Contrasted Epidemiological Patterns of West Nile Virus Lineages 1 and 2 Infections in France from 2015 to 2019

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Comparison of WNV Seasonal Patterns in France between 2015 and 2019
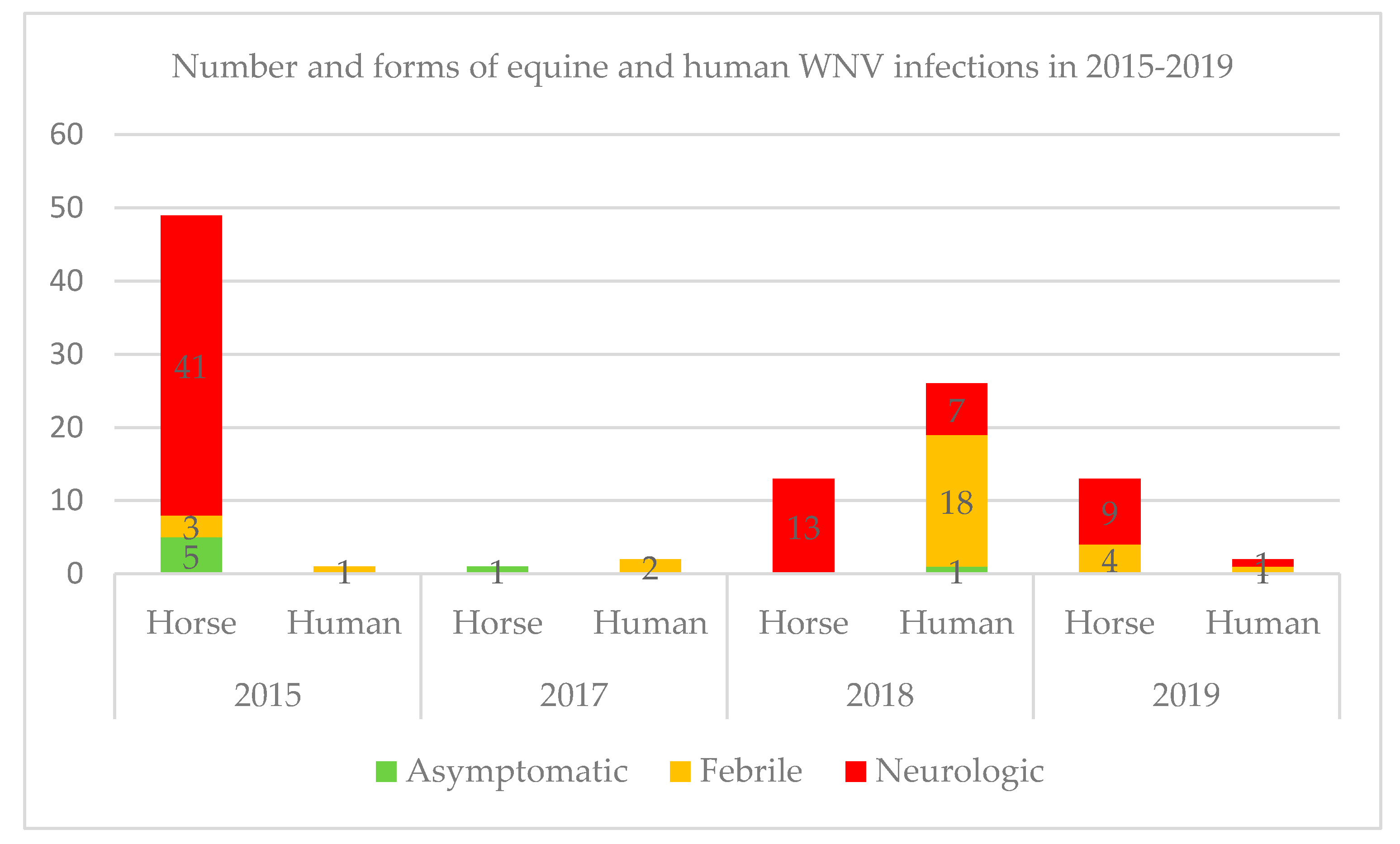
2.1.1. WNV Outbreaks in France in 2015–2019
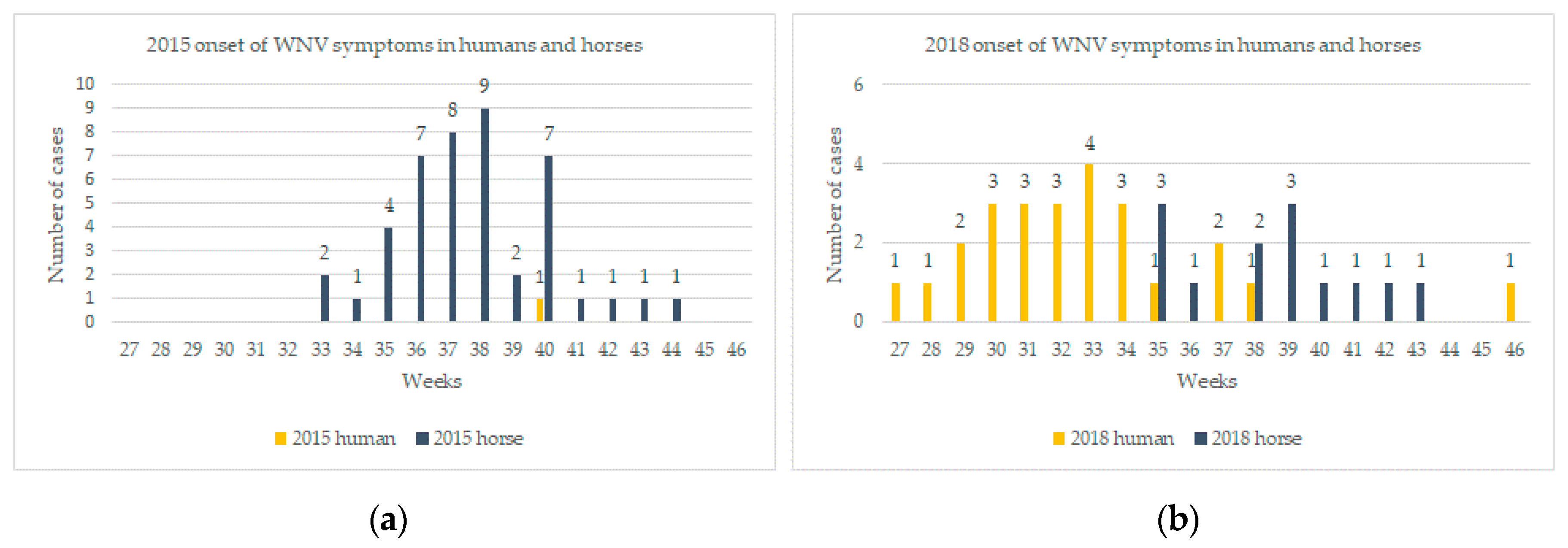
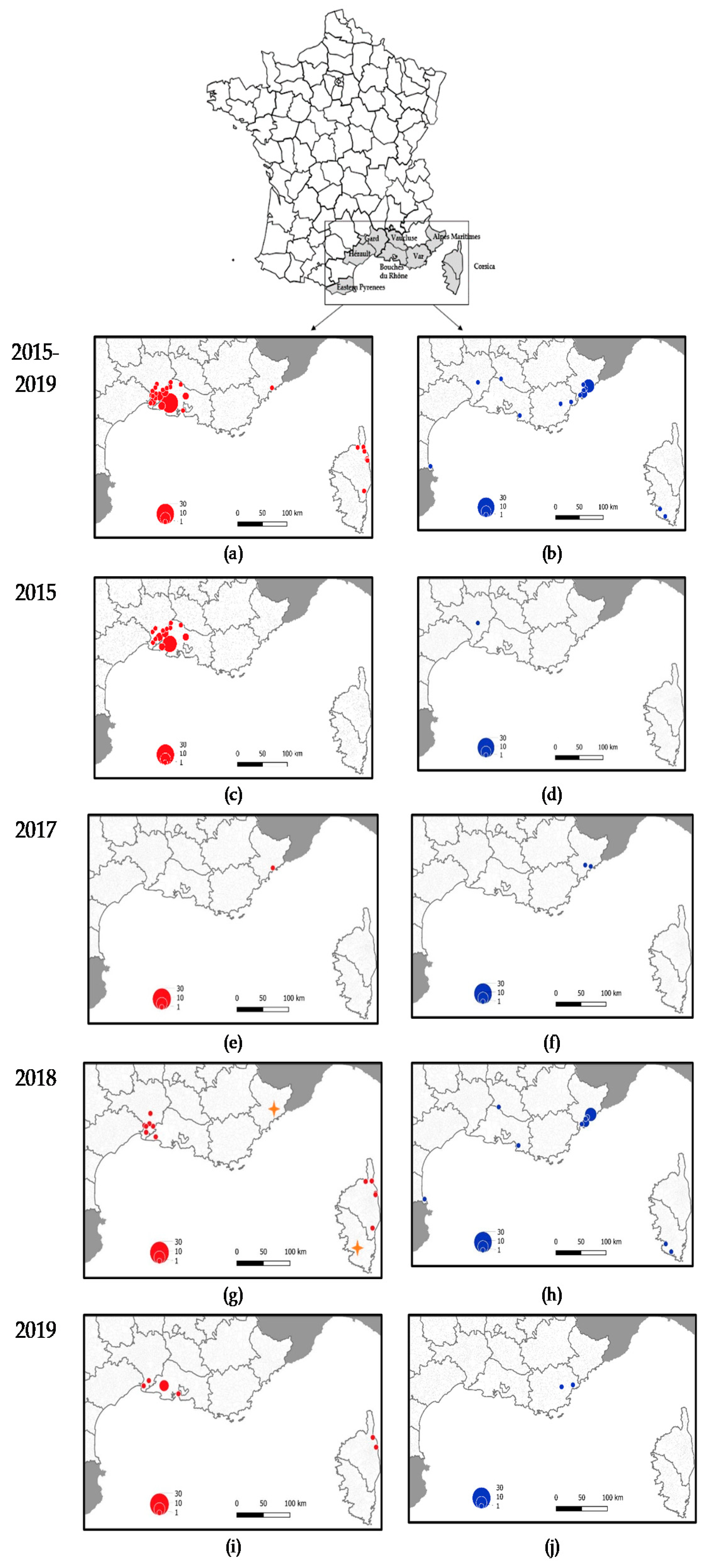
2.1.2. Shifts in Temporal and Spatial Distribution of WNV Cases
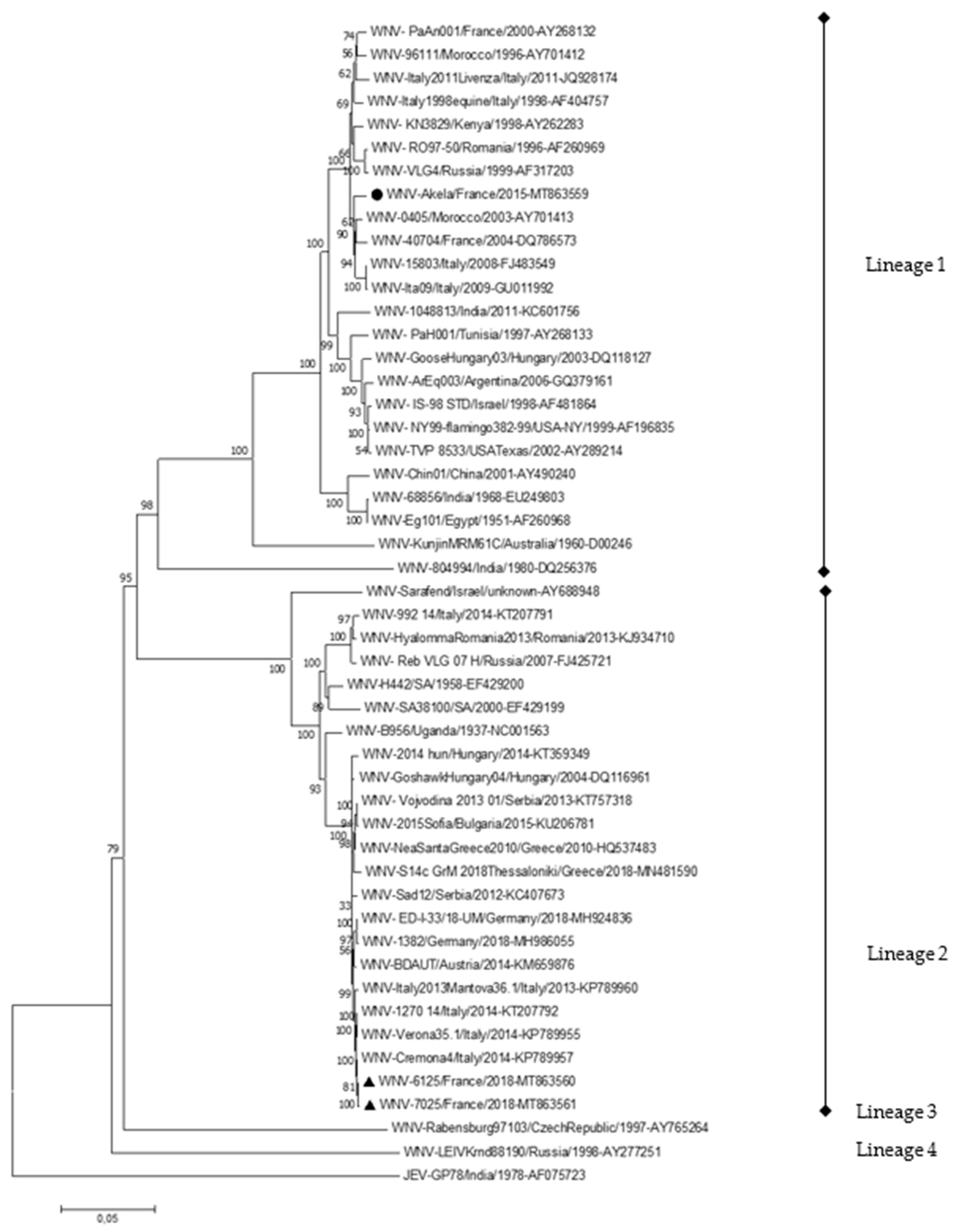
2.2. First Description of WNV Lineage 2 Isolates in France in 2018
3. Discussion
4. Materials and Methods
4.1. Samples
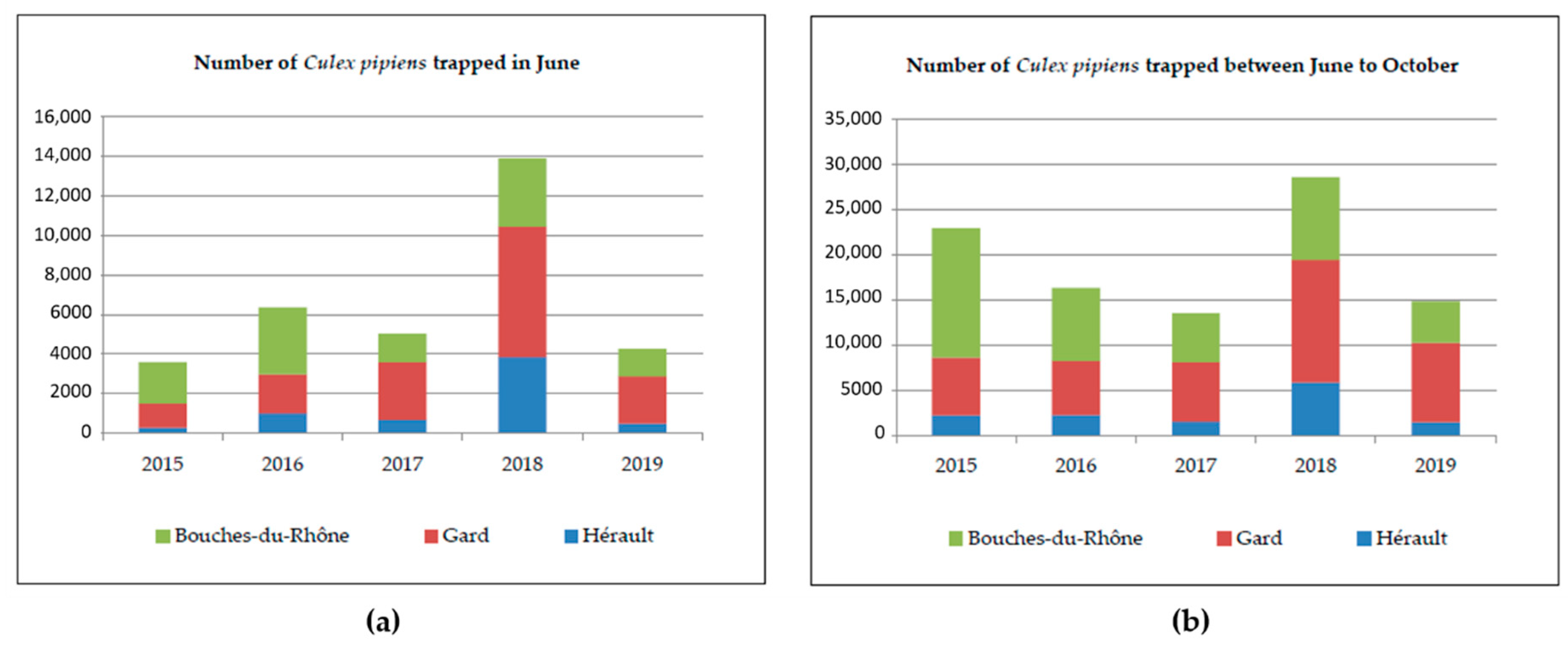
4.2. Mosquito Collection
4.3. Serology
4.4. Real-Time RT-PCR
4.5. Virus Isolation
4.6. Nucleotide Sequencing and Sequence Analysis
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hubalek, Z.; Halouzka, J. West Nile fever—A reemerging mosquito-borne viral disease in Europe. Emerg. Infect. Dis. 1999, 5, 643–650. [Google Scholar] [CrossRef]
- Kramer, L.D.; Li, J.; Shi, P.-Y. West Nile virus. Lancet Neurol. 2007, 6, 171–181. [Google Scholar] [CrossRef]
- Perez-Ramirez, E.; Llorente, F.; del Amo, J.; Fall, G.; Sall, A.A.; Lubisi, A.; Lecollinet, S.; Vazquez, A.; Jimenez-Clavero, M.A. Pathogenicity evaluation of twelve West Nile virus strains belonging to four lineages from five continents in a mouse model: Discrimination between three pathogenicity categories. J. Gen. Virol. 2017, 98, 662–670. [Google Scholar] [CrossRef]
- Beck, C.; Jimenez-Clavero, M.A.; Leblond, A.; Durand, B.; Nowotny, N.; Leparc-Goffart, I.; Zientara, S.; Jourdain, E.; Lecollinet, S. Flaviviruses in Europe: Complex circulation patterns and their consequences for the diagnosis and control of West Nile disease. Int. J. Environ. Res. Public Health 2013, 10, 6049–6083. [Google Scholar] [CrossRef] [Green Version]
- Joubert, L.; Oudar, J.; Hannoun, C.; Beytout, D.; Corniou, B.; Guillon, J.C.; Panthier, R. Epidemiology of the West Nile virus: Study of a focus in Camargue. Iv. Meningo-Encephalomyelitis of the horse. Ann. Inst. Pasteur 1970, 118, 239–247. [Google Scholar]
- Filipe, A.R.; Pinto, M.R. Survey for antibodies to arboviruses in serum of animals from Southern Portugal. Am. J. Trop. Med. Hyg. 1969, 18, 423–426. [Google Scholar] [CrossRef] [PubMed]
- Murgue, B.; Murri, S.; Triki, H.; Deubel, V.; Zeller, H.G. West Nile in the Mediterranean basin: 1950–2000. Ann. N. Y. Acad. Sci. 2001, 951, 117–126. [Google Scholar] [CrossRef]
- Sotelo, E.; Fernandez-Pinero, J.; Llorente, F.; Vazquez, A.; Moreno, A.; Aguero, M.; Cordioli, P.; Tenorio, A.; Jimenez-Clavero, M.A. Phylogenetic relationships of Western Mediterranean West Nile virus strains (1996–2010) using full-length genome sequences: Single or multiple introductions? J. Gen. Virol. 2001, 92, 2512–2522. [Google Scholar] [CrossRef] [PubMed]
- Autorino, G.L.; Battisti, A.; Deubel, V.; Ferrari, G.; Forletta, R.; Giovannini, A.; Lelli, R.; Murri, S.; Scicluna, M.T. West Nile virus epidemic in horses, Tuscany Region, Italy. Emerg. Infect. Dis. 2002, 8, 1372–1378. [Google Scholar] [CrossRef] [PubMed]
- Ceianu, C.S.; Ungureanu, A.; Nicolescu, G.; Cernescu, C.; Nitescu, L.; Tardei, G.; Petrescu, A.; Pitigoi, D.; Martin, D.; Ciulacu-Purcarea, V.; et al. West Nile virus surveillance in Romania: 1997–2000. Viral Immunol. 2001, 14, 251–262. [Google Scholar] [CrossRef]
- Bahuon, C.; Marcillaud-Pitel, C.; Bournez, L.; Leblond, A.; Beck, C.; Hars, J.; Leparc-Goffart, I.; L’Ambert, G.; Paty, M.C.; Cavalerie, L.; et al. West Nile virus epizootics in the Camargue (France) in 2015 and reinforcement of surveillance and control networks. Rev. Sci. Tech. 2016, 35, 811–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, N.; de Marco, M.F.; Giovannini, A.; Ippoliti, C.; Danzetta, M.L.; Svartz, G.; Erster, O.; Groschup, M.H.; Ziegler, U.; Mirazimi, A.; et al. Emerging mosquito-borne threats and the response from European and Eastern Mediterranean countries. Int. J. Environ. Res. Public Health 2018, 15, 2775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Giudice, P.; Schuffenecker, I.; Vandenbos, F.; Counillon, E.; Zellet, H. Human West Nile virus, France. Emerg. Infect. Dis. 2004, 10, 1885–1886. [Google Scholar] [CrossRef] [PubMed]
- Lecollinet, S.; Pronost, S.; Coulpier, M.; Beck, C.; Gonzalez, G.; Leblond, A.; Tritz, P. Viral equine encephalitis, a growing threat to the horse population in Europe? Viruses 2019, 12, 23. [Google Scholar] [CrossRef] [Green Version]
- de Heus, P.; Kolodziejek, J.; Camp, J.V.; Dimmel, K.; Bago, Z.; Hubalek, Z.; van den Hoven, R.; Cavalleri, J.V.; Nowotny, N. Emergence of West Nile virus lineage 2 in Europe: Characteristics of the first seven cases of West Nile neuroinvasive disease in horses in Austria. Transbound. Emerg. Dis. 2019, 67, 1189–1197. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Triana, L.M.; Jeffries, C.L.; Mansfield, K.L.; Carnell, G.; Fooks, A.R.; Johnson, N. Emergence of West Nile virus lineage 2 in Europe: A review on the introduction and spread of a mosquito-borne disease. Front. Public Health 2014, 2, 271. [Google Scholar] [CrossRef] [Green Version]
- Bakonyi, T.; Ivanics, E.; Erdelyi, K.; Ursu, K.; Ferenczi, E.; Weissenbock, H.; Nowotny, N. Lineage 1 and 2 strains of encephalitic West Nile virus, Central Europe. Emerg. Infect. Dis. 2006, 12, 618–623. [Google Scholar] [CrossRef]
- Bakonyi, T.; Ferenczi, E.; Erdelyi, K.; Kutasi, O.; Csorgo, T.; Seidel, B.; Weissenbock, H.; Brugger, K.; Ban, E.; Nowotny, N. Explosive spread of a neuroinvasive lineage 2 West Nile virus in Central Europe, 2008/2009. Vet. Microbiol. 2013, 165, 61–70. [Google Scholar] [CrossRef]
- Papa, A.; Danis, K.; Baka, A.; Bakas, A.; Dougas, G.; Lytras, T.; Theocharopoulos, G.; Chrysagis, D.; Vassiliadou, E.; Kamaria, F.; et al. Ongoing outbreak of West Nile virus infections in humans in Greece, July–August 2010. Eurosurveillance 2010, 15, 19644. [Google Scholar] [CrossRef]
- Napp, S.; Petric, D.; Busquets, N. West Nile virus and other mosquito-borne viruses present in Eastern Europe. Pathog. Glob. Health 2018, 112, 233–248. [Google Scholar] [CrossRef]
- Merdic, E.; Peric, L.; Pandak, N.; Kurolt, I.C.; Turic, N.; Vignjevic, G.; Stolfa, I.; Milas, J.; Bogojevic, M.S.; Markotic, A. West Nile virus outbreak in humans in Croatia, 2012. Coll. Antropol. 2013, 37, 943–947. [Google Scholar] [PubMed]
- Savini, G.; Capelli, G.; Monaco, F.; Polci, A.; Russo, F.; di Gennaro, A.; Marini, V.; Teodori, L.; Montarsi, F.; Pinoni, C.; et al. Evidence of West Nile virus lineage 2 circulation in Northern Italy. Vet. Microbiol. 2012, 158, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Busquets, N.; Laranjo-Gonzalez, M.; Soler, M.; Nicolas, O.; Rivas, R.; Talavera, S.; Villalba, R.; Miguel, E.S.; Torner, N.; Aranda, C.; et al. Detection of West Nile virus lineage 2 in North-Eastern Spain (Catalonia). Transbound. Emerg. Dis. 2019, 66, 617–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michel, F.; Sieg, M.; Fischer, D.; Keller, M.; Eiden, M.; Reuschel, M.; Schmidt, V.; Schwehn, R.; Rinder, M.; Urbaniak, S.; et al. Evidence for West Nile virus and Usutu virus infections in wild and resident birds in Germany, 2017 and 2018. Viruses 2019, 11, 674. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, U.; Luhken, R.; Keller, M.; Cadar, D.; van der Grinten, E.; Michel, F.; Albrecht, K.; Eiden, M.; Rinder, M.; Lachmann, L.; et al. West Nile virus epizootic in Germany, 2018. Antiviral. Res. 2019, 162, 39–43. [Google Scholar] [CrossRef]
- Platonov, A.E.; Karan, L.S.; Shopenskaia, T.A.; Fedorova, M.V.; Koliasnikova, N.M.; Rusakova, N.M.; Shishkina, L.V.; Arshba, T.E.; Zhuravlev, V.I.; Govorukhina, M.V.; et al. Genotyping of West Nile fever virus strains circulating in Southern Russia as an epidemiological investigation method: Principles and Results. Zhurnal Mikrobiol. Epidemiol. Immunobiol. 2011, 2, 29–37. [Google Scholar]
- Cotar, A.I.; Falcuta, E.; Dinu, S.; Necula, A.; Birlutiu, V.; Ceianu, C.S.; Prioteasa, F.L. West Nile virus lineage 2 in Romania, 2015–2016: Co-circulation and strain replacement. Parasit. Vectors 2018, 11, 562. [Google Scholar] [CrossRef] [Green Version]
- Ravagnan, S.; Montarsi, F.; Cazzin, S.; Porcellato, E.; Russo, F.; Palei, M.; Monne, I.; Savini, G.; Marangon, S.; Barzon, L.; et al. First report outside eastern europe of West Nile virus lineage 2 related to the Volgograd 2007 Strain, Northeastern Italy, 2014. Parasit. Vectors 2015, 8, 418. [Google Scholar] [CrossRef] [Green Version]
- Papa, A.; Papadopoulou, E.; Chatzixanthouliou, C.; Glouftsios, P.; Pappa, S.; Pervanidou, D.; Georgiou, L. Emergence of West Nile virus lineage 2 belonging to the Eastern European Subclade, Greece. Arch. Virol. 2019, 164, 1673–1675. [Google Scholar] [CrossRef]
- Saegerman, C.; Alba-Casals, A.; Garcia-Bocanegra, I.; Pozzo, F.D.; van Galen, G. Clinical sentinel surveillance of equine West Nile fever, Spain. Transbound. Emerg. Dis. 2016, 63, 184–193. [Google Scholar] [CrossRef]
- Ramalli, L.; Grard, G.; Beck, C.; Gallian, P.; L’Ambert, G.; Desvaux, S.; Jourdan, M.; Ortmans, C.; Paty, M.C.; Franke, F. West Nile virus infections in France, July to November 2018. Eur. J. Public Health 2019, 29, ckz186.631. [Google Scholar] [CrossRef]
- Leblond, A.; Hendrikx, P.; Sabatier, P. West Nile virus outbreak detection using syndromic monitoring in horses. Vector Borne Zoonotic Dis. 2007, 7, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Durand, B.; Dauphin, G.; Zeller, H.; Labie, J.; Schuffenecker, I.; Murri, S.; Moutou, F.; Zientara, S. Serosurvey for West Nile virus in horses in Southern France. Vet. Rec. 2005, 157, 711–713. [Google Scholar] [CrossRef] [PubMed]
- Paz, S.; Semenza, J.C. Environmental drivers of West Nile fever epidemiology in Europe and Western Asia—A review. Int. J. Environ. Res. Public Health 2013, 10, 3543–3562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calzolari, M.; Angelini, P.; Bolzoni, L.; Bonilauri, P.; Cagarelli, R.; Canziani, S.; Cereda, D.; Cerioli, M.P.; Chiari, M.; Galletti, G.; et al. Enhanced West Nile virus circulation in the Emilia-Romagna and Lombardy Regions (Northern Italy) in 2018 detected by entomological surveillance. Front. Vet. Sci. 2020, 7, 243. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. Mega7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Chaintoutis, S.C.; Papa, A.; Pervanidou, D.; Dovas, C.I. Evolutionary dynamics of lineage 2 West Nile virus in Europe, 2004–2018: Phylogeny, selection pressure and phylogeography. Mol. Phylogenet. Evol. 2019, 141, 106617. [Google Scholar] [CrossRef]
- Kaiser, J.A.; Wang, T.; Barrett, A.D. Virulence determinants of West Nile virus: How can these be used for vaccine design? Future Virol. 2019, 12, 283–295. [Google Scholar] [CrossRef] [Green Version]
- Charrel, R.N.; Brault, A.C.; Gallian, P.; Lemasson, J.J.; Murgue, B.; Murri, S.; Pastorino, B.; Zeller, H.; de Chesse, R.; de Micco, P.; et al. Evolutionary relationship between old world West Nile virus strains. evidence for viral gene flow between Africa, the Middle East, and Europe. Virology 2003, 315, 381–388. [Google Scholar] [CrossRef] [Green Version]
- Jourdain, E.; Schuffenecker, I.; Korimbocus, J.; Reynard, S.; Murri, S.; Kayser, Y.; Gauthier-Clerc, M.; Sabatier, P.; Zeller, H.G. West Nile virus in wild resident birds, Southern France, 2004. Vector Borne Zoonotic Dis. 2007, 7, 448–452. [Google Scholar] [CrossRef]
- Bahuon, C.; Pitel, C.M.; Bournez, L.; Leblond, A.; Hars, J.; Beck, C.; Goffart, I.L.; L’Ambert, G.; Paty, M.C.; Cavalerie, L.; et al. Wnv epizootics in Camargue, France, 2015 and reinforcement of Wnv surveillance and control networks. OIE Bull. Épidémiol. 2016. [Google Scholar] [CrossRef]
- Murgue, B.; Murri, S.; Zientara, S.; Durand, B.; Durand, J.P.; Zeller, H. West Nile outbreak in horses in Southern France, 2000: The return after 35 years. Emerg. Infect. Dis. 2001, 7, 692–696. [Google Scholar] [CrossRef]
- Pradier, S.; Sandoz, A.; Paul, M.C.; Lefebvre, G.; Tran, A.; Maingault, J.; Lecollinet, S.; Leblond, A. Importance of wetlands management for West Nile virus circulation risk, Camargue, Southern France. Int. J. Environ. Res. Public Health 2014, 11, 7740–7754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vittecoq, M.; Lecollinet, S.; Jourdain, E.; Thomas, F.; Blanchon, T.; Arnal, A.; Lowenski, S.; Gauthier-Clerc, M. Recent circulation of West Nile virus and potentially other closely related flaviviruses in Southern France. Vector Borne Zoonotic Dis. 2013, 13, 610–613. [Google Scholar] [CrossRef]
- Balanca, G.; Gaidet, N.; Savini, G.; Vollot, B.; Foucart, A.; Reiter, P.; Boutonnier, A.; Lelli, R.; Monicat, F. Low West Nile virus circulation in wild birds in an area of recurring outbreaks in Southern France. Vector Borne Zoonotic Dis. 2009, 9, 737–741. [Google Scholar] [CrossRef]
- McMullen, A.R.; Albayrak, H.; May, F.J.; Davis, C.T.; Beasley, D.W.C.; Barrett, A.D.T. Molecular evolution of lineage 2 West Nile virus. J. Gen. Virol. 2013, 94, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, U.; Santos, P.D.; Groschup, M.H.; Hattendorf, C.; Eiden, M.; Hoper, D.; Eisermann, P.; Keller, M.; Michel, F.; Klopfleisch, R.; et al. West Nile virus epidemic in Germany triggered by epizootic emergence, 2019. Viruses 2020, 12, 448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Ramirez, E.; Llorente, F.; Jimenez-Clavero, M.A. Experimental infections of wild birds with West Nile virus. Viruses 2014, 6, 752–781. [Google Scholar] [CrossRef] [Green Version]
- Vidaña, B.; Busquets, N.; Napp, S.; Pérez-Ramírez, E.; Jiménez-Clavero, M.; Johnson, N. The role of birds of prey in West Nile virus epidemiology. Vaccines 2020, 8, 550. [Google Scholar] [CrossRef]
- Zana, B.; Erdelyi, K.; Nagy, A.; Mezei, E.; Nagy, O.; Takacs, M.; Bakonyi, T.; Forgach, P.; Korbacska-Kutasi, O.; Feher, O.; et al. Multi-approach investigation regarding the West Nile virus situation in Hungary, 2018. Viruses 2020, 12, 123. [Google Scholar] [CrossRef] [Green Version]
- Haussig, J.M.; Young, J.J.; Gossner, C.M.; Mezei, E.; Bella, A.; Sirbu, A.; Pervanidou, D.; Drakulovic, M.B.; Sudre, B. Early start of the West Nile fever transmission season 2018 in Europe. Eurosurveillance 2018, 23, 1800428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ECDC. West Nile Virus Infections by Affected Areas, in the EU/EEA Member States and EU Neighbouring Countries, 2018 Transmission Season. Available online: https://www.ecdc.europa.eu/en/publications-data/west-nile-virus-infections-affected-areas-eueea-member-states-and-eu-neighbouring (accessed on 14 May 2020).
- ECDC. Epidemiological Update: West Nile Virus Transmission Season in Europe. 2018. Available online: https://www.ecdc.europa.eu/en/news-events/epidemiological-update-west-nile-virus-transmission-season-europe-2018 (accessed on 14 December 2018).
- Marini, G.; Calzolari, M.; Angelini, P.; Bellini, R.; Bellini, S.; Bolzoni, L.; Torri, D.; Defilippo, F.; Dorigatti, I.; Nikolay, B.; et al. A quantitative comparison of West Nile virus incidence from 2013 to 2018 in Emilia-Romagna, Italy. PLoS Negl. Trop. Dis. 2020, 14, e0007953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veo, C.; della Ventura, C.; Moreno, A.; Rovida, F.; Percivalle, E.; Canziani, S.; Torri, D.; Calzolari, M.; Baldanti, F.; Galli, M.; et al. Evolutionary dynamics of the lineage 2 West Nile virus that caused the largest European epidemic: Italy 2011–2018. Viruses 2019, 11, 814. [Google Scholar] [CrossRef] [Green Version]
- Langevin, S.A.; Bowen, R.A.; Reisen, W.K.; Andrade, C.C.; Ramey, W.N.; Maharaj, P.D.; Anishchenko, M.; Kenney, J.L.; Duggal, N.K.; Romo, H.; et al. Host competence and helicase activity differences exhibited by West Nile viral variants expressing Ns3-249 amino acid polymorphisms. PLoS ONE 2014, 9, e100802. [Google Scholar] [CrossRef]
- Dridi, M.; van den Berg, T.; Lecollinet, S.; Lambrecht, B. Evaluation of the pathogenicity of West Nile virus (WNV) lineage 2 strains in a SPF chicken model of infection: Ns3-249Pro mutation is neither sufficient nor necessary for conferring virulence. Vet. Res. 2015, 46, 130. [Google Scholar] [CrossRef] [Green Version]
- Durand, B.; Tran, A.; Balanca, G.; Chevalier, V. Geographic variations of the bird-borne structural risk of West Nile virus circulation in Europe. PLoS ONE 2017, 12, e0185962. [Google Scholar] [CrossRef] [Green Version]
- IFCE. Tableau Économique, Statistique et Graphique du Cheval en France Données 2016/2017. 2017. Available online: https://www.ifce.fr/wp-content/uploads/2017/11/OESC-Annuaire-ECUS-2017.pdf (accessed on 14 May 2020).
- Maquart, M.; Dahmani, M.; Marie, J.L.; Gravier, P.; Leparc-Goffart, I.; Davoust, B. First serological evidence of West Nile virus in horses and dogs from Corsica Island, France. Vector Borne Zoonotic Dis. 2017, 17, 275–277. [Google Scholar] [CrossRef]
- Magurano, F.; Remoli, M.E.; Baggieri, M.; Fortuna, C.; Marchi, A.; Fiorentini, C.; Bucci, P.; Benedetti, E.; Ciufolini, M.G.; Rizzo, C.; et al. Circulation of West Nile virus lineage 1 and 2 during an outbreak in Italy. Clin. Microbiol. Infect. 2012, 18, E545–E547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzoli, A.; Jimenez-Clavero, M.A.; Barzon, L.; Cordioli, P.; Figuerola, J.; Koraka, P.; Martina, B.; Moreno, A.; Nowotny, N.; Pardigon, N.; et al. The challenge of West Nile virus in Europe: Knowledge gaps and research priorities. Eurosurveillance 2015, 20, 21135. [Google Scholar] [CrossRef] [Green Version]
- Beck, C.; Gonzalez, G.; Decors, A.; Lemberger, K.; Lowenski, S.; Dumarest, D.; Lecollinet, S. Surveillance épidémiologique du virus Usutu dans l’avifaune. Virologie 2018, 22, 261–263. [Google Scholar]
- Semenza, J.C.; Tran, A.; Espinosa, L.; Sudre, B.; Domanovic, D.; Paz, S. Climate change projections of West Nile virus infections in Europe: Implications for blood safety practices. Environ. Health 2016, 15, 28. [Google Scholar] [CrossRef] [Green Version]
- Chevalier, V.; Tran, A.; Durand, B. Predictive modeling of West Nile virus transmission risk in the Mediterranean basin: How far from landing? Int. J. Environ. Res. Public Health 2013, 11, 67–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beck, C.; Despres, P.; Paulous, S.; Vanhomwegen, J.; Lowenski, S.; Nowotny, N.; Durand, B.; Garnier, A.; Blaise-Boisseau, S.; Guitton, E.; et al. A high-performance multiplex immunoassay for serodiagnosis of flavivirus-associated neurological diseases in horses. BioMed Res. Int. 2015, 2015, 678084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carteaux, G.; Maquart, M.; Bedet, A.; Contou, D.; Brugières, P.; Fourati, S.; de Langavant, L.C.; de Broucker, T.; Brun-Buisson, C.; Leparc-Goffart, I.; et al. Zika virus associated with meningoencephalitis. New Engl. J. Med. 2016, 374, 1595–1596. [Google Scholar] [CrossRef] [PubMed]
- Linke, S.; Ellerbrok, H.; Niedrig, M.; Nitsche, A.; Pauli, G. Detection of West Nile virus lineages 1 and 2 by real-time PCR. J. Virol. Methods 2007, 146, 355–358. [Google Scholar] [CrossRef]
- OIE. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. 2018. Available online: https://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/3.01.24_WEST_NILE.pdf (accessed on 14 May 2020).
- Piorkowski, G.; Richard, P.; Baronti, C.; Gallian, P.; Charrel, R.; Leparc-Goffart, I.; de Lamballerie, X. Complete coding sequence of Zika virus from Martinique outbreak in 2015. New Microbes New Infect. 2016, 11, 52–53. [Google Scholar] [CrossRef] [Green Version]
- Sailleau, C.; Breard, E.; Viarouge, C.; Gorlier, A.; Quenault, H.; Hirchaud, E.; Touzain, F.; Blanchard, Y.; Vitour, D.; Zientara, S. Complete genome sequence of bluetongue virus serotype 4 that emerged on the French island of Corsica in December 2016. Transbound. Emerg. Dis. 2018, 65, e194–e197. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R-project.org (accessed on 14 May 2020).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Value | (a) Cx. pipiens Trapped in June | (b) Total Cx. pipiens Trapped | ||
|---|---|---|---|---|---|
| Abundance Ratio | p-Value | Coefficient | p-Value | ||
| Department | Bouches du Rhône | Reference | Reference | ||
| Gard | 1.14 | 0.53 | 1.02 | 0.92 | |
| Hérault | 0.36 | <0.0001 | 0.30 | <0.0001 | |
| Year | 2018 | Reference | Reference | ||
| 2015 | 0.20 | <0.0001 | 0.66 | 0.054 | |
| 2016 | 0.40 | 0.0007 | 0.51 | 0.002 | |
| 2017 | 0.30 | <0.0001 | 0.40 | <0.0001 | |
| 2019 | 0.24 | <0.0001 | 0.43 | <0.0001 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beck, C.; Leparc Goffart, I.; Franke, F.; Gonzalez, G.; Dumarest, M.; Lowenski, S.; Blanchard, Y.; Lucas, P.; Lamballerie, X.d.; Grard, G.; et al. Contrasted Epidemiological Patterns of West Nile Virus Lineages 1 and 2 Infections in France from 2015 to 2019. Pathogens 2020, 9, 908. https://doi.org/10.3390/pathogens9110908
Beck C, Leparc Goffart I, Franke F, Gonzalez G, Dumarest M, Lowenski S, Blanchard Y, Lucas P, Lamballerie Xd, Grard G, et al. Contrasted Epidemiological Patterns of West Nile Virus Lineages 1 and 2 Infections in France from 2015 to 2019. Pathogens. 2020; 9(11):908. https://doi.org/10.3390/pathogens9110908
Chicago/Turabian StyleBeck, Cécile, Isabelle Leparc Goffart, Florian Franke, Gaelle Gonzalez, Marine Dumarest, Steeve Lowenski, Yannick Blanchard, Pierrick Lucas, Xavier de Lamballerie, Gilda Grard, and et al. 2020. "Contrasted Epidemiological Patterns of West Nile Virus Lineages 1 and 2 Infections in France from 2015 to 2019" Pathogens 9, no. 11: 908. https://doi.org/10.3390/pathogens9110908