A Predicted Mannoprotein Cmp1 Regulates Fungal Virulence in Cryptococcus neoformans



Abstract
:1. Introduction
2. Results
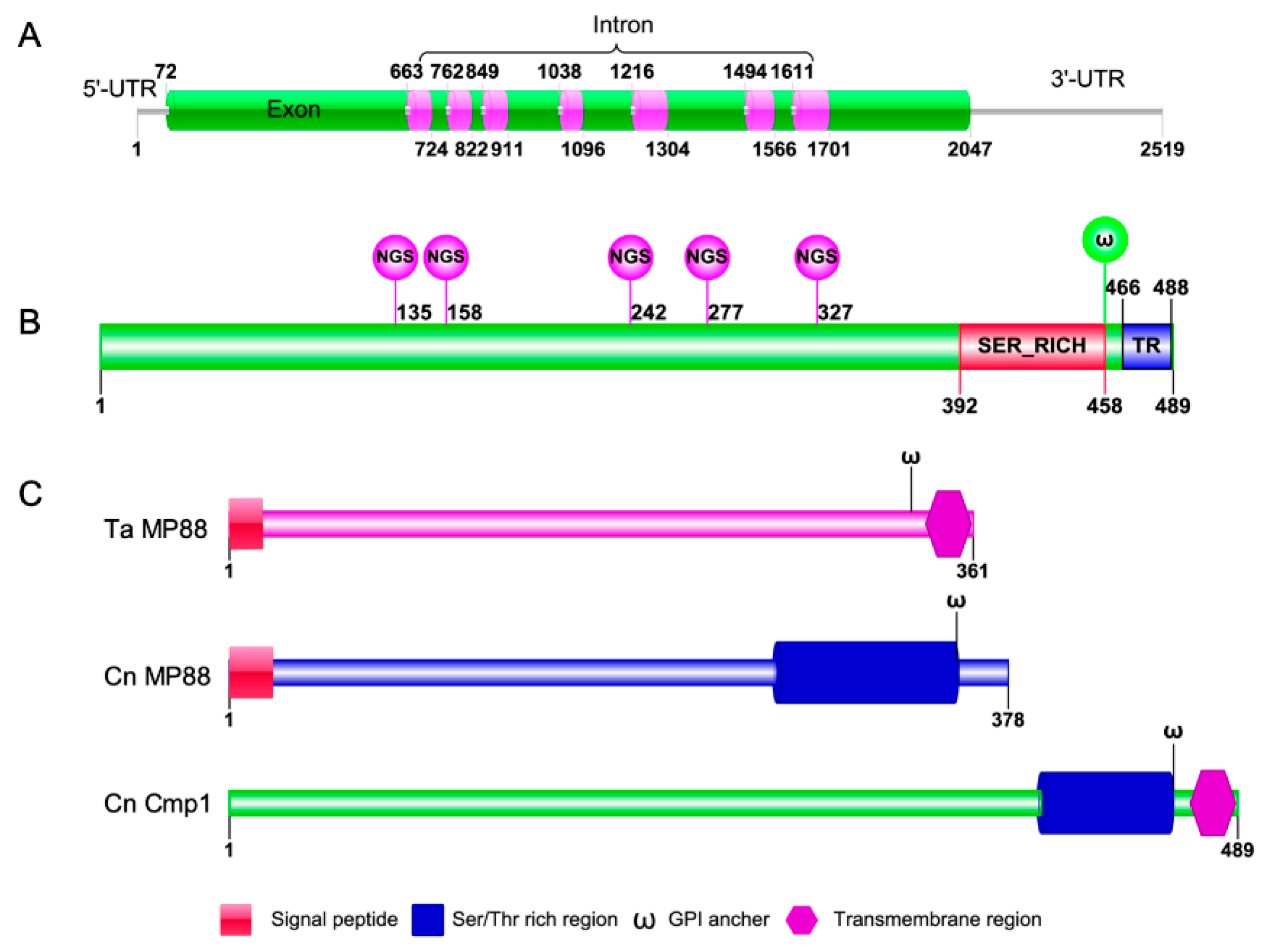
2.1. Identification of the Mannoprotein Cmp1 in C. neoformans
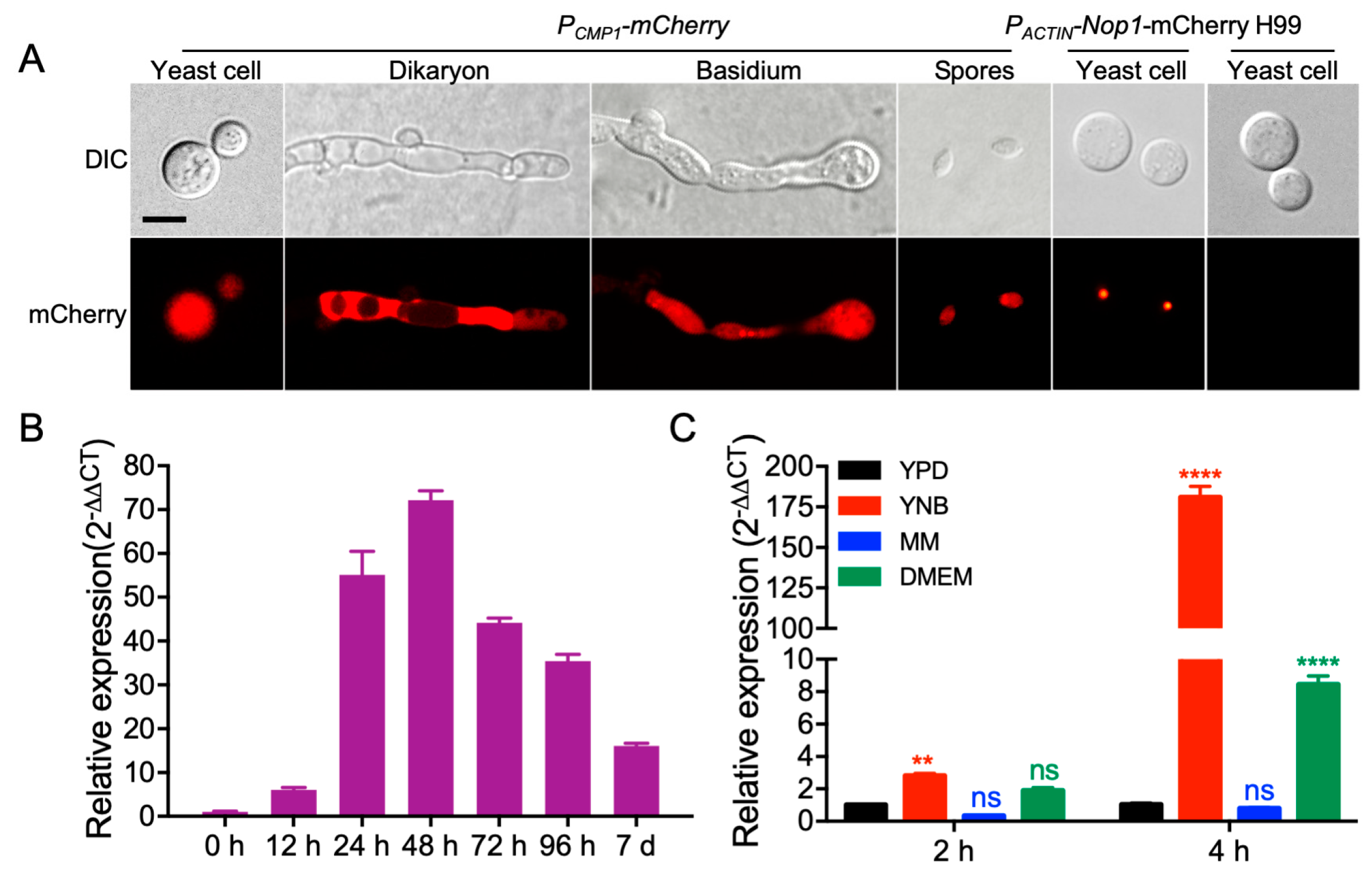
2.2. CMP1 Expression Pattern Analyses
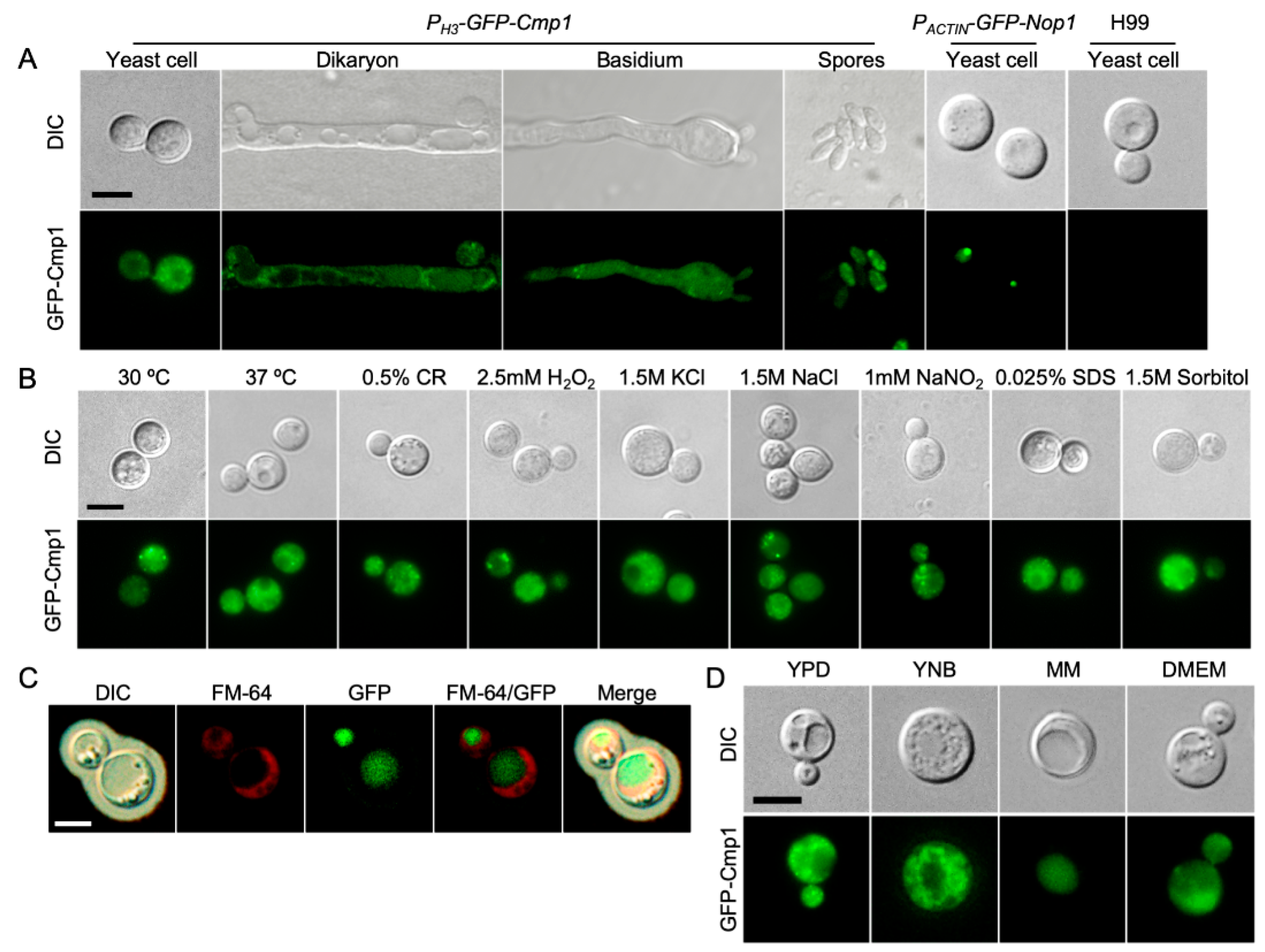
2.3. Cmp1 Localization in C. neoformans
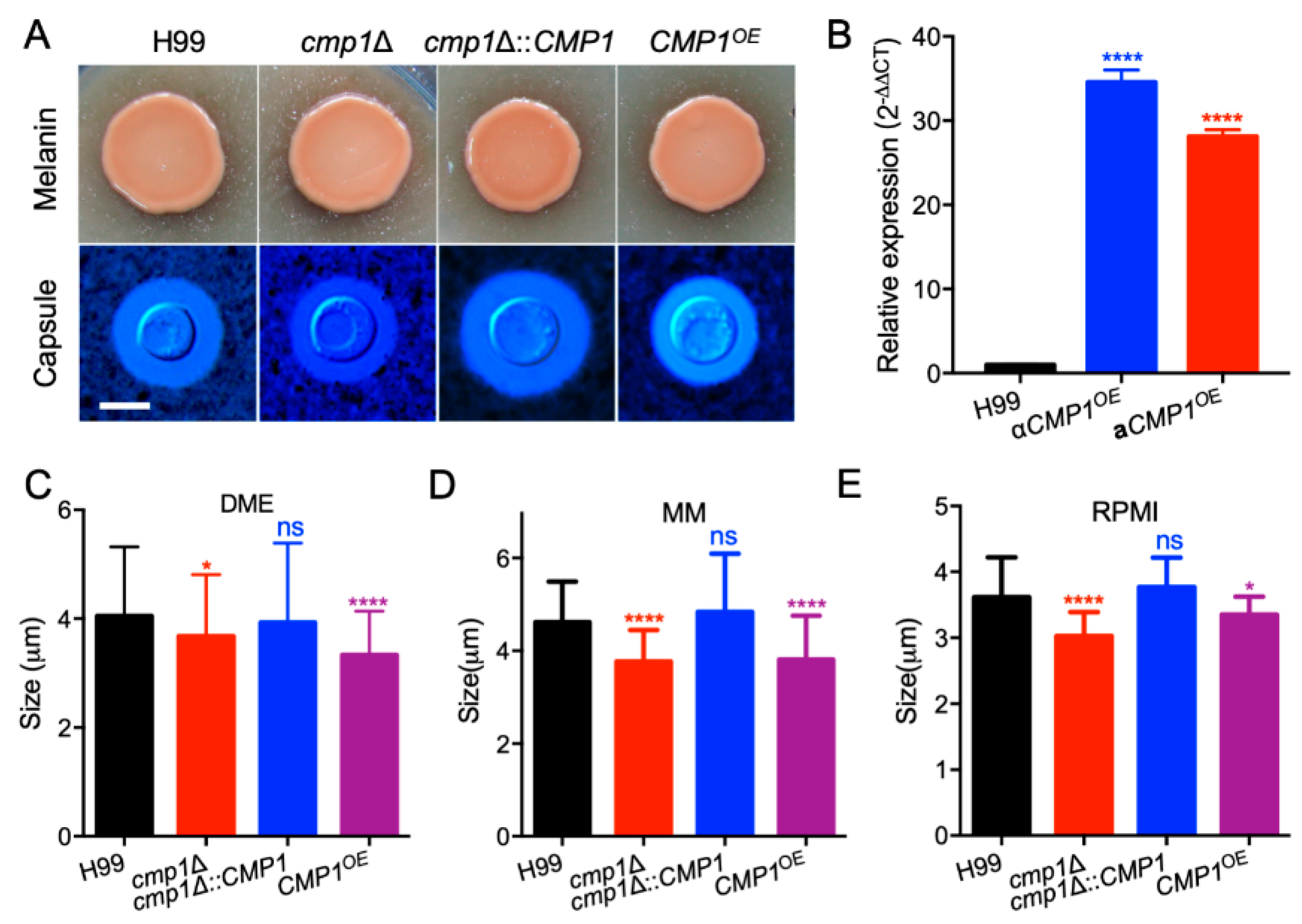
2.4. Cmp1 Regulates Capsule Formation
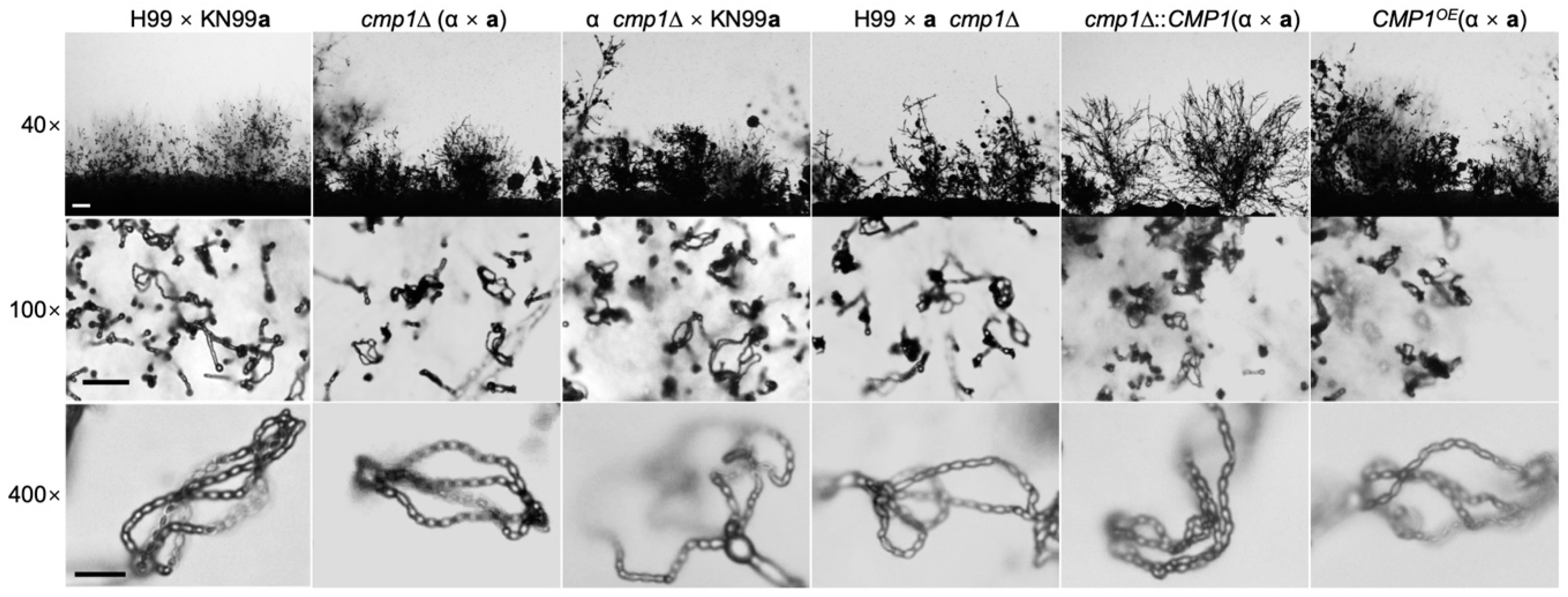
2.5. Cmp1 is not Required for Sexual Reproduction
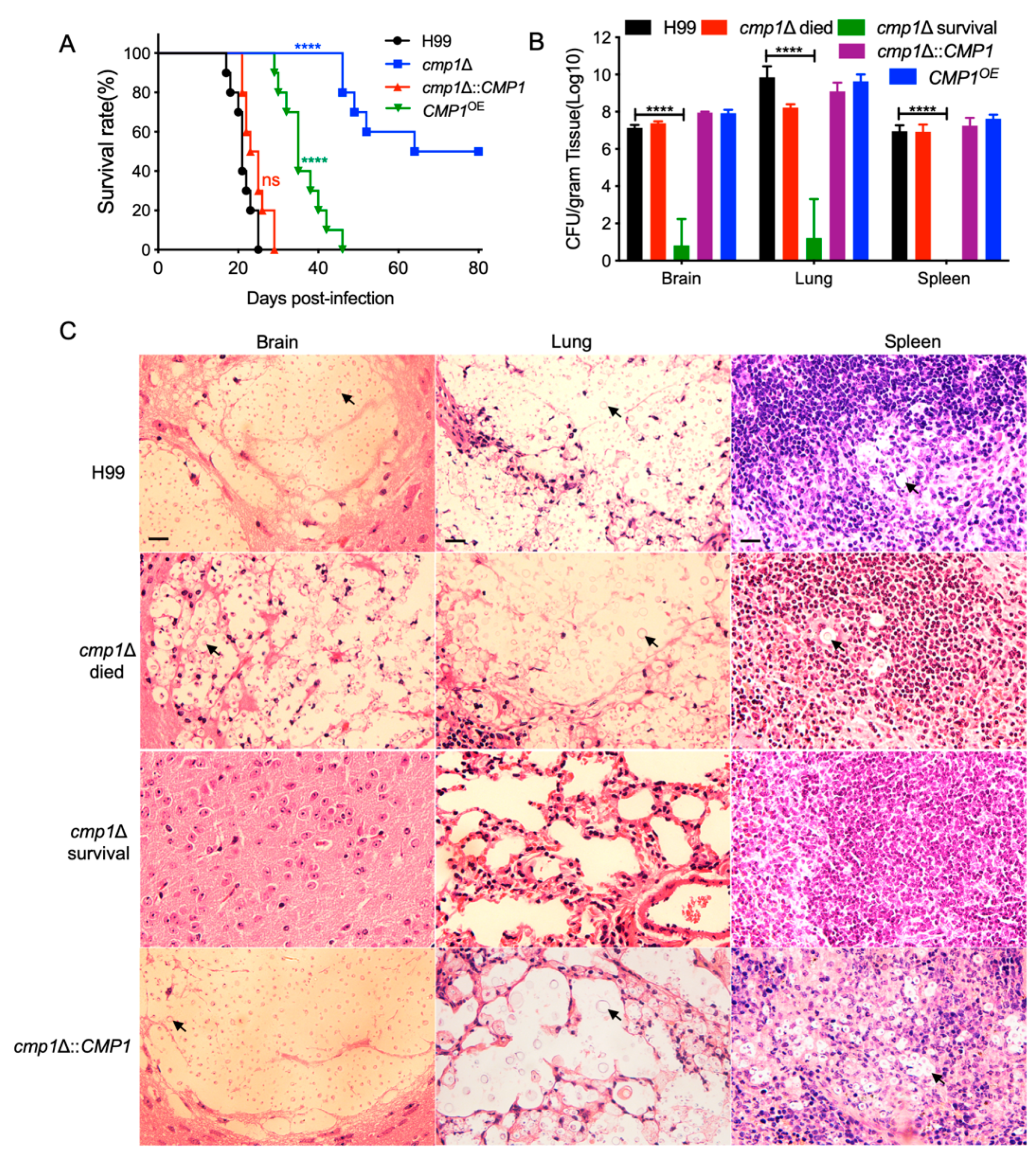
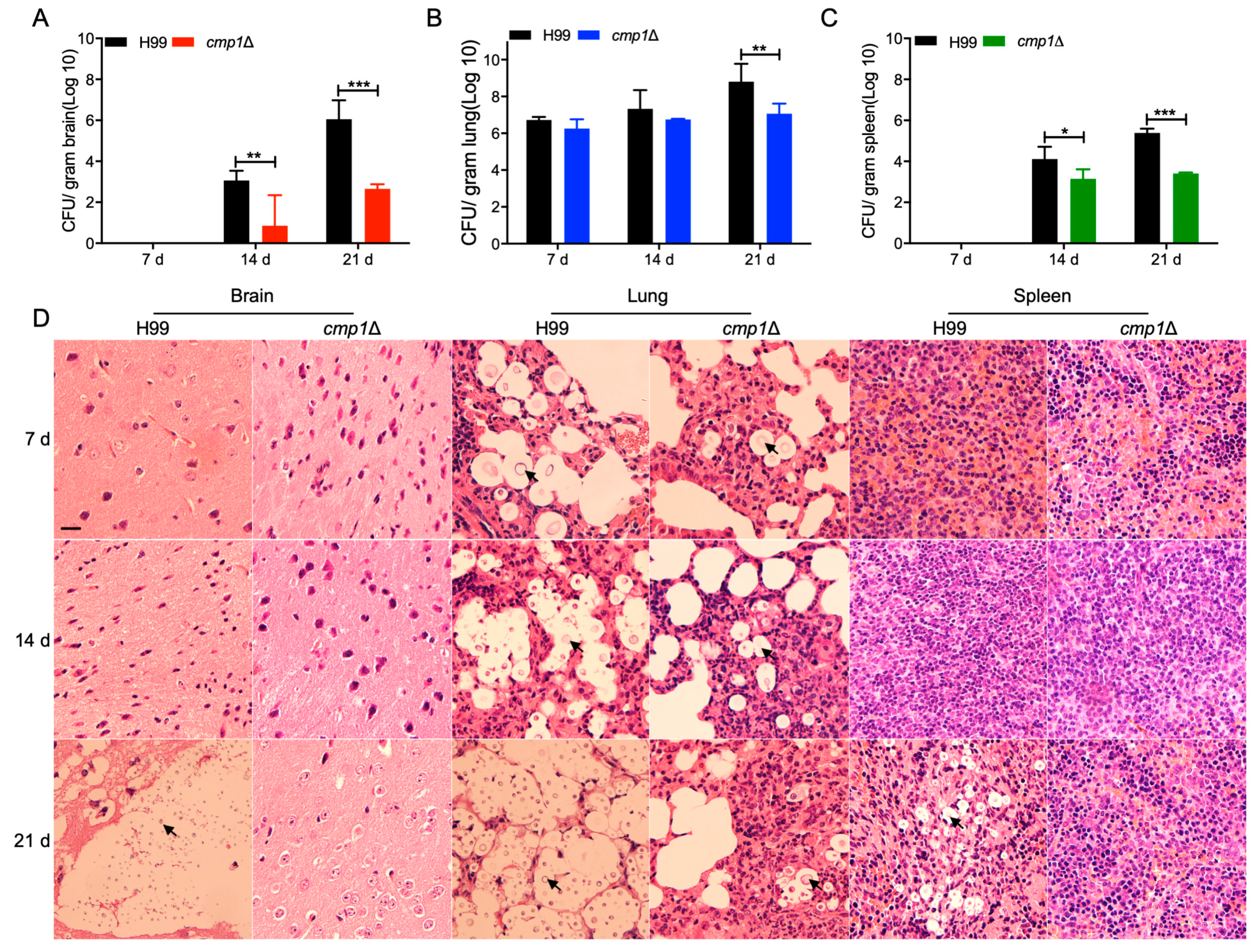
2.6. Cmp1 is Required for Fungal Infection
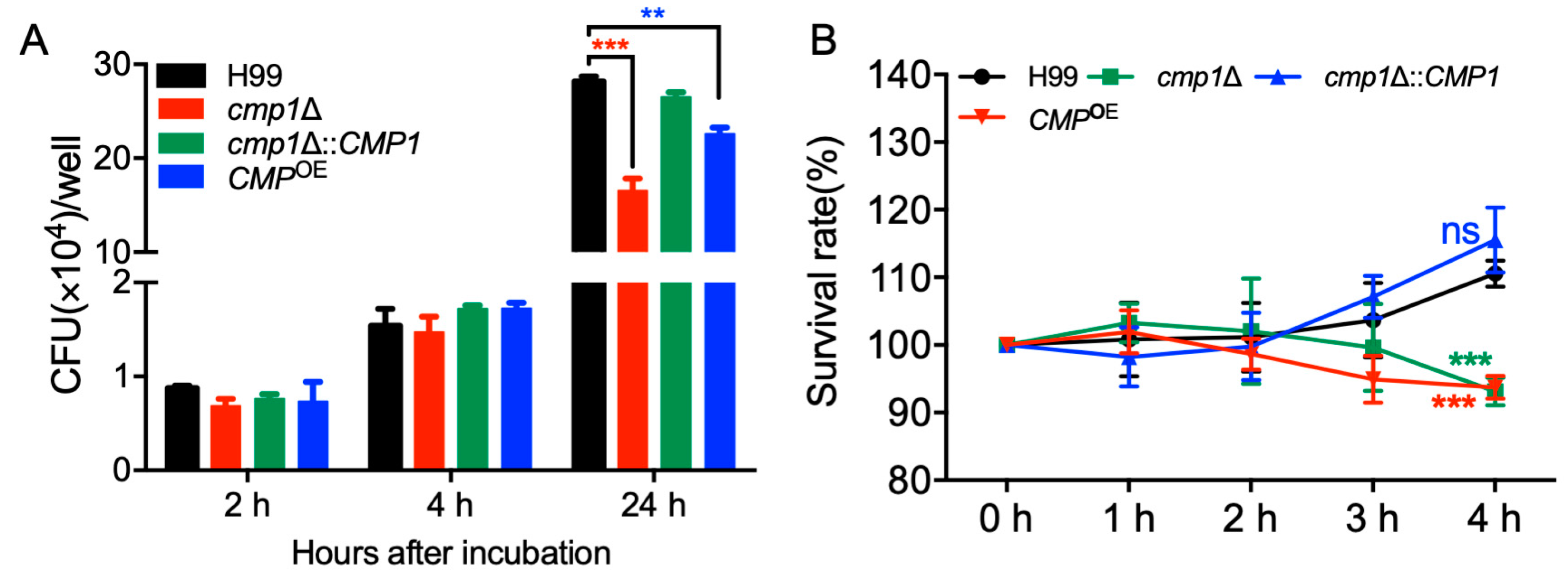
2.7. Cmp1 is Important for Proliferation Inside Macrophage and Survival in the Host Complement System
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Strains and Growth Conditions
4.3. CMP1 Expression Pattern Assay
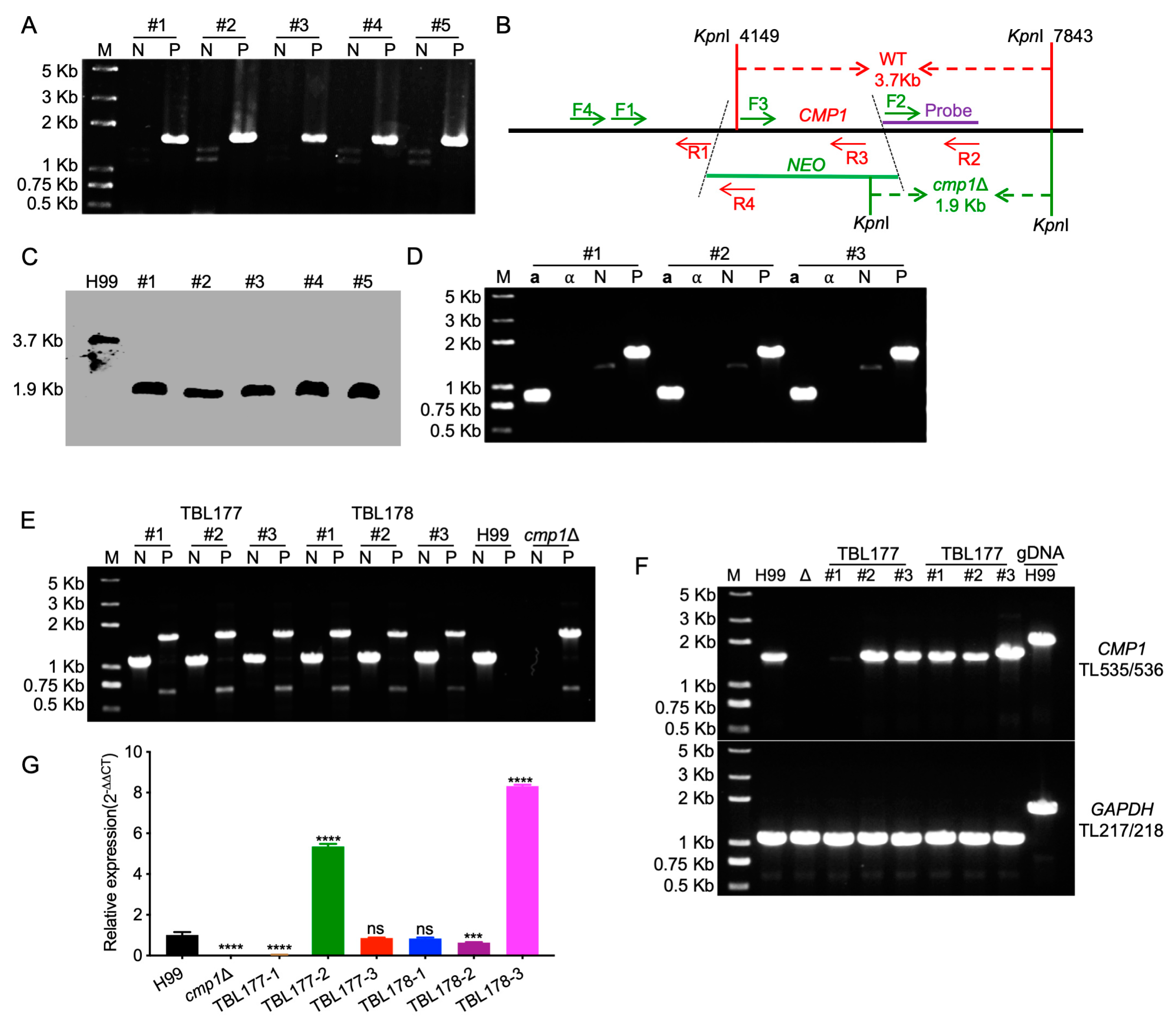
4.4. Generation of CMP1 Deletion, Complementation, and Overexpression Strains
4.5. Assays of Melanin and Capsule Production, and Mating
4.6. Virulence Studies
4.7. Fungal Loads and Histopathology of Infected Organs
4.8. Cryptococcus-Macrophage Interaction and Serum Treatment Assay
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Casadevall, A.; Perfect, J.R. Cryptococcus Neoformans; ASM Press: Washington, DC, USA, 1998. [Google Scholar]
- Park, B.J.; A Wannemuehler, K.; Marston, B.J.; Govender, N.; Pappas, P.G.; Chiller, T.M. Estimation of the current global burden of cryptococcal meningitis among persons living with HIV/AIDS. AIDS 2009, 23, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Rajasingham, R.; Smith, R.M.; Park, B.J.; Jarvis, J.N.; Govender, N.P.; Chiller, T.M.; Denning, D.W.; Loyse, A.; Boulware, D.R. Global burden of disease of HIV-associated cryptococcal meningitis: An updated analysis. Lancet Infect. Dis. 2017, 17, 873–881. [Google Scholar] [CrossRef] [Green Version]
- Kozel, T.R. Virulence factors of Cryptococcus neoformans. Trends Microbiol. 1995, 3, 295–299. [Google Scholar] [CrossRef]
- Kronstad, J.W.; Jung, W.H.; Hu, G. Beyond the Big Three: Systematic Analysis of Virulence Factors in Cryptococcus neoformans. Cell Host Microbe 2008, 4, 308–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaragoza, O. Basic principles of the virulence of Cryptococcus. Virulence 2019, 10, 490–501. [Google Scholar] [CrossRef] [Green Version]
- Doering, T.L. How Sweet it is! Cell Wall Biogenesis and Polysaccharide Capsule Formation inCryptococcus neoformans. Annu. Rev. Microbiol. 2009, 63, 223–247. [Google Scholar] [CrossRef] [Green Version]
- Taylor-Smith, L.M.; May, R.C. New weapons in the Cryptococcus infection toolkit. Curr. Opin. Microbiol. 2016, 34, 67–74. [Google Scholar] [CrossRef]
- De Jesus, M.; Nicola, A.M.; Chow, S.-K.; Lee, I.R.; Nong, S.; Specht, C.A.; Levitz, S.M.; Casadevall, A. Glucuronoxylomannan, galactoxylomannan, and mannoprotein occupy spatially separate and discrete regions in the capsule ofCryptococcus neoformans. Virulence 2010, 1, 500–508. [Google Scholar] [CrossRef]
- Biondo, C.; Messina, L.; Bombaci, M.; Mancuso, G.; Midiri, A.; Beninati, C.; Cusumano, V.; Gerace, E.; Papasergi, S.; Teti, G. Characterization of Two Novel Cryptococcal Mannoproteins Recognized by Immune Sera. Infect. Immun. 2005, 73, 7348–7355. [Google Scholar] [CrossRef] [Green Version]
- Eigenheer, R.A.; Lee, Y.J.; Blumwald, E.; Phinney, B.S.; Gelli, A. Extracellular glycosylphosphatidylinositol-anchored mannoproteins and proteases ofCryptococcus neoformans. FEMS Yeast Res. 2007, 7, 499–510. [Google Scholar] [CrossRef] [Green Version]
- Levitz, S.M.; Specht, C.A. The molecular basis for the immunogenicity ofCryptococcus neoformansmannoproteins. FEMS Yeast Res. 2006, 6, 513–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- E Vartivarian, S.; Reyes, G.H.; Jacobson, E.S.; James, P.G.; Cherniak, R.; Mumaw, V.R.; Tingler, M.J. Localization of mannoprotein in Cryptococcus neoformans. J. Bacteriol. 1989, 171, 6850–6852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, P.A.C.; Penha, L.L.; Mendonça -Previato, L.; Previato, J.L. Mannoprotein MP84 mediates the adhesion of Cryptococcus neoformans to epithelial lung cells. Front. Cell. Infect. Microbiol. 2014, 4, 106. [Google Scholar] [CrossRef] [Green Version]
- Mansour, M.K.; Yauch, L.E.; Rottman, J.B.; Levitz, S.M. Protective Efficacy of Antigenic Fractions in Mouse Models of Cryptococcosis. Infect. Immun. 2004, 72, 1746–1754. [Google Scholar] [CrossRef] [Green Version]
- Murphy, J.W. Influence of Cryptococcal Antigens on Cell-Mediated Immunity. Rev. Infect. Dis. 1988, 10 (Suppl. S2), S432–S435. [Google Scholar] [CrossRef]
- Murphy, J. Protective cell-mediated immunity against Cryptococcus neoformans. Res. Immunol. 1998, 149, 373–386. [Google Scholar] [CrossRef]
- Specht, C.A.; Nong, S.; Dan, J.M.; Lee, C.K.; Levitz, S.M. Contribution of Glycosylation to T Cell Responses Stimulated by RecombinantCryptococcus neoformansMannoprotein. J. Infect. Dis. 2007, 196, 796–800. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Nong, S.-H.; Mansour, M.K.; Specht, C.A.; Levitz, S.M. Purification and Characterization of a Second Immunoreactive Mannoprotein from Cryptococcus neoformans That Stimulates T-Cell Responses. Infect. Immun. 2002, 70, 5485–5493. [Google Scholar] [CrossRef] [Green Version]
- Mansour, M.K.; Schlesinger, L.S.; Levitz, S.M. Optimal T Cell Responses toCryptococcus neoformansMannoprotein Are Dependent on Recognition of Conjugated Carbohydrates by Mannose Receptors. J. Immunol. 2002, 168, 2872–2879. [Google Scholar] [CrossRef] [Green Version]
- Cadieux, B.; Lian, T.; Hu, G.; Wang, J.; Biondo, C.; Teti, G.; Liu, V.; Murphy, M.E.P.; Creagh, A.L.; Kronstad, J.W. The Mannoprotein Cig1 Supports Iron Acquisition from Heme and Virulence in the Pathogenic Fungus Cryptococcus neoformans. J. Infect. Dis. 2013, 207, 1339–1347. [Google Scholar] [CrossRef] [Green Version]
- Reuwsaat, J.C.V.; Motta, H.; Garcia, A.W.A.; Vasconcelos, C.B.; Marques, B.M.; Oliveira, N.K.; Rodrigues, J.; Ferrareze, P.A.G.; Frases, S.; Lopes, W.; et al. A Predicted Mannoprotein Participates inCryptococcus gattiiCapsular Structure. mSphere 2018, 3, e00023-18. [Google Scholar] [CrossRef] [Green Version]
- Viudes, A.; Lazzell, A.; Kirkpatrick, W.R.; Patterson, T.F.; Perea, S.; Pemán, J.; Martinez, J.P.; Loópez-Ribot, J.L. The C-terminal antibody binding domain ofCandida albicansmp58 represents a protective epitope during candidiasis. FEMS Microbiol. Lett. 2004, 232, 133–138. [Google Scholar] [CrossRef] [Green Version]
- Inukai, T.; Nagi, M.; Morita, A.; Tanabe, K.; Aoyama, T.; Miyazaki, Y.; Bard, M.; Nakayama, H. The mannoprotein TIR3 (CAGL0C03872g) is required for sterol uptake in Candida glabrata. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2015, 1851, 141–151. [Google Scholar] [CrossRef]
- Cao, L.; Chan, C.-M.; Lee, C.; Wong, S.S.-Y.; Yuen, K.-Y. MP1 Encodes an Abundant and Highly Antigenic Cell Wall Mannoprotein in the Pathogenic Fungus Penicillium marneffei. Infect. Immun. 1998, 66, 966–973. [Google Scholar] [CrossRef] [Green Version]
- Woo, P.C.Y.; Lau, S.K.P.; Lau, C.C.Y.; Tung, E.T.K.; Chong, K.T.K.; Yang, F.; Zhang, H.; Lo, R.K.C.; Cai, J.-P.; Au-Yeung, R.K.H.; et al. Mp1p Is a Virulence Factor in Talaromyces (Penicillium) marneffei. PLoS Negl. Trop. Dis. 2016, 10, e0004907. [Google Scholar] [CrossRef] [Green Version]
- Basenko, E.Y.; Pulman, J.A.; Shanmugasundram, A.; Harb, O.S.; Crouch, K.; Starns, D.; Warrenfeltz, S.; Aurrecoechea, C.; Stoeckert, J.C.J.; Kissinger, J.C.; et al. FungiDB: An Integrated Bioinformatic Resource for Fungi and Oomycetes. J. Fungi (Basel) 2018, 4, 39. [Google Scholar] [CrossRef] [Green Version]
- Perfect, J.R.; Ketabchi, N.; Cox, G.M.; Ingram, C.W.; Beiser, C.L. Karyotyping of Cryptococcus neoformans as an epidemiological tool. J. Clin. Microbiol. 1993, 31, 3305–3309. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, K.; Cox, G.M.; Wang, P.; Toffaletti, D.L.; Perfect, J.R.; Heitman, J. Sexual Cycle of Cryptococcus neoformans var. grubii and Virulence of Congenic a and α Isolates. Infect. Immun. 2003, 71, 4831–4841. [Google Scholar] [CrossRef] [Green Version]
- Price, M.S.; Nichols, C.B.; Alspaugh, J.A. The Cryptococcus neoformans Rho-GDP Dissociation Inhibitor Mediates Intracellular Survival and Virulence. Infect. Immun. 2008, 76, 5729–5737. [Google Scholar] [CrossRef] [Green Version]
- Fan, C.-L.; Han, L.-T.; Jiang, S.-T.; Chang, A.-N.; Zhou, Z.-Y.; Liu, T.-B. The Cys2His2 zinc finger protein Zfp1 regulates sexual reproduction and virulence in Cryptococcus neoformans. Fungal Genet. Biol. 2019, 124, 59–72. [Google Scholar] [CrossRef]
- Lee, S.C.; Heitman, J. Function of Cryptococcus neoformans KAR7 (SEC66) in Karyogamy during Unisexual and Opposite-Sex Mating. Eukaryot. Cell 2012, 11, 783–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levitz, S.M.; Nong, S.-H.; Mansour, M.K.; Huang, C.; Specht, C.A. Molecular characterization of a mannoprotein with homology to chitin deacetylases that stimulates T cell responses to Cryptococcus neoformans. Proc. Natl. Acad. Sci. USA 2001, 98, 10422–10427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kainz, K.; Bauer, M.A.; Madeo, F.; Carmona-Gutierrez, D. Fungal infections in humans: The silent crisis. Microb. Cell 2020, 7, 143–145. [Google Scholar] [CrossRef] [PubMed]
- Kabir, V.; Maertens, J.A.; Kuypers, D. Fungal infections in solid organ transplantation: An update on diagnosis and treatment. Transplant. Rev. 2019, 33, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Hasim, S.; Coleman, J.L. Targeting the fungal cell wall: Current therapies and implications for development of alternative antifungal agents. Futur. Med. Chem. 2019, 11, 869–883. [Google Scholar] [CrossRef]
- A Wang, Z.; Li, L.X.; Doering, T.L. Unraveling synthesis of the cryptococcal cell wall and capsule. Glycobiol. 2018, 28, 719–730. [Google Scholar] [CrossRef]
- Xue, C.; Tada, Y.; Dong, X.; Heitman, J. The Human Fungal Pathogen Cryptococcus Can Complete Its Sexual Cycle during a Pathogenic Association with Plants. Cell Host Microbe 2007, 1, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.-B.; Wang, Y.; Stukes, S.; Chen, Q.; Casadevall, A.; Xue, C. The F-Box Protein Fbp1 Regulates Sexual Reproduction and Virulence in Cryptococcus neoformans. Eukaryot. Cell 2011, 10, 791–802. [Google Scholar] [CrossRef] [Green Version]
- Vij, R.; Cordero, R.J.; Casadevall, A. The Buoyancy ofCryptococcus neoformansIs Affected by Capsule Size. mSphere 2018, 3, e00534-18. [Google Scholar] [CrossRef] [Green Version]
- Blackstock, R.; Murphy, J.W. Secretion of the C3 component of complement by peritoneal cells cultured with encapsulated Cryptococcus neoformans. Infect. Immun. 1997, 65, 4114–4121. [Google Scholar] [CrossRef] [Green Version]
- Davidson, R.C.; Cruz, M.; Sia, R.A.; Allen, B.; Alspaugh, J.; Heitman, J. Gene Disruption by Biolistic Transformation in Serotype D Strains of Cryptococcus neoformans. Fungal Genet. Biol. 2000, 29, 38–48. [Google Scholar] [CrossRef]
- Kim, M.S.; Kim, S.-Y.; Yoon, J.K.; Lee, Y.-W.; Bahn, Y.-S. An efficient gene-disruption method in Cryptococcus neoformans by double-joint PCR with NAT-split markers. Biochem. Biophys. Res. Commun. 2009, 390, 983–988. [Google Scholar] [CrossRef]
- Fraser, J.A.; Subaran, R.L.; Nichols, C.B.; Heitman, J. Recapitulation of the Sexual Cycle of the Primary Fungal Pathogen Cryptococcus neoformans var. gattii: Implications for an Outbreak on Vancouver Island, Canada. Eukaryot. Cell 2003, 2, 1036–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, G.M.; Mukherjee, J.; Cole, G.T.; Casadevall, A.; Perfect, J.R. Urease as a Virulence Factor in Experimental Cryptococcosis. Infect. Immun. 2000, 68, 443–448. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.-B.; Xue, C. Fbp1-Mediated Ubiquitin-Proteasome Pathway Controls Cryptococcus neoformans Virulence by Regulating Fungal Intracellular Growth in Macrophages. Infect. Immun. 2013, 82, 557–568. [Google Scholar] [CrossRef] [Green Version]
- Kingsbury, J.M.; McCusker, J.H. Fungal Homoserine Kinase (thr1Δ) Mutants Are Attenuated in Virulence and Die Rapidly upon Threonine Starvation and Serum Incubation. Eukaryot. Cell 2010, 9, 729–737. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Description | Average fbp1Δ/H99 | PEST Domain |
|---|---|---|---|
| CNAG_00626 | Uncharacterized protein | 3.02656981 | 1 |
| CNAG_06000 | Glycoprotein | 1.87701854 | 1 |
| CNAG_06195 | Uncharacterized protein | 1.59340506 | 0 |
| CNAG_05395 | Rab guanyl-nucleotide exchange factor | 1.5884178 | 3 |
| CNAG_00700 | Purine nucleotide biosynthesis-related protein | 1.54435399 | 0 |
| CNAG_02455 | Choline transporter | 1.46895293 | 0 |
| CNAG_06871 | Uncharacterized protein | 1.45686983 | 1 |
| CNAG_02138 | DNA replication ATP-dependent helicase Dna2 | 1.44213261 | 2 |
| CNAG_01536 | Nonmuscle myosin heavy chain b | 1.3779496 | 1 |
| CNAG_04056 | Rhomboid-like protein | 1.37338159 | 0 |
| CNAG_04669 | Mitochondrial matrix protein import protein | 1.36855445 | 0 |
| CNAG_03099 | Chitin synthase 1 | 1.36597408 | 1 |
| CNAG_01193 | Uncharacterized protein | 1.36434725 | 1 |
| CNAG_05817 | GDP-mannose transporter 1 | 1.35714493 | 0 |
| CNAG_05967 | Uncharacterized protein | 1.35536384 | 1 |
| CNAG_04327 | SCP160 protein | 1.35312155 | 1 |
| CNAG_00634 | Uncharacterized protein | 1.3519146 | 1 |
| CNAG_05173 | DNA-3-methyladenine glycosylase II | 1.34218534 | 0 |
| CNAG_03281 | Glycine-rich RNA binding protein, variant 2 | 1.33612571 | 0 |
| Strains/Plasmids | Genotypes/Properties | Sources/References |
|---|---|---|
| C. neoformans | ||
| H99 | MATα | Perfect et al., 1993 [28] |
| KN99a | MATa | Nielsen et al., 2003 [29] |
| TBL106 | MATα cmp1Δ::NEO | In this study |
| TBL137 | MATa cmp1Δ::NEO | In this study |
| TBL175 | MATa PCMP1-mCherry::NAT | In this study |
| TBL177 | MATα cmp1Δ::NEO CMP1::NAT | In this study |
| TBL178 | MATa cmp1Δ::NEO CMP1::NAT | In this study |
| TBL186 | MATα cmp1Δ::NEO PH3-GFP-CMP1::NAT | In this study |
| TBL187 | MATa cmp1Δ::NEO PH3-GFP-CMP1::NAT | In this study |
| TBL209 | MATα PCMP1-mCherry::NAT | In this study |
| Plasmids | ||
| pCN19 | Ampr Vector carrying GFP under the control of histone H3 promoter | Price et al., 2008 [30] |
| pTBL1 | Ampr Vector carrying NAT marker | Fan et al., 2019 [31] |
| pTBL3 | Ampr Vector carrying mCherry-GPD1 terminator | Fan et al., 2019 [31] |
| pTBL82 | Ampr Vector carrying PCMP1-mCherry-NAT for temporal expression assay | In this study |
| pTBL92 | Ampr Vector carrying PH3-GFP-CMP1 gene fusion for Cmp1 localization | In this study |
| pTBL94 | Ampr Vector carrying PCMP1-CMP1-NAT for CMP1 complementation | In this study |
| Primers | Targeted Genes | Sequence (5′-3′) |
|---|---|---|
| TL17 | M13F | GTAAAACGACGGCCAG |
| TL18 | M13R | CAGGAAACAGCTATGAC |
| TL19 | NEO split F | GGGCGCCCGGTTCTTTTTGTCA |
| TL20 | NEO split R | TTGGTGGTCGAATGGGCAGGTAGC |
| TL59 | NEO R4 | TGTGGATGCTGGCGGAGGATA |
| TL67 | STE20A ⍺ F | CCAAAAGCTGATGCTGTGGA |
| TL68 | STE20A a R | AGGACATCTATAGCAGAT |
| TL69 | STE20A a F | TCCACTGGCAACCCTGCGAG |
| TL70 | STE20A a R | ATCAGAGACAGAGGAGCAAGAC |
| TL217 | GAPDH qRT-PCR F | TGAGAAGGACCCTGCCAACA |
| TL218 | GAPDH qRT-PCR R | ACTCCGGCTTGTAGGCATCAA |
| TL235 | CMP1 KO F1 | GGGGTAAAAGAGGGAGGATGAGAC |
| TL236 | CMP1 KO R1 | CTGGCCGTCGTTTTACAGAACACCCGCCGCTGAACTTT |
| TL237 | CMP1 KO F2 | GTCATAGCTGTTTCCTGCTCCGCTGCAACCAAGGCTACCA |
| TL238 | CMP1 KO R2 | TGCGCGGCTCGAGACACAAGA |
| TL239 | CMP1 KO F3 | TCCTCCGACTCGCGCCTCATCAG |
| TL240 | CMP1 KO R3 | AGCTATCGCCGGCCCATTACCATC |
| TL241 | CMP1 KO F4 | GCCCACGCGCCCACATACAT |
| TL481 | CMP1 PRO F1 | ACGGTATCGATAAGCTTGACGGGTGCGGACGACATTTAGATTT (HindIII) |
| TL482 | CMP1 PRO R1 | CTAGAACTAGTGGATCCACAGCATGCACACTCTCTGCAT (BamHI) |
| TL483 | CMP1-mCherry F1 | TTAGTAAACTCGCCCAACATGTCTGGATCCATGGCTGGCAGGTGGCCGCTGC (BamHI) |
| TL484 | CMP1-mCherry R1 | CTTGCTCACCATTCTAGAACTAGTGGATCCAAGCAAGACGGCAGCACCAAAC (BamHI) |
| TL535 | CMP1 QPCR F1 | TCCTGGTATCTCCACCTCTTC |
| TL536 | CMP1 QPCR R1 | CAAACAGCATGCCGACAAC |
| TL554 | GFP-CMP1 F | GACGAGCTGTAcGGATCCATGGCTGGCAGGTGGCCGCTGCAC (BamHI) |
| TL555 | GFP-CMP1 R | CTGGCGGCCGTTACTAGTTTAAAGCAAGACGGCAGCACCAAA (SpeI) |
| TL562 | CMP1 Comp F | GATATCGAATTCCTGCAGCCCGGGGGATCCGCCCACGCGCCCACATACATCCTCGC (BamHI) |
| TL563 | CMP1 Comp R | CGGTGGCGGCCGCTCTAGAACTAGTGGATCCTGCGCGGCTCGAGACACAAGAGTAGA (BamHI) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, L.-T.; Wu, L.; Liu, T.-B. A Predicted Mannoprotein Cmp1 Regulates Fungal Virulence in Cryptococcus neoformans. Pathogens 2020, 9, 881. https://doi.org/10.3390/pathogens9110881
Han L-T, Wu L, Liu T-B. A Predicted Mannoprotein Cmp1 Regulates Fungal Virulence in Cryptococcus neoformans. Pathogens. 2020; 9(11):881. https://doi.org/10.3390/pathogens9110881
Chicago/Turabian StyleHan, Lian-Tao, Lei Wu, and Tong-Bao Liu. 2020. "A Predicted Mannoprotein Cmp1 Regulates Fungal Virulence in Cryptococcus neoformans" Pathogens 9, no. 11: 881. https://doi.org/10.3390/pathogens9110881