Human Campylobacteriosis Cases Traceable to Chicken Meat—Evidence for Disseminated Outbreaks in Finland


Abstract
:1. Introduction
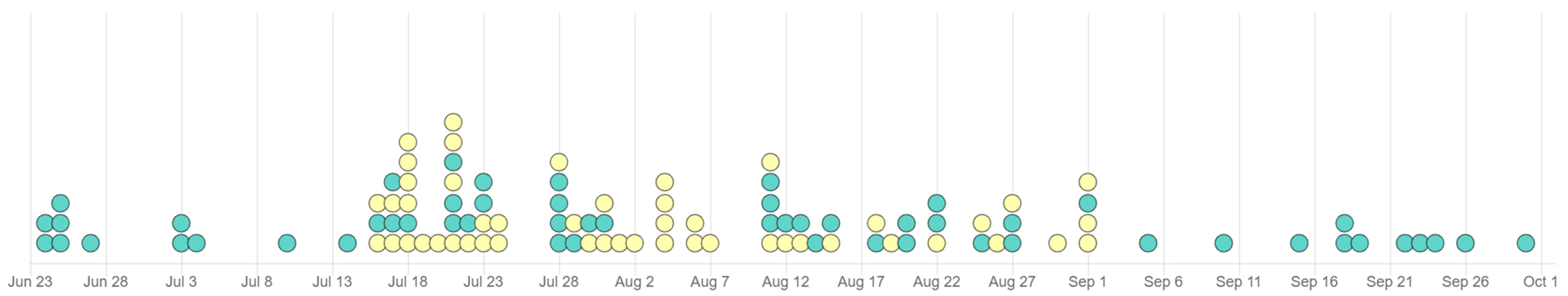
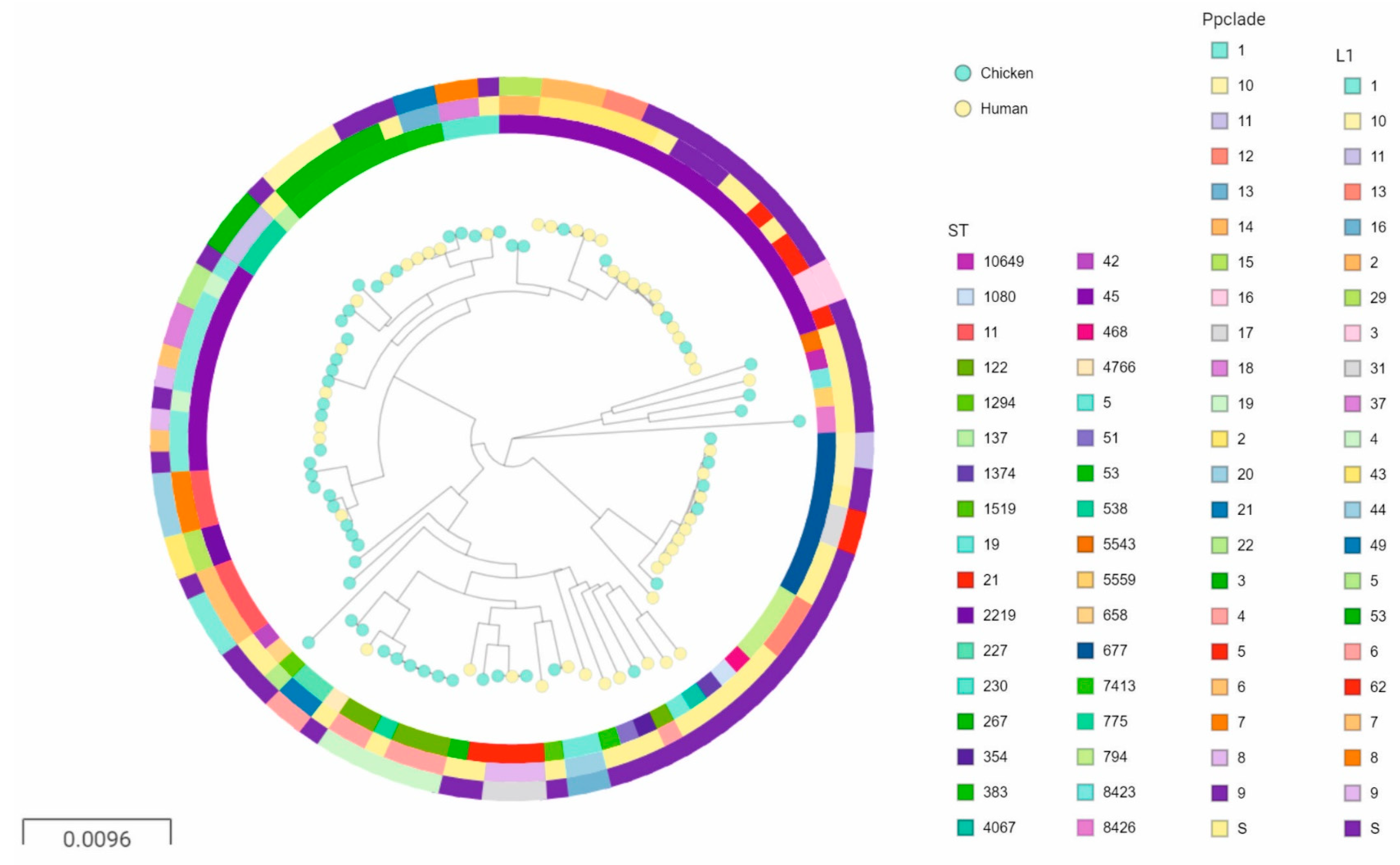
2. Results
2.1. Dataset
2.2. Identifying Clusters of Human and Chicken Isolates
3. Discussion
4. Materials and Methods
4.1. Selection of Isolates
4.2. DNA Extraction, WGS, Assembly and Multilocus Sequence Typing (MLST)
4.3. Analysis of the Clonal Relationship between the Human and Chicken C. jejuni Isolates
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Scarborough, P.; Appleby, P.N.; Mizdrak, A.; Briggs, A.D.M.; Travis, R.C.; Bradbury, K.E.; Key, T.J. Dietary greenhouse gas emissions of meat-eaters, fish-eaters, vegetarians and vegans in the UK. Clim. Chang. 2014, 125, 179–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suomen Siipikarjaliitto Ry. Siipikarjatuotanto Suomessa 1995–2019. Available online: http://www.siipi.net/index.php/siipikarjaliitto/tilastoa (accessed on 22 October 2020).
- EFSA Panel on Biological Hazards (BIOHAZ). Scientific Opinion on Quantification of the risk posed by broiler meat to human campylobacteriosis in the EU. EFSA J. 2010, 8, 1437. [Google Scholar] [CrossRef]
- Romero-Barrios, P.; Hempen, M.; Messens, W.; Stella, P.; Hugas, M. Quantitative microbiological risk assessment (QMRA) of food-borne zoonoses at the European level. Food Control 2013, 29, 343–349. [Google Scholar] [CrossRef]
- WHO (World Health Organization); OIE (World Organisation for Animal Health). The global view of campylobacteriosis: Report of an expert consultation. In WHO Report; WHO: Utrecht, The Netherlands, 2013. [Google Scholar]
- Maa-Ja Metsätalousministeriö. Maa-Ja Metsätalousministeriön Asetusbroilereiden Kampylobakteerivalvonnasta; EEO: Maa-Ja Metsätalousministeriö, Finland, 2007. [Google Scholar]
- Gonzalez, M.; Mikkelä, A.; Tuominen, P.; Ranta, J.; Hakkinen, M.; Hänninen, M.-L.; Llarena, A.-K. Risk Assessment of Campylobacter spp. in Finland. In Evira Research Report; Finnish Food Safety Authority: Evira, Finland, 2016. [Google Scholar]
- Jaakola, S.L.; Rimhanen-Finne, R.; Salmenlinna, S.; Savolainen-Kopra, C.; Liitsola, K.; Jalava, J.; Toropainen, M.; Nohynek, H.; Virtanen, M.; Löflund, J.-E.; et al. Infectious Diseases in Finland 2017; Nationl Institute for Health and Welfare (THL): Helsinki, Finland, 2018. [Google Scholar]
- Rosef, O.; Paulauskas, A.; Grude, N.; Haslekås, C.; Jenkins, A. Comparison of Norwegian poultry, waterborne and clinical isolates of Campylobacter jejuni by ribotyping. J. Bacteriol. 2009, 1, 51–57. [Google Scholar]
- Kapperud, G.; Espeland, G.; Wahl, E.; Walde, A.; Herikstad, H.; Gustavsen, S.; Digranes, A. Factors Associated with Increased and Decreased Risk of Campylobacter Infection: A Prospective Case-Control Study in Norway. Am. J. Epidemiol. 2003, 158, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Schönberg-Norio, D.; Takkinen, J.; Hänninen, M.-L.; Katila, M.-L.; Kaukoranta, S.-S.; Mattila, L.; Rautelin, H. Swimming and Campylobacter infections. Emerg. Infect. Dis. 2004, 10, 1474–1477. [Google Scholar] [CrossRef] [PubMed]
- Kovanen, S.; Kivistö, R.; Llarena, A.K.; Zhang, J.; Kärkkäinen, U.M.; Tuuminen, T.; Uksila, J.; Hakkinen, M.; Rossi, M.; Hänninen, M.L. Tracing isolates from domestic human Campylobacter jejuni infections to chicken slaughter batches and swimming water using whole-genome multilocus sequence typing. Int. J. Food Microbiol. 2016, 226, 53–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castrodale, L.J.; Provo, G.M.; Xavier, C.M.; McLaughlin, J.B. Calling all Campy—how routine investigation and molecular characterization impacts the understanding of campylobacteriosis epidemiology—Alaska, United States, 2004–2013. Epidemiol. Infect. 2016, 144, 265–267. [Google Scholar] [CrossRef]
- Strachan, N.F.; Forbes, K.J. Extensive Spatial and Temporal Clustering of Campylobacter Infections Evident in High-Resolution Genotypes. In Campylobacter Ecology and Evolution; Sheppard, S., Ed.; Caister Academic Press: Norfolk, UK, 2014; Volume 1. [Google Scholar]
- Joensen, K.G.; Kuhn, K.G.; Muller, L.; Bjorkman, J.T.; Torpdahl, M.; Engberg, J.; Holt, H.M.; Nielsen, H.L.; Petersen, A.M.; Ethelberg, S.; et al. Whole-genome sequencing of Campylobacter jejuni isolated from Danish routine human stool samples reveals surprising degree of clustering. Clin. Microbiol. Infect. 2018, 24, 201.e205–201.e208. [Google Scholar] [CrossRef]
- Llarena, A.-K.; Ribeiro-Gonçalves, B.F.; Nuno Silva, D.; Halkilahti, J.; Machado, M.P.; Da Silva, M.S.; Jaakkonen, A.; Isidro, J.; Hämäläinen, C.; Joenperä, J.; et al. INNUENDO: A cross-sectoral platform for the integration of genomics in the surveillance of food-borne pathogens. EFSA Supporting Publ. 2018, 15, 1498E. [Google Scholar] [CrossRef]
- Lees, J.A.; Harris, S.R.; Tonkin-Hill, G.; Gladstone, R.A.; Lo, S.W.; Weiser, J.N.; Corander, J.; Bentley, S.D.; Croucher, N.J. Fast and flexible bacterial genomic epidemiology with PopPUNK. Genome Res. 2019, 29, 304–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, M.; Da Silva, M.S.; Ribeiro-Gonçalves, B.F.; Silva, D.N.; Machado, M.P.; Oleastro, M.; Borges, V.; Isidro, J.; Viera, L.; Barker, D.; et al. NNUENDO whole genome and core genome MLST schemas and datasets for Campylobacter jejuni (Version 1.0) [Data set]. Zenodo 2018. [Google Scholar] [CrossRef]
- Argimón, S.; Abudahab, K.; Goater, R.J.E.; Fedosejev, A.; Bhai, J.; Glasner, C.; Feil, E.J.; Holden, M.T.G.; Yeats, C.A.; Grundmann, H.; et al. Microreact: Visualizing and sharing data for genomic epidemiology and phylogeography. Microb. Genome 2016, 2. [Google Scholar] [CrossRef] [Green Version]
- Dingle, K.E.; Colles, F.M.; Wareing, D.R.A.; Ure, R.; Fox, A.J.; Bolton, F.E.; Bootsma, H.J.; Willems, R.J.L.; Urwin, R.; Maiden, M.C.J. Multilocus Sequence Typing System forCampylobacter jejuni. J. Clin. Microbiol. 2001, 39, 14–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, M.; Machado, M.P.; Silva, D.N.; Rossi, M.; Moran-Gilad, J.; Santos, S.; Ramirez, M.; Carriço, J.A. chewBBACA: A complete suite for gene-by-gene schema creation and strain identification. Microb. Genome 2018, 4. [Google Scholar] [CrossRef]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef]
- Neimann, J.; Engberg, J.; Molbak, K.; Wegener, H.C. A case-control study of risk factors for sporadic campylobacter infections in Denmark. Epidemiol. Infect. 2003, 130, 353–366. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, E.; White, R.; Mexia, R.; Bruun, T.; Kapperud, G.; Lange, H.; Nygard, K.; Vold, L. Risk Factors for Sporadic Domestically Acquired Campylobacter Infections in Norway 2010-2011: A National Prospective Case-Control Study. PLoS ONE 2015, 10, e0139636. [Google Scholar] [CrossRef] [Green Version]
- Llarena, A.-K.; Huneau, A.; Hakkinen, M.; Hänninen, M.-L. Predominant Campylobacter jejuni sequence types persist in Finnish chicken production. PLoS ONE 2015, 10, e0116585. [Google Scholar] [CrossRef]
- De Haan, C.P.; Kivistö, R.I.; Hakkinen, M.; Corander, J.; Hänninen, M.-L.J.B.M. Multilocus sequence types of Finnish bovine Campylobacter jejuni isolates and their attribution to human infections. BMC Microbiol. 2010, 10, 200. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.; Messam, L.L.M.; Meade, J.; Gibbons, J.; McGill, K.; Bolton, D.; Whyte, P. The impact of biosecurity and partial depopulation on Campylobacter prevalence in Irish broiler flocks with differing levels of hygiene and economic performance. Infect. Ecol. Epidemiol. 2016, 6, 31454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llarena, A.-K.; Zhang, J.; Vehkala, M.; Välimäki, N.; Hakkinen, M.; Hänninen, M.-L.; Roasto, M.; Mäesaar, M.; Taboada, E.; Barker, D.; et al. Monomorphic genotypes within a generalist lineage of Campylobacter jejuni show signs of global dispersion. Microb. Genome 2016, 2, e000088. [Google Scholar] [CrossRef] [PubMed]
- Lihatiedotusyhdistys, Ry. Lihatuotanto Suomessa. Available online: https://www.lihatiedotus.fi/tilastotietoa/lihantuotanto-suomessa-2.html (accessed on 22 October 2020).
- Elvers, K.T.; Morris, V.K.; Newell, D.G.; Allen, V.M. Molecular Tracking, through Processing, of Campylobacter Strains Colonizing Broiler Flocks. Appl. Environ. Microbiol. 2011, 77, 5722–5729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jonsson, M.E.; Chriél, M.; Norström, M.; Hofshagen, M. Effect of climate and farm environment on Campylobacter spp. colonisation in Norwegian broiler flocks. Prev. Vet. Med. 2012, 107, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Tam, C.C.; Rodrigues, L.C.; Viviani, L.; Dodds, J.P.; Evans, M.R.; Hunter, P.R.; Gray, J.J.; Letley, L.H.; Rait, G.; Tompkins, D.S.; et al. Longitudinal study of infectious intestinal disease in the UK (IID2 study): Incidence in the community and presenting to general practice. Gut 2012, 61, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, S.K.; Maiden, M.C.J. The Evolution of Campylobacter jejuni and Campylobacter coli. Cold Spring Harb. Perspect. Biol. 2015, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maiden, M.C.J.; van Rensburg, M.J.J.; Bray, J.E.; Earle, S.G.; Ford, S.A.; Jolley, K.A.; McCarthy, N.D. MLST revisited: The gene-by-gene approach to bacterial genomics. Nat. Rev. Microbiol. 2013, 11, 728–736. [Google Scholar] [CrossRef] [Green Version]
- Satakuntoliitto. Available online: http://www.satakunta.fi/ (accessed on 22 October 2020).
- THL. Tartuntatautirekisterin Tilastotietokanta. Available online: https://www.thl.fi/ttr/gen/rpt/tilastot.html (accessed on 15 May 2020).
- Suomen Broileryhdistys, Ry. Missä Broilerituotantoa on? Available online: http://suomibroileri.fi/fi/missa (accessed on 22 October 2020).
- Kärenlampi, R.; Rautelin, H.; Hakkinen, M.; Hänninen, M.L. Temporal and geographical distribution and overlap of Penner heat-stable serotypes and pulsed-field gel electrophoresis genotypes of Campylobacter jejuni isolates collected from humans and chickens in Finland during a seasonal peak. J. Clin. Microbiol. 2003, 41, 4870–4872. [Google Scholar] [CrossRef] [Green Version]
- Institute, N.F. Thermotolerant Campylobacter. Detection, Semi-Quantitative and Quantitative Determination in Foods and Drinking Water, 3rd ed.; Technical University of Denmark: Lyngby, Denmark, 2007. [Google Scholar]
- European Food Safety Authority and European Centre for Disease Prevention and Control (ECDC). The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2014. EFSA J. 2015, 13, 4329. [Google Scholar] [CrossRef]
- Machado, M.P.H.; Halkilahti, J.; Jaakkonen, A.; Silva, D.N.; Mendes, I.; Nalbantoglu, Y.; Borges, V.; Ramirez, M.; Rossi, M.; Carriço, J.A. INNUca GitHub. Available online: https://github.com/B-UMMI/INNUca (accessed on 7 October 2020).
- Francisco, A.P.; Bugalho, M.; Ramirez, M.; Carriço, J.A. Global optimal eBURST analysis of multilocus typing data using a graphic matroid approach. BMC Bioinform. 2009, 10, 152. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, M.; Sousa, A.; Ramirez, M.; Francisco, A.P.; Carriço, J.A.; Vaz, C. PHYLOViZ 2.0: Providing scalable data integration and visualization for multiple phylogenetic inference methods. Bioinformatics 2016, 33, 128–129. [Google Scholar] [CrossRef]
- Carriço, J.A.; Silva-Costa, C.; Melo-Cristino, J.; Pinto, F.R.; de Lencastre, H.; Almeida, J.S.; Ramirez, M. Illustration of a Common Framework for Relating Multiple Typing Methods by Application to Macrolide-Resistant Streptococcus pyogenes. J. Clin. Microbiol. 2006, 44, 2524–2532. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| ST | Human | Chicken | Human Cases Preceded by Chicken | Temporal Human Clusters | Total |
|---|---|---|---|---|---|
| 45 | 19 | 13 | 18 | All | 32 |
| 677 | 5 | 4 | 5 | 1 | 9 |
| 122 | 0 | 6 | NA | NA | 6 |
| 267 | 5 | 2 | 3 | 2 | 7 |
| 11 | 1 | 6 | 0 | 0 | 7 |
| 794 | 4 | 0 | 0 | 1 | 4 |
| 230 | 1 | 2 | 1 | NA | 3 |
| 538 | 1 | 2 | 0 | 0 | 3 |
| 21 | 1 | 3 | 1 | 0 | 4 |
| 19 | 1 | 1 | 1 | NA | 2 |
| 227 | 0 | 2 | NA | NA | 2 |
| 383 | 0 | 2 | NA | NA | 2 |
| 2219 | 0 | 2 | 0 | 0 | 2 |
| Other * | 11 | 10 | 0 | 0 | 21 |
| ST | Isolates n (Human/Chicken) | Allele Difference a | Collection Dates Human | Collection Dates Chicken | L1:L2:L3 |
|---|---|---|---|---|---|
| Chicken Before Human Clusters | |||||
| 19 | 1/1 | 0 | August 31 | July 17 | 16:13:8 |
| 21 | 1/2 | 0 | August 26 c | August 14 c and August 20 c | 31:25:3 |
| 45 | 2/1 | 0 | July 21 c and July 31 | July 17 c | 2:6:1 |
| 230 | 1/1 | 0 | August 4 | July 23 | 8:8:1 |
| 267 | 2/1 | 0 | August 11 and August 12 | July 28 | 10:4:1 |
| 538 b | 1/1 | 0 | July 18 | June 25 | 53:38:1 |
| 677 | 1/1 | 0 | July 19 c | July 4 c | 11:3:2 |
| 677 | 1/1 | 0 | August 13 | July 31 | 62:3:2 |
| Human Clusters | |||||
| 45 b | 2 | 3 | July 21 and July 23 | NA | 7:2:1 |
| 45 | 2 | 0 | July 22 and July 29 | NA | 13:6:1 |
| Humans Before/Simultaneous as Chicken | |||||
| 45 | 1/1 | 2 | July 18 | July 28 | 5:2:1 |
| 45 b | 1/1 | 3 | September 1 | September 19 | 3:1:1 |
| 267 | 2/1 | 0 | August 11 c and August 12 | August 12 c | 10:4:1 |
| 538 | 1/1 | 0 | July 18 | August 11 | 53:38:1 |
| Chicken Clusters | |||||
| 2219 | 2 | 0 | NA | July 18 and July 21 | 44:5:1 |
| 11 b | 3 | 2 | NA | July 3, July 21 and July 23 | 1:5:1 |
| 11 | 3 | 0 | NA | July 28 and August 11 | 44:31:1 |
| 45 | 2 | 0 | NA | August 18 and August 22 | 37:2:1 |
| 383 | 2 | 0 | NA | August 20 and August 22 | 49:34:1 |
| 122 and 755 | 6 | 0 | NA | September 10 and September 24 | 4:9:4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Llarena, A.-K.; Kivistö, R. Human Campylobacteriosis Cases Traceable to Chicken Meat—Evidence for Disseminated Outbreaks in Finland. Pathogens 2020, 9, 868. https://doi.org/10.3390/pathogens9110868
Llarena A-K, Kivistö R. Human Campylobacteriosis Cases Traceable to Chicken Meat—Evidence for Disseminated Outbreaks in Finland. Pathogens. 2020; 9(11):868. https://doi.org/10.3390/pathogens9110868
Chicago/Turabian StyleLlarena, Ann-Katrin, and Rauni Kivistö. 2020. "Human Campylobacteriosis Cases Traceable to Chicken Meat—Evidence for Disseminated Outbreaks in Finland" Pathogens 9, no. 11: 868. https://doi.org/10.3390/pathogens9110868