Mitigation of Expression of Virulence Genes in Legionella pneumophila Internalized in the Free-Living Amoeba Willaertia magna C2c Maky


Abstract
:1. Introduction
2. Results
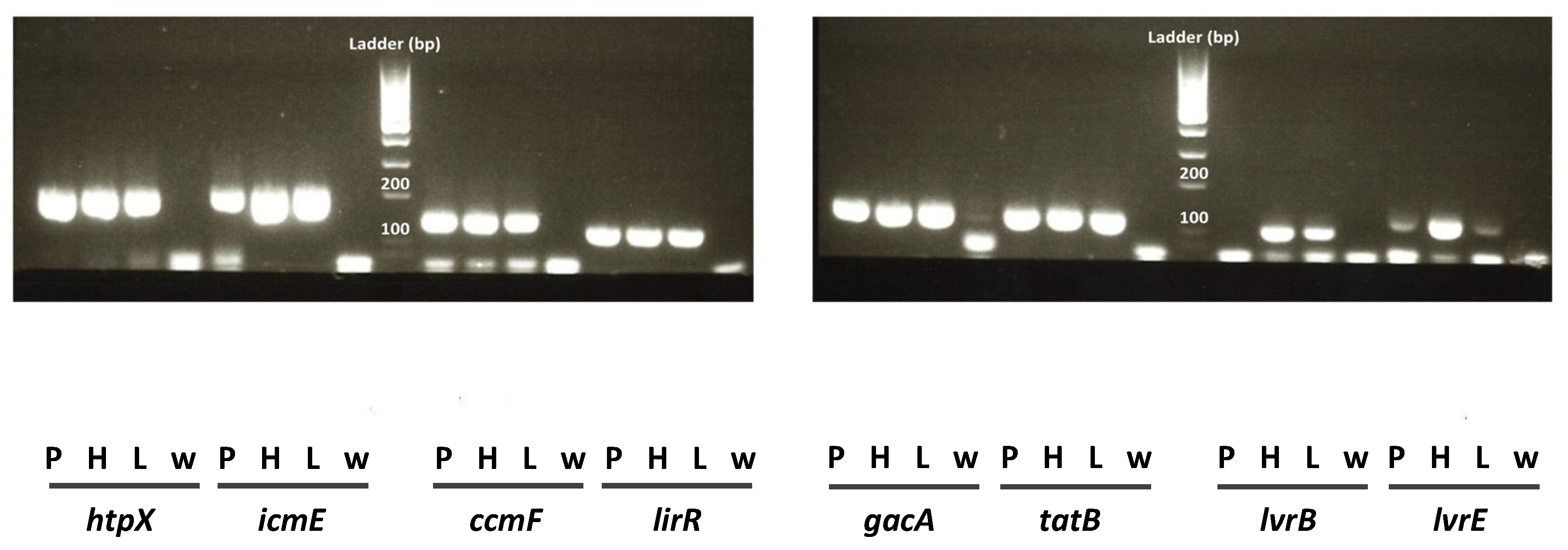
2.1. Virulence Gene Selection
2.2. Validation of rpsL as a Non-Acceptable Housekeeping Gene, in Which Transcript Level Should Be Stable between the Different Tested Conditions
2.3. Definition of a Gene Expression-Based Virulence Index
2.4. Comparison of the Virulence Index of three L. pneumophila Strains
3. Discussion
4. Materials and Methods
4.1. L. pneumophila Cultures
4.2. FLA Culture
4.3. Coculture of L. pneumophila with Amoebae
4.4. Preservation of RNA Samples
4.5. Total RNA Extraction
4.6. Calibrated Reverse Transcription (cRT)
4.7. Quantitative PCR (qPCR)
4.8. Calculation of the Virulence Index
4.9. Data and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cunha, C.B.; Cunha, B.A. Legionnaire’s Disease Since Philadelphia. Infect. Dis. Clin. N. Am. 2017, 31, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Khodr, A.; Kay, E.; Gomez-Valero, L.; Ginevra, C.; Doublet, P.; Buchrieser, C.; Jarraud, S. Molecular epidemiology, phylogeny and evolution of Legionella. Infect. Genet. Evol. 2016, 43, 108–122. [Google Scholar] [CrossRef] [PubMed]
- Rossier, O.; Starkenburg, S.R.; Cianciotto, N.P. Legionella pneumophila Type II Protein Secretion Promotes Virulence in the A/J Mouse Model of Legionnaires’ Disease Pneumonia. Infect. Immun. 2004, 72, 310–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowbotham, T.J. Preliminary report on the pathogenicity of Legionella pneumophila for freshwater and soil amoebae. J. Clin. Pathol. 1980, 33, 1179–1183. [Google Scholar] [CrossRef] [Green Version]
- Balczun, C.; Scheid, P. Free-Living Amoebae as Hosts for and Vectors of Intracellular Microorganisms with Public Health Significance. Viruses 2017, 9, 65. [Google Scholar] [CrossRef] [PubMed]
- Berjeaud, J.-M.; Chevalier, S.; Schlusselhuber, M.; Portier, E.; Loiseau, C.; Aucher, W.; Lesouhaitier, O.; Verdon, J. Legionella pneumophila: The Paradox of a Highly Sensitive Opportunistic Waterborne Pathogen Able to Persist in the Environment. Front. Microbiol. 2016, 7, 21660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borella, P.; Guerrieri, E.; Marchesi, I.; Bondi, M.; Messi, P. Water ecology of Legionella and protozoan: Environmental and public health perspectives. Biotechnol. Annu. Rev. 2005, 11, 355–380. [Google Scholar] [CrossRef]
- Neumeister, B.; Reiff, G.; Faigle, M.; Dietz, K.; Northoff, H.; Lang, F. Influence of Acanthamoeba castellanii on Intracellular Growth of Different Legionella Species in Human Monocytes. Appl. Environ. Microbiol. 2000, 66, 914–919. [Google Scholar] [CrossRef] [Green Version]
- Cirillo, J.D.; Falkow, S.; Tompkins, L.S. Growth of Legionella pneumophila in Acanthamoeba castellanii enhances invasion. Infect. Immun. 1994, 62, 3254–3261. [Google Scholar] [CrossRef] [Green Version]
- Cirillo, J.D.; Cirillo, S.L.G.; Yan, L.; Bermudez, L.E.; Falkow, S.; Tompkins, L.S. Intracellular Growth in Acanthamoeba castellanii Affects Monocyte Entry Mechanisms and Enhances Virulence of Legionella pneumophila. Infect. Immun. 1999, 67, 4427–4434. [Google Scholar] [CrossRef] [Green Version]
- Michard, C.; Doublet, P. Post-translational modifications are key players of the Legionella pneumophila infection strategy. Front. Microbiol. 2015, 6, 87. [Google Scholar] [CrossRef] [PubMed]
- Prashar, A.; Terebiznik, M.R. Legionella pneumophila: Homeward bound away from the phagosome. Curr. Opin. Microbiol. 2015, 23, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Zink, S.; Pedersen, L.; Cianciotto, N.P.; Abu Kwaik, Y. The Dot/Icm Type IV Secretion System of Legionella pneumophila Is Essential for the Induction of Apoptosis in Human Macrophages. Infect. Immun. 2002, 70, 1657–1663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, T.S.; Gjiknuri, J.; Magnet, A.; Vaccaro, L.; Ollero, D.; Izquierdo, F.; Fenoy, S.; Hurtado, C.; Del Águila, C. The Influence of Acanthamoeba–Legionella Interaction in the Virulence of Two Different Legionella Species. Front. Microbiol. 2018, 9, 2962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aurass, P.; Schlegel, M.; Metwally, O.; Harding, C.R.; Schroeder, G.; Frankel, G.; Flieger, A. The Legionella pneumophila Dot/Icm-secreted Effector PlcC/CegC1 Together with PlcA and PlcB Promotes Virulence and Belongs to a Novel Zinc Metallophospholipase C Family Present in Bacteria and Fungi. J. Boil. Chem. 2013, 288, 11080–11092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harding, C.R.; Stoneham, C.; Schuelein, R.; Newton, H.J.; Oates, C.V.; Hartland, E.L.; Schroeder, G.; Frankel, G. The Dot/Icm Effector SdhA Is Necessary for Virulence of Legionella pneumophila in Galleria mellonella and A/J Mice. Infect. Immun. 2013, 81, 2598–2605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontana, M.F.; Banga, S.; Barry, K.; Shen, X.; Tan, Y.; Luo, Z.-Q.; E Vance, R. Secreted Bacterial Effectors That Inhibit Host Protein Synthesis Are Critical for Induction of the Innate Immune Response to Virulent Legionella pneumophila. PLoS Pathog. 2011, 7, e1001289. [Google Scholar] [CrossRef]
- Buse, H.Y.; Lu, J.; Ashbolt, N.J. Exposure to Synthetic Gray Water Inhibits Amoeba Encystation and Alters Expression of Legionella pneumophila Virulence Genes. Appl. Environ. Microbiol. 2015, 81, 630–639. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Struewing, I.; Buse, H.Y.; Kou, J.; Shuman, H.A.; Faucher, S.P.; Ashbolt, N.J. Legionella pneumophila Transcriptional Response following Exposure to CuO Nanoparticles. Appl. Environ. Microbiol. 2013, 79, 2713–2720. [Google Scholar] [CrossRef] [Green Version]
- Hasni, I.; Jarry, A.; Quelard, B.; Carlino, A.; Eberst, J.-B.; Abbe, O.; Demanèche, S. Intracellular Behaviour of Three Legionella pneumophila Strains within Three Amoeba Strains, Including Willaertia magna C2c Maky. Pathogens 2020, 9, 105. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, G.; Petty, N.K.; Mousnier, A.; Harding, C.R.; Vogrin, A.J.; Wee, B.; Fry, N.K.; Harrison, T.G.; Newton, H.J.; Thomson, N.R.; et al. Legionella pneumophila Strain 130b Possesses a Unique Combination of Type IV Secretion Systems and Novel Dot/Icm Secretion System Effector Proteins. J. Bacteriol. 2010, 192, 6001–6016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xuereb, B.; Bezin, L.; Chaumot, A.; Budzinski, H.; Augagneur, S.; Tutundjian, R.; Garric, J.; Geffard, O. Vitellogenin-like gene expression in freshwater amphipod Gammarus fossarum (Koch, 1835): Functional characterization in females and potential for use as an endocrine disruption biomarker in males. Ecotoxicology 2011, 20, 1286–1299. [Google Scholar] [CrossRef] [PubMed]
- Dey, R.; Bodennec, J.; Mameri, M.O.; Pernin, P. Free-living freshwater amoebae differ in their susceptibility to the pathogenic bacteriumLegionella pneumophila. FEMS Microbiol. Lett. 2009, 290, 10–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vranckx, L.; De Buck, E.; Anné, J.; Lammertyn, E. Legionella pneumophila exhibits plasminogen activator activity. Microbiology 2007, 153, 3757–3765. [Google Scholar] [CrossRef] [Green Version]
- Zusman, T.; Degtyar, E.; Segal, G. Identification of a Hypervariable Region Containing New Legionella pneumophila Icm/Dot Translocated Substrates by Using the Conserved icmQ Regulatory Signature. Infect. Immun. 2008, 76, 4581–4591. [Google Scholar] [CrossRef] [Green Version]
- Cianciotto, N.P. Many substrates and functions of type II secretion: Lessons learned fromLegionella pneumophila. Futur. Microbiol. 2009, 4, 797–805. [Google Scholar] [CrossRef] [Green Version]
- Banga, S.; Gao, P.; Shen, X.; Fiscus, V.; Zong, W.-X.; Chen, L.; Luo, Z.-Q. Legionella pneumophila inhibits macrophage apoptosis by targeting pro-death members of the Bcl2 protein family. Proc. Natl. Acad. Sci. USA 2007, 104, 5121–5126. [Google Scholar] [CrossRef] [Green Version]
- Hsu, F.; Zhu, W.; Brennan, L.; Tao, L.; Luo, Z.-Q.; Mao, Y. Structural basis for substrate recognition by a unique Legionella phosphoinositide phosphatase. Proc. Natl. Acad. Sci. USA 2012, 109, 13567–13572. [Google Scholar] [CrossRef] [Green Version]
- Bosshardt, S.C.; Benson, R.F.; Fields, B.S. Flagella are a positive predictor for virulence inLegionella. Microb. Pathog. 1997, 23, 107–112. [Google Scholar] [CrossRef]
- Nowicki, M.; Bornstein, N.; Paucod, J.; Binder, P.; Fleurette, J. Effect of Culture Medium on Morphology and Virulence of Legionella pneumophila Serogroup 1. Zent. Bakteriol. Mikrobiol. Hyg. 1987, 264, 167–177. [Google Scholar] [CrossRef]
- De Jonckheere, J. Use of an axenic medium for differentiation between pathogenic and nonpathogenic Naegleria fowleri isolates. Appl. Environ. Microbiol. 1977, 33, 751–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laskowski-Arce, M.A.; Orth, K. Acanthamoeba castellanii Promotes the Survival of Vibrio parahaemolyticus. Appl. Environ. Microbiol. 2008, 74, 7183–7188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustin, S.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.F.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bezin, L.; Morales, A. Method of Calibration of Reverse Transcription Using a Synthetic Messenger RNA (SmRNA) as an Internal Control. Patent WO20044092414, 28 October 2004. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Gene ID | Size (bp) | Forward Primer (5′->3′) | Reverse Primer (5′->3′) | a | E |
|---|---|---|---|---|---|
| ccmF | 149 | TGA ATA CAC AGG GCC GTG ATC TGA | ACT GGT TTC TAC TTT CCC TGC CCA | −3.369 | 1.98 |
| dotA | 81 | CTG AGA TGG ATA GGT GGT AGT C | TCT TAC TCT ACC TTT GGC TTC CTC | −3.473 | 1.94 |
| enhC | 438 | AAT GCT TTG TAT GCC CTC GG | CAT ATC AGC GCT TTG GCC ATC | −3.401 | 1.97 |
| gacA | 120 | TTT AAA CGA CGC GTC ACT TCC CAC | TGC AGA TGC TGA AAG TGG TGA GCA | −3.386 | 1.97 |
| htpX | 196 | ATT GAC TCT CAT GGT TGC CGT GCT | AGC CAT GTA TTC TCT GGT TCG GCT | −3.330 | 2.00 |
| icmE | 200 | GCT CAA ATC AAA GCT GCT CAG GCA | CCT GCG TTT GCT AAA TCC GCA TCA | −3.331 | 2.00 |
| lirR | 124 | CCA TGC TTA ATG CTC TCT ACC A | GGG TTG CTC CGC AAT TAA AC | −3.541 | 1.92 |
| lvrB | 99 | CAT TGG TGT ACT CTC GGT CTT C | AGC ACC ATG CAG AGC ATA C | −3.385 | 1.97 |
| lvrE | 128 | CCG TAA CAA GTG GGT GAT TCT | CAT TGC CCA ACA AAC CAT AGA C | −3.330 | 2.00 |
| rpsL | 132 | GAA AGC CTC GTG TGG ACG TA | CAA CCT TAC GCA TAG CTG AGT TA | −3.340 | 1.99 |
| tatB | 115 | ATT GTG TTT GGG CCA TCA AAG | CAT TGA GTT GTT GCT GCC AAA | −3.484 | 1.94 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mameri, R.M.; Bodennec, J.; Bezin, L.; Demanèche, S. Mitigation of Expression of Virulence Genes in Legionella pneumophila Internalized in the Free-Living Amoeba Willaertia magna C2c Maky. Pathogens 2020, 9, 447. https://doi.org/10.3390/pathogens9060447
Mameri RM, Bodennec J, Bezin L, Demanèche S. Mitigation of Expression of Virulence Genes in Legionella pneumophila Internalized in the Free-Living Amoeba Willaertia magna C2c Maky. Pathogens. 2020; 9(6):447. https://doi.org/10.3390/pathogens9060447
Chicago/Turabian StyleMameri, Rayane Mouh, Jacques Bodennec, Laurent Bezin, and Sandrine Demanèche. 2020. "Mitigation of Expression of Virulence Genes in Legionella pneumophila Internalized in the Free-Living Amoeba Willaertia magna C2c Maky" Pathogens 9, no. 6: 447. https://doi.org/10.3390/pathogens9060447