | 1 | 18 | 26 | 18 | |
| Mix of Vitis vinifera, Hyalesthes obsoletus, Catharanthus roseus, Urtica dioica | ‘Ca. P. solani’ | Bois noir | XII-A | DNA, Brader | 1 | 16 | Negative | 24 |
| Mix of Hyalesthes obsoletus, Catharanthus roseus | 1 | 16 | Negative | 24 | ||||
| Mix of Hyalesthes obsoletus, Catharanthus roseus, Agallia ribauti | 1 | 17 | Negative | 27 | ||||
| Mix of Vitis vinifera, Catharanthus roseus, Agallia ribauti, Reptalus panzeri | 1 | 18 | Negative | 30 | ||||
| Vitis vinifera | 1 | 26 | Negative | 33 | ||||
| Zea mays | ‘Ca. P. solani’ | Maize redness | XII-A | DNA, Jović | 1 | 17 | Negative | 25 |
| 1 | 17 | Negative | 24 | |||||
| Reptalus panzeri | 1 | 24 | Negative | 31 | ||||
| Catharanthus roseus | ‘Ca. P. solani’ | XII-A | DNA, Foissac | 1 | 20 | Negative | 27 | |
| ‘Ca. P. ulmi’ | V-A | DNA, Foissac | 1 | 22 | Negative | Negative | ||
| ‘Ca. P. rubi’ | V-E | DNA, Foissac | 1 | 20 | Negative | Negative | ||
| Aster yellows group | I | Plant, tissue culture | 2 | 20–27 | Negative | Negative | ||
| Vitis vinifera | ‘Ca. P. solani’ | Bois noir | XII-A | Plant, field sampling (2018) | 10 | 25–27 | Negative | 38–40 |
| 45 | 26–32 | Negative | Negative | |||||
| Plant, field sampling (2019) | 188 | 24–32 | Negative | 32–40 | ||||
| 30 | 25–34 | Negative | Negative | |||||
| Vitis vinifera | Flavescence dorée and Bois noir | V and XII-A | Plant, field sampling (2018) | 10 | 23–30 | Negative | Negative | |
| Plant, field sampling (2019) | 4 | 25–28 | Negative | 34–39 | ||||
| 19 | 23–30 | Negative | Negative | |||||
| Vitis vinifera | Flavescence dorée | V | Plant, field sampling (2015) | 1 | 25 | Negative | Negative | |
| Plant, field sampling (2018) | 23 | 24–33 | Negative | Negative | ||||
| Plant, field sampling (2019) | 90 | 21–31 | Negative | Negative | ||||
| Vitis vinifera | Aster yellows group | I | Plant, field sampling (2019) | 1 | 28 | Negative | Negative | |
| Prunus persica | ‘Ca. P. prunorum’ | European stone fruit yellows | X-B | Plant, field sampling (2010) | 1 | 26 | Negative | Negative |
| Plant, field sampling (2018) | 2 | 24–28 | Negative | Negative | ||||
| Plant, field sampling (2019) | 1 | 24 | Negative | Negative | ||||
| Prunus armeniaca | ‘Ca. P. prunorum’ | European stone fruit yellows | X-B | Plant, field sampling (2018) | 1 | 24 | Negative | Negative |
| Plant, field sampling (2019) | 1 | 27 | Negative | Negative | ||||
| Malus domestica | ‘Ca. P. mali’ | Apple proliferation | X-A | Plant, field sampling (2010) | 2 | 28–32 | Negative | Negative |
| Pyrus communis | ‘Ca. P. pyri’ | Pear decline | X-C | Plant, field sampling (2018) | 1 | 24 | Negative | Negative |
| Prunus persica | Negative for phytoplasmas | Plant, field sampling (2018) | 1 | Negative | Negative | Negative | ||
| Prunus subg. Prunus | Plant, field sampling (2016) | 1 | Negative | Negative | Negative | |||
| Malus domestica | Plant, field sampling (2018) | 1 | Negative | Negative | Negative | |||
| Vitis vinifera | Plant, field sampling (2018) | 1 | Negative | Negative | Negative |
| 16SrIII-A Isolate | Dilution Factors | Linear Regression | |||
|---|---|---|---|---|---|
| Range of Detection | Dynamic Range | k | R2 | E | |
| Green Valley X-disease | 10−1–10−7 | 10−1–10−6 | −3.389 | 0.998 | 0.973 |
| Peach yellow leaf roll | 10−1–10−7 | 10−1–10−5 | −3.386 | 0.999 | 0.974 |
| Western X | 10−1–10−7 | 10−1–10−6 | −3.358 | 0.998 | 0.985 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kogej, Z.; Dermastia, M.; Mehle, N. Development and Validation of a New TaqMan Real-Time PCR for Detection of ‘Candidatus Phytoplasma pruni’. Pathogens 2020, 9, 642. https://doi.org/10.3390/pathogens9080642
Kogej Z, Dermastia M, Mehle N. Development and Validation of a New TaqMan Real-Time PCR for Detection of ‘Candidatus Phytoplasma pruni’. Pathogens. 2020; 9(8):642. https://doi.org/10.3390/pathogens9080642
Chicago/Turabian StyleKogej, Zala, Marina Dermastia, and Nataša Mehle. 2020. "Development and Validation of a New TaqMan Real-Time PCR for Detection of ‘Candidatus Phytoplasma pruni’" Pathogens 9, no. 8: 642. https://doi.org/10.3390/pathogens9080642