Polycystic Kidney Disease Ryanodine Receptor Domain (PKDRR) Proteins in Oomycetes



{kind=link}
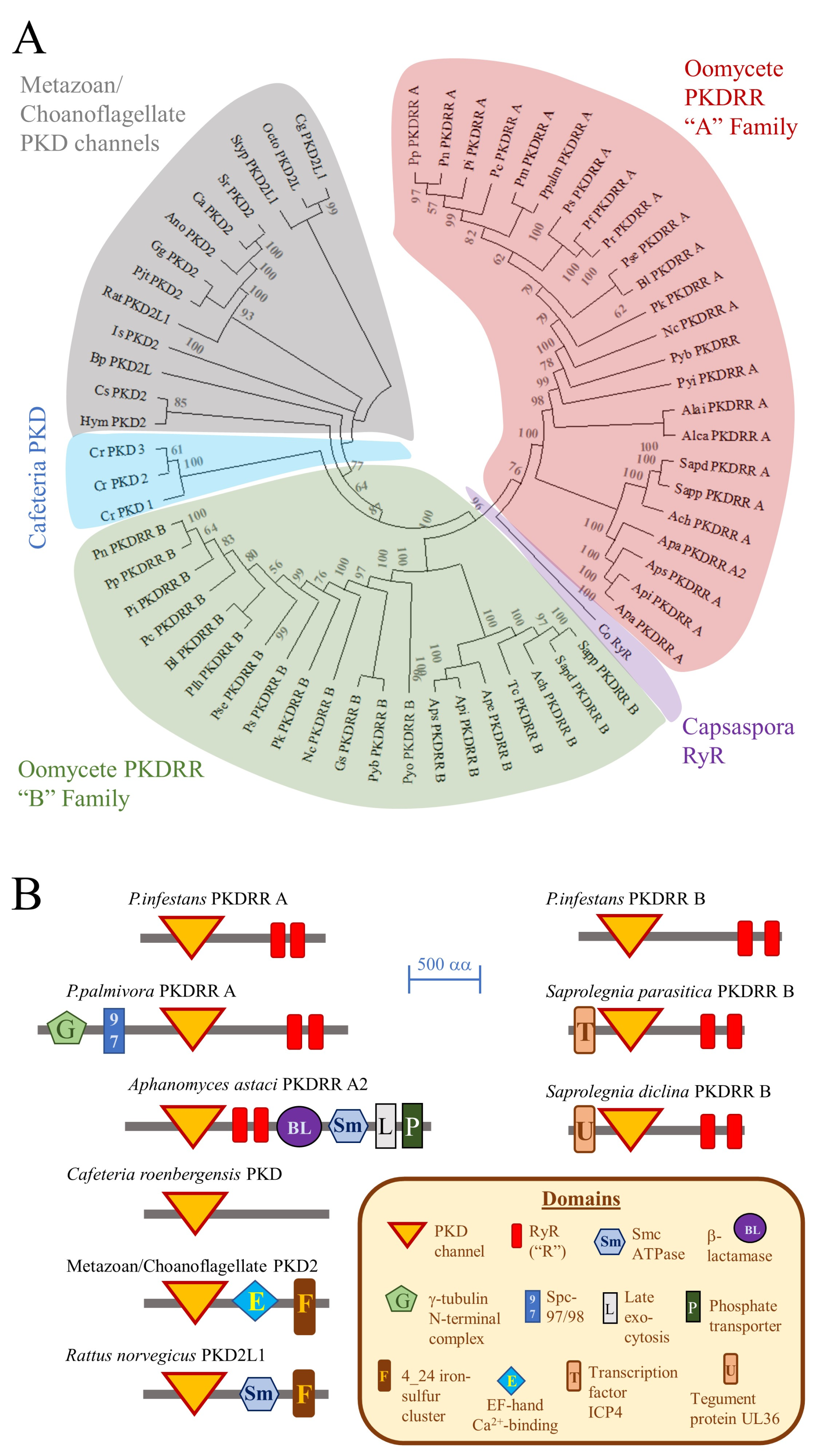{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Oomycetes Genomes Encode Putative ITPR Homologues
2.2. Characterisation of the PKDRR Family of Proteins
2.3. Analysis of the Predicted Channel Domains of Oomycete ITPR and PKDRR Channels
2.4. Biochemical Characterisation of the PKDRR B Protein from Phytophthora infestans
2.5. Generation of a Heterologous Expression System for Analysis of PKDRR B Channel Function
3. Conclusions
4. Materials and Methods
4.1. Materials
4.2. Evolutionary History of Phytophthora Infestans ITPR and PKDRR Proteins
4.3. Culture of Phytophthora Infestans
4.4. Heterologous Expression of the P. infestans PKDRR B Protein in Human Embryonic Kidney Cell-line
4.5. Protein Characterisation
4.6. Indirect Immunofluorescent Microscopy
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cavalier-Smith, T.; Scoble, J.M. Phylogeny of Heterokonta: Incisomonas marina, a uniciliate gliding opalozoan related to Solenicola (Nanomonadea), and evidence that Actinophryida evolved from raphidophytes. Eur. J. Protistol. 2013, 49, 328–353. [Google Scholar] [CrossRef] [PubMed]
- Judelson, H.S.; Blanco, F.A. The spores of Phytophthora: Weapons of the plant destroyer. Nat. Rev. Microbiol. 2005, 3, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.; Cai, M.; Dong, X.; Liu, L.; Lin, D.; Zhang, C.; Pang, Z.; Liu, X. Resistance assessment for oxathiapiprolin in phytophthora capsici and the detection of a point mutation (G769W) in PcORP1 that confers resistance. Front. Microbiol. 2016, 7, 615. [Google Scholar] [CrossRef]
- Plattner, H.; Verkhratsky, A. Inseparable tandem: Evolution chooses ATP and Ca2+ to control life, death and cellular signalling. Philos. Trans. R Soc. Lond. B Biol. Sci. 2016, 371, 20150419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berridge, M.J.; Lipp, P.; Bootman, M.D. The versatility and universality of calcium signalling. Nat. Rev. Mol. Cell Biol. 2000, 1, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Samanta, K.; Parekh, A.B. Spatial Ca(2+) profiling: Decrypting the universal cytosolic Ca(2+) oscillation. J. Physiol. 2017, 595, 3053–3062. [Google Scholar] [CrossRef]
- Mikoshiba, K. Role of IP3 receptor signaling in cell functions and diseases. Adv. Biol. Regul. 2015, 57, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Taylor, C.W.; Tovey, S.C. IP(3) receptors: Toward understanding their activation. Cold Spring Harb. Perspect. Biol. 2010, 2, a004010. [Google Scholar] [CrossRef]
- Cai, X.; Clapham, D.E. Ancestral Ca2+ signaling machinery in early animal and fungal evolution. Mol. Biol. Evol. 2012, 29, 91–100. [Google Scholar] [CrossRef]
- Mackrill, J.J. Ryanodine receptor calcium release channels: An evolutionary perspective. Adv. Exp. Med. Biol. 2012, 740, 159–182. [Google Scholar]
- Endo, M. Calcium-induced release of calcium from the sarcoplasmic reticulum. Adv. Exp. Med. Biol. 2007, 592, 275–285. [Google Scholar]
- Zheng, L.; Mackrill, J.J. Calcium signaling in oomycetes: An evolutionary perspective. Front. Physiol. 2016, 7, 123. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, S.P.; Deacon, J.W. Changes in motility of Pythium zoospores induced by calcium and calcium-modulating drugs. Mycol. Res. 1993, 97, 877–883. [Google Scholar] [CrossRef]
- Hua, C.; Wang, Y.; Zheng, X.; Dou, D.; Zhang, Z.; Govers, F.; Wang, Y. A Phytophthora sojae G-protein alpha subunit is involved in chemotaxis to soybean isoflavones. Eukaryot. Cell 2008, 7, 2133–2140. [Google Scholar] [CrossRef]
- Liu, P.; Gong, J.; Ding, X.; Jiang, Y.; Chen, G.; Li, B.; Weng, Q.; Chen, Q. The L-type Ca(2+) Channel Blocker Nifedipine Inhibits Mycelial Growth, Sporulation, and Virulence of Phytophthora capsici. Front. Microbiol. 2016, 7, 1236. [Google Scholar] [PubMed] [Green Version]
- Jackson, S.L.; Hardham, A.R. A transient rise in cytoplasmic free calcium is required to induce cytokinesis in zoosporangia of Phytophthora cinnamomi. Eur. J. Cell Biol. 1996, 69, 180–188. [Google Scholar]
- Hwu, F.Y.; Lai, M.W.; Liou, R.F. PpMID1 Plays a Role in the Asexual Development and Virulence of Phytophthora parasitica. Front. Microbiol. 2017, 8, 610. [Google Scholar] [CrossRef]
- Tani, S.; Yatzkan, E.; Judelson, H.S. Multiple pathways regulate the induction of genes during zoosporogenesis in Phytophthora infestans. Mol. Plant Microbe Interact. 2004, 17, 330–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ah-Fong, A.M.; Kim, K.S.; Judelson, H.S. RNA-seq of life stages of the oomycete Phytophthora infestans reveals dynamic changes in metabolic, signal transduction, and pathogenesis genes and a major role for calcium signaling in development. BMC Genom. 2017, 18, 198. [Google Scholar] [CrossRef]
- Chapman, J.A.; Vujicic, R. The fine structure of sporangia of Phytophthora erythroseptica Pethyb. J. Gen. Microbiol. 1965, 41, 275–282. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Matari, N.H.; Blair, J.E. A multilocus timescale for oomycete evolution estimated under three distinct molecular clock models. BMC Evol. Biol. 2014, 14, 101. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, K.; Anamika, K.; Srinivasan, N. Evolution of domain combinations in protein kinases and its implications for functional diversity. Prog. Biophys. Mol. Biol. 2010, 102, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; et al. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2017, 45, D200–D203. [Google Scholar] [CrossRef]
- Ponting, C.P. Novel repeats in ryanodine and IP3 receptors and protein O-mannosyltransferases. Trends Biochem. Sci. 2000, 25, 48–50. [Google Scholar] [CrossRef]
- Birnbaumer, L.; Yildirim, E.; Abramowitz, J. A comparison of the genes coding for canonical TRP channels and their M, V and P relatives. Cell Calcium 2003, 33, 419–432. [Google Scholar] [CrossRef]
- Hackl, T.; Martin, R.; Barenhoff, K.; Duponchel, S.; Heider, D.; Fischer, M.G. Four high-quality draft genome assemblies of the marine heterotrophic nanoflagellate Cafeteria roenbergensis. Sci. Data 2020, 7, 1–9. [Google Scholar] [CrossRef]
- Plattner, H.; Verkhratsky, A. Ca2+ signalling early in evolution—All but primitive. J. Cell Sci. 2013, 126, 2141–2150. [Google Scholar] [CrossRef] [Green Version]
- Schug, Z.T.; da Fonseca, P.C.; Bhanumathy, C.D.; Wagner, L., II; Zhang, X.; Bailey, B.; Morris, E.P.; Yule, D.I.; Joseph, S.K. Molecular characterization of the inositol 1,4,5-trisphosphate receptor pore-forming segment. J. Biol. Chem. 2008, 283, 2939–2948. [Google Scholar] [CrossRef]
- Su, Q.; Hu, F.; Liu, Y.; Ge, X.; Mei, C.; Yu, S.; Shen, A.; Zhou, Q.; Yan, C.; Lei, J.; et al. Cryo-EM structure of the polycystic kidney disease-like channel PKD2L1. Nat. Commun. 2018, 9, 1192. [Google Scholar] [CrossRef]
- Delmas, P. Polycystins: Polymodal receptor/ion-channel cellular sensors. Pflugers Arch. 2005, 451, 264–276. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Hackmann, K.; Gao, J.; He, X.; Piontek, K.; García-González, M.A.; Menezes, L.F.; Xu, H.; Germino, G.G.; Zuo, J.; et al. Essential role of cleavage of Polycystin-1 at G protein-coupled receptor proteolytic site for kidney tubular structure. Proc. Natl. Acad. Sci. USA 2007, 104, 18688–18693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letunic, I.; Bork, P. 20 years of the SMART protein domain annotation resource. Nucleic Acids Res. 2017, 46, D493–D496. [Google Scholar] [CrossRef] [PubMed]
- Zorzato, F.; Volpe, P.; Damiani, E.; Quaglino, D., Jr.; Margreth, A. Terminal cisternae of denervated rabbit skeletal muscle: Alterations of functional properties of Ca2+ release channels. Am. J. Physiol. 1989, 257, C504–C511. [Google Scholar] [CrossRef]
- Wang, D. IntronDB: A database for eukaryotic intron features. Bioinformatics 2019, 35, 4400–4401. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, T.J.; Butler, M.I.; Poulter, R.T. Multiple, non-allelic, intein-coding sequences in eukaryotic RNA polymerase genes. BMC Biol. 2006, 4, 38. [Google Scholar] [CrossRef] [Green Version]
- Mills, K.V.; Johnson, M.A.; Perler, F.B. Protein splicing: How inteins escape from precursor proteins. J. Biol. Chem. 2014, 289, 14498–14505. [Google Scholar] [CrossRef] [Green Version]
- Perler, F.B. InBase: The Intein Database. Nucleic Acids Res. 2002, 30, 383–384. [Google Scholar] [CrossRef]
- Yang, Y.; Keeler, C.; Kuo, I.Y.; Lolis, E.J.; Ehrlich, B.E.; Hodsdon, M.E. Oligomerization of the polycystin-2 C-terminal tail and effects on its Ca2+-binding properties. J. Biol. Chem. 2015, 290, 10544–10554. [Google Scholar] [CrossRef] [Green Version]
- Leuenroth, S.J.; Okuhara, D.; Shotwell, J.D.; Markowitz, G.S.; Yu, Z.; Somlo, S.; Crews, C.M. Triptolide is a traditional Chinese medicine-derived inhibitor of polycystic kidney disease. Proc. Natl. Acad. Sci. USA 2007, 104, 4389–4394. [Google Scholar] [CrossRef]
- Hu, L.; Wang, D.; Liu, L.; Chen, J.; Xue, Y.; Shi, Z. Ca(2+) efflux is involved in cinnamaldehyde-induced growth inhibition of Phytophthora capsici. PLoS ONE 2013, 8, e76264. [Google Scholar]
- Forsberg, E.J.; Feuerstein, G.; Shohami, E.; Pollard, H.B. Adenosine triphosphate stimulates inositol phospholipid metabolism and prostacyclin formation in adrenal medullary endothelial cells by means of P2-purinergic receptors. Proc. Natl. Acad. Sci. USA 1987, 84, 5630–5634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, C.L.; Abdallah, S.; Wong, Y.Y.; Le, P.; Harracksingh, A.N.; Artinian, L.; Tamvacakis, A.N.; Rehder, V.; Reese, T.S.; Senatore, A. Evolutionary insights into T-type Ca(2+) channel structure, function, and ion selectivity from the Trichoplax adhaerens homologue. J. Gen. Physiol. 2017, 149, 483–510. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Tyler, B.M. Efficient disruption and replacement of an effector gene in the oomycete Phytophthora sojae using CRISPR/Cas9. Mol. Plant Pathol. 2016, 17, 127–139. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Caten, C.E.; Jinks, J.L. Spontaneous variability of single isolates of Phytophthora infestans. I. Cultural variation. Can. J. Bot. 1968, 46, 329–348. [Google Scholar] [CrossRef]
- Schüepp, H.; Herzog, J. Reliable method to produce single-zoospore cultures of Plasmopara viticola/Zuverlässige Methode zur Herstellung von Monozoosporkulturen von Plasmopara viticola. Z. Pflanzenkrankh. Pflanzenschutz J. Plant Dis. Prot. 1986, 93, 30–32. [Google Scholar]
- Lordan, S.; O’Brien, N.M.; Mackrill, J.J. The role of calcium in apoptosis induced by 7beta-hydroxycholesterol and cholesterol-5beta,6beta-epoxide. J. Biochem. Mol. Toxicol. 2009, 23, 324–332. [Google Scholar] [CrossRef]
- Grynkiewicz, G.; Poenie, M.; Tsien, R.Y. A new generation of Ca2+ indicators with greatly improved fluorescence properties. J. Biol. Chem. 1985, 260, 3440–3450. [Google Scholar] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Hammoud, Y.; Rice, T.; Mackrill, J.J. Oxysterols modulate calcium signalling in the A7r5 aortic smooth muscle cell-line. Biochimie 2013, 95, 568–577. [Google Scholar] [CrossRef] [PubMed]
- Ah-Fong, A.M.; Judelson, H.S. New role for Cdc14 phosphatase: Localization to basal bodies in the oomycete phytophthora and its evolutionary coinheritance with eukaryotic flagella. PLoS ONE 2011, 6, e16725. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, L.; Prestwich, B.D.; Harrison, P.T.; Mackrill, J.J. Polycystic Kidney Disease Ryanodine Receptor Domain (PKDRR) Proteins in Oomycetes. Pathogens 2020, 9, 577. https://doi.org/10.3390/pathogens9070577
Zheng L, Prestwich BD, Harrison PT, Mackrill JJ. Polycystic Kidney Disease Ryanodine Receptor Domain (PKDRR) Proteins in Oomycetes. Pathogens. 2020; 9(7):577. https://doi.org/10.3390/pathogens9070577
Chicago/Turabian StyleZheng, Limian, Barbara Doyle Prestwich, Patrick T. Harrison, and John J. Mackrill. 2020. "Polycystic Kidney Disease Ryanodine Receptor Domain (PKDRR) Proteins in Oomycetes" Pathogens 9, no. 7: 577. https://doi.org/10.3390/pathogens9070577