Changes in miR-122 and Cholesterol Expression in Chronic Hepatitis C Patients after PegIFN-Alpha/Ribavirin Treatment


Abstract
:1. Introduction
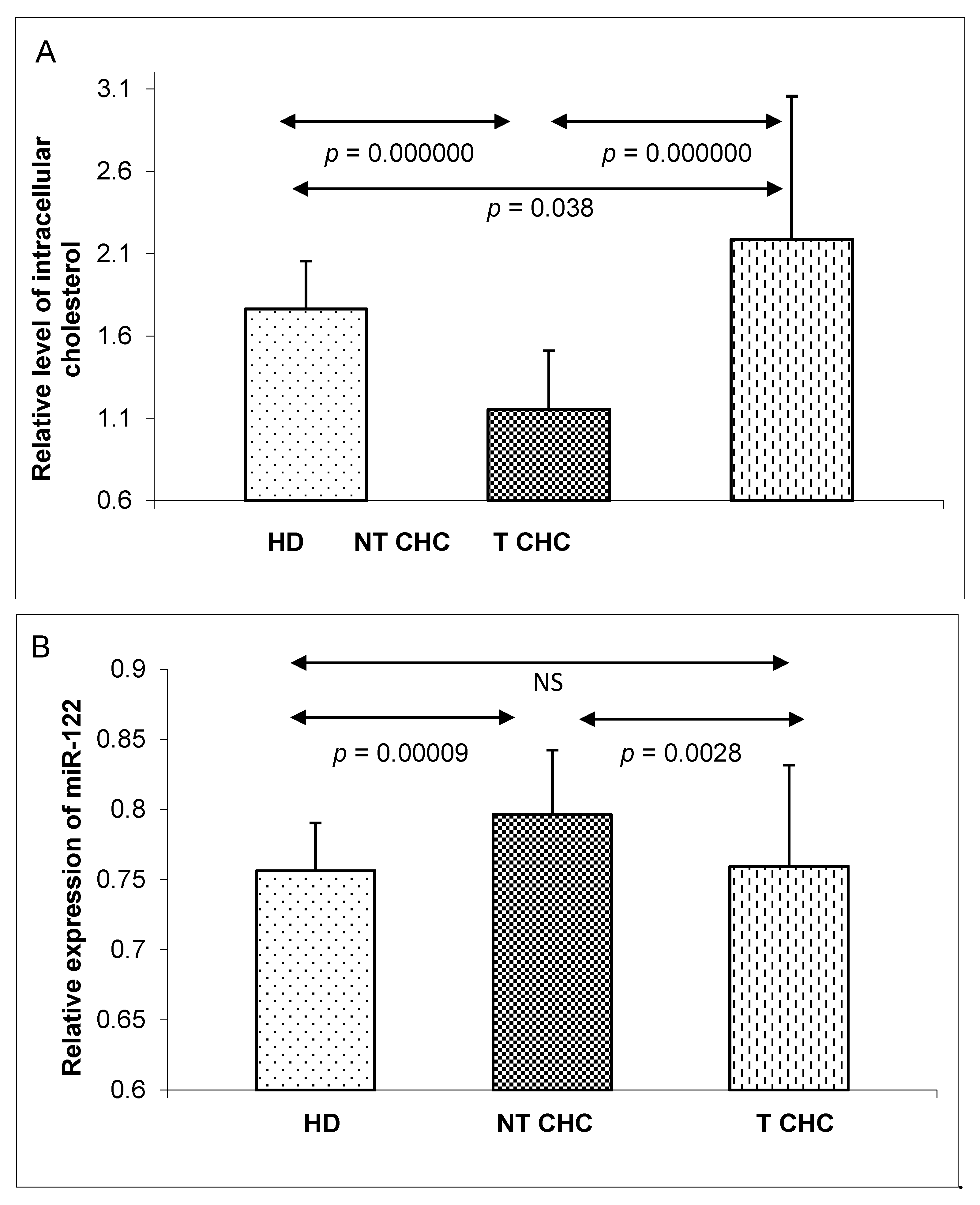
2. Results
3. Discussion
4. Materials and Methods
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gower, E.; Estes, C.; Blach, S.; Razavi-Shearer, K.; Razavi, H. Global epidemiology and genotype distribution of the hepatitis C virus infection. J. Hepatol. 2014, 61, S45–S57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popescu, C.I.; Riva, L.; Vlaicu, O.; Farhat, R.; Rouillé, Y.; Dubuisson, J. Hepatitis C virus life cycle and lipid metabolism. Biol. (Basel) 2014, 3, 892–921. [Google Scholar] [CrossRef] [PubMed]
- André, P.; Komurian-Pradel, F.; Deforges, S.; Perret, M.; Berland, J.L.; Sodoyer, M.; Pol, S.; Bréchot, C.; Paranhos-Baccalà, G.; Lotteau, V. Characterization of low- and very-low-density hepatitis C virus RNA-containing particles. J. Virol. 2002, 76, 6919–6928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albecka, A.; Belouzard, S.; Op de Beeck, A.; Descamps, V.; Goueslain, L.; Bertrand-Michel, J.; Tercé, F.; Duverlie, G.; Rouillé, Y.; Dubuisson, J. Role of low-density lipoprotein receptor in the hepatitis C virus life cycle. Hepatology 2012, 55, 998–1007. [Google Scholar] [CrossRef]
- Scarselli, E.; Ansuini, H.; Cerino, R.; Roccasecca, R.M.; Acali, S.; Filocamo, G.; Traboni, C.; Nicosia, A.; Cortese, R.; Vitelli, A. The human scavenger receptor class B type I is a novel candidate receptor for the hepatitis C virus. EMBO J. 2002, 21, 5017–5025. [Google Scholar] [CrossRef] [Green Version]
- Kapadia, S.B.; Chisari, F.V. Hepatitis C virus RNA replication is regulated by host geranylgeranylation and fatty acids. Proc. Natl. Acad. Sci. USA 2005, 102, 2561–2566. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, S.; Krajewski, M.; Scherer, C.; Scholz, V.; Mordhorst, V.; Truschow, P.; Schöbel, A.; Reimer, R.; Schwudke, D.; Herker, E. Complex lipid metabolic remodeling is required for efficient hepatitis C virus replication. Biochim. Et Biophys. Acta (Bba) Mol. Cell Biol. Lipids 2018, 1863, 1041–1056. [Google Scholar] [CrossRef]
- Carreño, V.; Pardo, M.; López-Alcorocho, J.M.; Rodríguez-Iñigo, E.; Bartolomé, J.; Castillo, I. Detection of hepatitis C virus (HCV) RNA in the liver of healthy, anti-HCV antibody-positive, serum HCV RNA-negative patients with normal alanine aminotransferase levels. J. Infect. Dis. 2006, 194, 53–60. [Google Scholar] [CrossRef]
- Baré, P.; Massud, I.; Parodi, C.; Belmonte, L.; García, G.; Nebel, M.C.; Corti, M.; Pinto, M.T.; Bianco, R.P.; Bracco, M.M.; et al. Continuous release of hepatitis C virus (HCV) by peripheral blood mononuclear cells and B-lymphoblastoid cell-line cultures derived from HCV-infected patients. J. Gen. Virol. 2005, 86, 1717–1727. [Google Scholar] [CrossRef]
- Pham, T.N.; MacParland, S.A.; Mulrooney, P.M.; Cooksley, H.; Naoumov, N.V.; Michalak, T.I. Hepatitis C virus persistence after spontaneous or treatment-induced resolution of hepatitis C. J. Virol. 2004, 78, 5867–5874. [Google Scholar] [CrossRef] [Green Version]
- Sidorkiewicz, M.; Grek, M.; Jozwiak, B.; Piekarska, A. Decreased level of intracellular cholesterol in peripheral blood mononuclear cells is associated with chronic hepatitis C virus infection. Virus Res. 2013, 178, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, I.M.; Cheng, G.; Wieland, S.; Volinia, S.; Croce, C.M.; Chisari, F.V.; David, M. Interferon modulation of cellular microRNAs as an antiviral mechanism. Nature 2007, 449, 919–922. [Google Scholar] [CrossRef]
- Jopling, C.L.; Yi, M.; Lancaster, A.M.; Lemon, S.M.; Sarnow, P. Modulation of hepatitis C virus RNA abundance by a liver-specific MicroRNA. Science 2005, 309, 1577–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henke, J.I.; Goergen, D.; Zheng, J.; Song, Y.; Schüttler, C.G.; Fehr, C.; Jünemann, C.; Niepmann, M. microRNA-122 stimulates translation of hepatitis C virus RNA. EMBO J. 2008, 27, 3300–3310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esau, C.; Davis, S.; Murray, S.F.; Yu, X.X.; Pandey, S.K.; Pear, M.; Watts, L.; Booten, S.L.; Graham, M.; McKay, R.; et al. miR-122 regulation of lipid metabolism revealed by in vivo antisense targeting. Cell Metab. 2006, 3, 87–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavie, M.; Dubuisson, J. Interplay between hepatitis C virus and lipid metabolism during virus entry and assembly. Biochimie 2017, 141, 62–69. [Google Scholar] [CrossRef]
- Lambert, J.E.; Bain, V.G.; Ryan, E.A.; Thomson, A.B.; Clandinin, M.T. Elevated lipogenesis and diminished cholesterol synthesis in patients with hepatitis C viral infection compared to healthy humans. Hepatology 2013, 57, 1697–1704. [Google Scholar] [CrossRef]
- Honda, A.; Matsuzaki, Y. Cholesterol and chronic hepatitis C virus infection. Hepatol. Res. 2011, 41, 697–710. [Google Scholar] [CrossRef]
- Tada, S.; Saito, H.; Ebinuma, H.; Ojiro, K.; Yamagishi, Y.; Kumagai, N.; Inagaki, Y.; Masuda, T.; Nishida, J.; Takahashi, M.; et al. Treatment of hepatitis C virus with peg-interferon and ribavirin combination therapy significantly affects lipid metabolism. Hepatol. Res. 2009, 39, 195–199. [Google Scholar] [CrossRef]
- Chida, T.; Kawata, K.; Ohta, K.; Matsunaga, E.; Ito, J.; Shimoyama, S.; Yamazaki, S.; Noritake, H.; Suzuki, T.; Suda, T.; et al. Rapid Changes in Serum Lipid Profiles during Combination Therapy with Daclatasvir and Asunaprevir in Patients Infected with Hepatitis C Virus Genotype 1b. Gut Liver 2018, 12, 201–207. [Google Scholar] [CrossRef] [Green Version]
- Ramcharran, D.; Wahed, A.S.; Conjeevaram, H.S.; Evans, R.W.; Wang, T.; Belle, S.H.; Yee, L.J.; Group, V.-C.S. Associations between serum lipids and hepatitis C antiviral treatment efficacy. Hepatology 2010, 52, 854–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackard, J.T.; Kemmer, N.; Sherman, K.E. Extrahepatic replication of HCV: Insights into clinical manifestations and biological consequences. Hepatology 2006, 44, 15–22. [Google Scholar] [CrossRef] [PubMed]
- MacParland, S.A.; Pham, T.N.; Guy, C.S.; Michalak, T.I. Hepatitis C virus persisting after clinically apparent sustained virological response to antiviral therapy retains infectivity in vitro. Hepatology 2009, 49, 1431–1441. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Liu, B.; Wu, J.Q.; Soriano, V.; Vispo, E.; Carroll, A.P.; Goldie, B.J.; Cairns, M.J.; Saksena, N.K. Genome-wide mRNA and miRNA analysis of peripheral blood mononuclear cells (PBMC) reveals different miRNAs regulating HIV/HCV co-infection. Virology 2014, 450, 336–349. [Google Scholar] [CrossRef]
- Boldanova, T.; Suslov, A.; Heim, M.H.; Necsulea, A. Transcriptional response to hepatitis C virus infection and interferon-alpha treatment in the human liver. EMBO Mol. Med. 2017, 9, 816–834. [Google Scholar] [CrossRef] [Green Version]
- Köberle, V.; Waidmann, O.; Kronenberger, B.; Andrei, A.; Susser, S.; Füller, C.; Perner, D.; Zeuzem, S.; Sarrazin, C.; Piiper, A. Serum microRNA-122 kinetics in patients with chronic hepatitis C virus infection during antiviral therapy. J. Viral. Hepat. 2013, 20, 530–535. [Google Scholar] [CrossRef]
- Van der Ree, M.H.; Stelma, F.; Willemse, S.B.; Brown, A.; Swadling, L.; Van der Valk, M.; Sinnige, M.J.; Van Nuenen, A.C.; De Vree, J.M.L.; Klenerman, P.; et al. Immune responses in DAA treated chronic hepatitis C patients with and without prior RG-101 dosing. Antivir. Res. 2017, 146, 139–145. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Analyzed Parameters | CHC Before | CHC After | p Value + |
|---|---|---|---|
| Serum HCV RNA positive (%) | 21 (100) | 7 (29.2) | ND |
| Serum HCV RNA load (× 105 IU/mL) | 3.245 ± 2.99 | 0.3333 ± 0.48 | 0.0008 |
| ALT (IU/L) | 67.8 ± 50.5 | 35.32 ± 18.5 | 0.003 |
| G/A HCV RNA strand (PBMCs) | 21/13 | 11/4 | ND |
| TC (mmol/L) | 2.669 ± 0.72 | 2.8476 ± 0.57 | 0.1098 |
| LDL (mmol/L) | 1.4629 ± 0.5 | 1.5938 ± 0.5 | 0.1023 |
| HDL (mmol/L) | 0.8062 ± 0.27 | 0.9162 ± 0.23 | 0.0137 |
| IC (PBMCs) | 1.8986 ± 0.56 | 2.4047 ± 0.82 | 0.0228 |
| miR-122 (PBMCs) | 0.8027 ± 0.06 | 0.7070 ± 0.17 | 0.0005 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sidorkiewicz, M.; Grek-Kowalinska, M.; Piekarska, A. Changes in miR-122 and Cholesterol Expression in Chronic Hepatitis C Patients after PegIFN-Alpha/Ribavirin Treatment. Pathogens 2020, 9, 514. https://doi.org/10.3390/pathogens9060514
Sidorkiewicz M, Grek-Kowalinska M, Piekarska A. Changes in miR-122 and Cholesterol Expression in Chronic Hepatitis C Patients after PegIFN-Alpha/Ribavirin Treatment. Pathogens. 2020; 9(6):514. https://doi.org/10.3390/pathogens9060514
Chicago/Turabian StyleSidorkiewicz, Malgorzata, Martyna Grek-Kowalinska, and Anna Piekarska. 2020. "Changes in miR-122 and Cholesterol Expression in Chronic Hepatitis C Patients after PegIFN-Alpha/Ribavirin Treatment" Pathogens 9, no. 6: 514. https://doi.org/10.3390/pathogens9060514