Molecular and Serological Prevalence of Leptospira spp. in Feral Pigs (Sus scrofa) and their Habitats in Alabama, USA


 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Sample Compositions
2.2. Molecular Detection of Leptospira Species
2.3. Detection of Anti-Leptospira IgG Antibody
3. Discussions
4. Materials and Methods
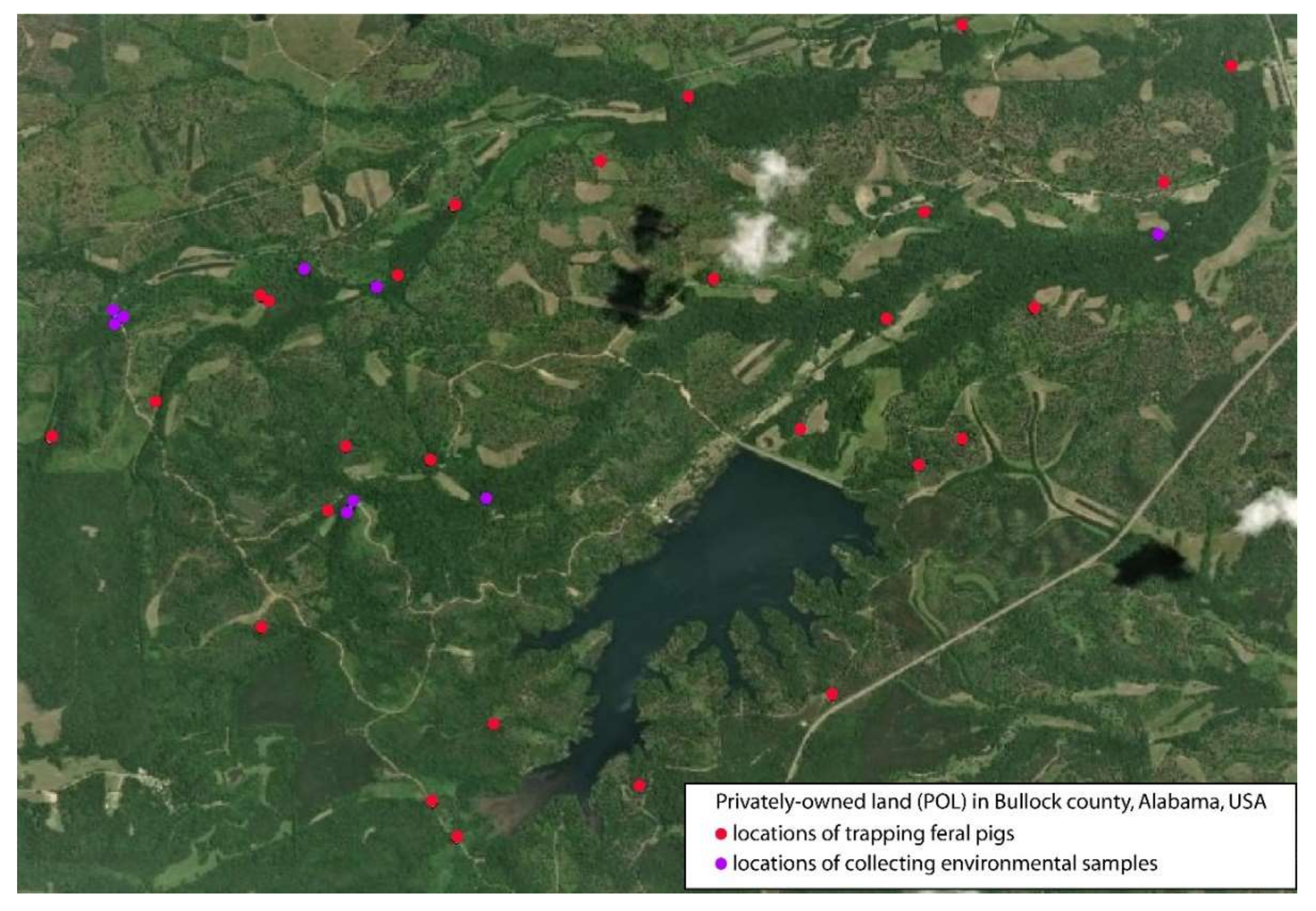
4.1. Study Areas
4.2. Trapping of Feral Pigs and Collection of Tissues
4.3. Collection of Environmental Samples
4.4. Extraction of Nucleic Acids
4.5. Molecular Detection of Leptospira spp. and DNA Sequencing
4.6. Anti-Leptospira spp. IgG ELISA
4.7. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Dupouey, J.; Faucher, B.; Edouard, S.; Richet, H.; Kodjo, A.; Drancourt, M.; Davoust, B. Human leptospirosis: An emerging risk in Europe? Comp. Immunol. Microbiol. Infect. Dis. 2014, 37, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Barragan, V.; Olivas, S.; Keim, P.; Pearson, T. Critical knowledge gaps in our understanding of environmental cycling and transmission of Leptospira spp. Appl. Environ. Microbiol. 2017, 83, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fentahun, T.; Alemayehu, M. Leptospirosis and its Public Health Significance: A Review. Eur. J. Appl. Sci. 2012, 4, 238–244. [Google Scholar] [CrossRef]
- Bharti, A.R.; Nally, J.E.; Ricaldi, J.N.; Matthias, M.A.; Diaz, M.M.; Lovett, M.A.; Levett, P.N.; Gilman, R.H.; Willig, M.R.; Gotuzzo, E.; et al. Leptospirosis: A zoonotic disease of global importance. Lancet Infect. Dis. 2003, 3, 757–771. [Google Scholar] [CrossRef]
- Cilia, G.; Bertelloni, F.; Angelini, M.; Cerri, D.; Fratini, F. Leptospira survey in Wild Boar (Sus scrofa) hunted in Tuscany, Central Italy. Pathogens 2020, 9, 377. [Google Scholar] [CrossRef]
- Ngugi, J.N.; Fèvre, E.M.; Mgode, G.F.; Obonyo, M.; Mhamphi, G.G.; Otieno, C.A.; Cook, E.A.J. Seroprevalence and associated risk factors of leptospirosis in slaughter pigs; a neglected public health risk, western Kenya. BMC Vet. Res. 2019, 15, 403. [Google Scholar] [CrossRef] [Green Version]
- Thibeaux, R.; Soupé-Gilbert, M.-E.; Kainiu, M.; Girault, D.; Bierque, E.; Fernandes, J.; Bähre, H.; Douyère, A.; Eskenazi, N.; Vinh, J.; et al. The zoonotic pathogen Leptospira interrogans mitigates environmental stress through cyclic-di-GMP-controlled biofilm production. NPJ Biofilms Microbiomes 2020, 6, 24. [Google Scholar] [CrossRef]
- Wynwood, S.J.; Graham, G.C.; Weier, S.L.; Collet, T.A.; McKay, D.B.; Craig, S.B. Leptospirosis from water sources. Pathog. Glob. Health 2014, 108, 334–338. [Google Scholar] [CrossRef]
- Bevins, S.N.; Pedersen, K.; Lutman, M.W.; Gidlewski, T.; Deliberto, T.J. Consequences associated with the recent range expansion of nonnative feral swine. Bioscience 2014, 64, 291–299. [Google Scholar] [CrossRef] [Green Version]
- Roney, M. Once Established, Feral Hogs Hard to Contain, Eradicate. AP News 2018. Available online: https://apnews.com/21973c7e025f4c48845178920e273e06 (accessed on 1 July 2020).
- Chatfield, J.; Milleson, M.; Stoddard, R.; Bui, D.M.; Galloway, R. Serosurvey of leptospirosis in feral hogs (Sus scrofa) in Florida. J. Zoo Wildl. Med. 2013, 44, 404–407. [Google Scholar] [CrossRef] [PubMed]
- Buchholz, A.E.; Katz, A.R.; Galloway, R.; Stoddard, R.A.; Goldstein, S.M. Feral Swine Leptospira Seroprevalence Survey in Hawaii, USA, 2007–2009. Zoonoses Public Health 2016, 63, 584–587. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, K.; Pabilonia, K.L.; Anderson, T.D.; Bevins, S.N.; Hicks, C.R.; Kloft, J.M.; Deliberto, T.J. Widespread detection of antibodies to Leptospira in feral swine in the United States. Epidemiol. Infect. 2015, 143, 2131–2136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, K.; Anderson, T.D.; Bevins, S.N.; Pabilonia, K.L.; Whitley, P.N.; Virchow, D.R.; Gidlewski, T. Evidence of leptospirosis in the kidneys and serum of feral swine (Sus scrofa) in the United States. Epidemiol. Infect. 2017, 145, 87–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaller, M.; Collier, B.; Achberger, E.; Barry, O. Detection of Feral Hog Impacts to Water Quality and Wildlife Final Report. 2000. Available online: https://www.wlf.louisiana.gov/assets/Resources/Publications/Feral_Hogs/Detection_of_Feral_Hog_Impacts_to_Water_Quality_and_Wildlife_2016.pdf (accessed on 1 July 2020).
- Thibeaux, R.; Geroult, S.; Benezech, C.; Chabaud, S.; Soupé-Gilbert, M.E.; Girault, D.; Bierque, E.; Goarant, C. Seeking the environmental source of Leptospirosis reveals durable bacterial viability in river soils. PLoS Negl. Trop. Dis. 2017, 11, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bierque, E.; Thibeaux, R.; Girault, D.; Soupé-Gilbert, M.E.; Goarant, C. A systematic review of Leptospira in water and soil environments. PLoS ONE 2020, 15, e0227055. [Google Scholar] [CrossRef]
- Aragón-Martínez, A.; Olivera-Gómez, L.D.; Jiménez-Domínguez, D. Seasonal prevalence of antibodies to Leptospira interrogans in Antillean manatees from a landlocked lake in Tabasco, Mexico. J. Wildl. Dis. 2014, 50, 505–511. [Google Scholar] [CrossRef] [Green Version]
- Barragan, V.; Chiriboga, J.; Miller, E.; Olivas, S.; Birdsell, D.; Hepp, C.; Hornstra, H.; Schupp, J.M.; Morales, M.; Gonzalez, M.; et al. High Leptospira diversity in animals and humans complicates the search for common reservoirs of human disease in rural Ecuador. PLoS Negl. Trop. Dis. 2016, 10, e0004990. [Google Scholar] [CrossRef]
- Hamond, C.; Pestana, C.P.; Medeiros, M.A.; Lilenbaum, W. Genotyping of Leptospira directly in urine samples of cattle demonstrates a diversity of species and strains in Brazil. Epidemiol. Infect. 2016, 144, 72–75. [Google Scholar] [CrossRef] [Green Version]
- Martins, G.; Loureiro, A.P.; Hamond, C.; Pinna, M.H.; Bremont, S.; Bourhy, P.; Lilenbaum, W. First isolation of Leptospira noguchii serogroups Panama and Autumnalis from cattle. Epidemiol. Infect. 2015, 143, 1538–1541. [Google Scholar] [CrossRef]
- Zarantonelli, L.; Suanes, A.; Meny, P.; Buroni, F.; Nieves, C.; Salaberry, X.; Briano, C.; Ashfield, N.; Da Silva Silveira, C.; Dutra, F.; et al. Isolation of pathogenic Leptospira strains from naturally infected cattle in Uruguay reveals high serovar diversity, and uncovers a relevant risk for human leptospirosis. PLoS Negl. Trop. Dis. 2008, 12, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Pellizzaro, M.; Conrado, F.D.O.; Martins, C.M.; Joaquim, S.F.; Ferreira, F.; Langoni, H.; Biondo, A.W. Serosurvey of Leptospira spp. and Toxoplasma gondii in rats captured from two zoos in Southern Brazil. Rev. Soc. Bras. Med. Trop. 2017, 50, 857–860. [Google Scholar] [CrossRef] [Green Version]
- Vieira, A.S.; D’Andrea, P.S.; Vilela, R.d.V.; Loretto, D.; Jaeger, L.H.; Carvalho-Costa, F.A.; Lilenbaum, W. Pathogenic Leptospira species are widely disseminated among small mammals in Atlantic Forest biome. Transbound. Emerg. Dis. 2019, 66, 1195–1201. [Google Scholar] [CrossRef] [PubMed]
- Ballados-González, G.G.; Sánchez-Montes, S.; Romero-Salas, D.; Colunga Salas, P.; Gutiérrez-Molina, R.; León-Paniagua, L.; Becker, I.; Méndez-Ojeda, M.L.; Barrientos-Salcedo, C.; Serna-Lagunes, R.; et al. Detection of pathogenic Leptospira species associated with phyllostomid bats (Mammalia: Chiroptera) from Veracruz, Mexico. Transbound. Emerg. Dis. 2018, 65, 773–781. [Google Scholar] [CrossRef] [PubMed]
- Flores, B.J.; Pérez-Sánchez, T.; Fuertes, H.; Sheleby-Elías, J.; Múzquiz, J.L.; Jirón, W.; Duttmann, C.; Halaihel, N. A cross-sectional epidemiological study of domestic animals related to human leptospirosis cases in Nicaragua. Acta Trop. 2017, 170, 79–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, É.F.; Cerqueira, G.M.; Seyffert, N.; Seixas, F.K.; Hartwig, D.D.; Athanazio, D.A.; Pinto, L.S.; Queiroz, A.; Ko, A.I.; Brod, C.S.; et al. Leptospira noguchii and human and animal Leptospirosis, Southern Brazil. Emerg. Infect. Dis. 2009, 15, 621–623. [Google Scholar] [CrossRef]
- Loureiro, A.P.; Jaeger, L.H.; Di Azevedo, M.I.N.; Miraglia, F.; Moreno, L.Z.; Moreno, A.M.; Pestana, C.P.; Carvalho-Costa, F.A.; Medeiros, M.A.; Lilenbaum, W. Molecular epidemiology of Leptospira noguchii reveals important insights into a One Health context. Transbound. Emerg. Dis. 2020, 67, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Richer, L.; Potula, H.H.; Melo, R.; Vieira, A.; Gomes-Solecki, M. Mouse model for sublethal Leptospira interrogans infection. Infect. Immun. 2015, 83, 4693–4700. [Google Scholar] [CrossRef] [Green Version]
- Dirsmith, K.; VanDalen, K.; Fry, T.; Charles, B.; VerCauteren, K.; Duncan, C. Leptospirosis in fox squirrels (Sciurus niger) of Larimer County, Colorado, USA. J. Wildl. Dis. 2013, 49, 641–645. [Google Scholar] [CrossRef] [Green Version]
- Miller, R.S.; Sweeney, S.J.; Slootmaker, C.; Grear, D.A.; Di Salvo, P.A.; Kiser, D.; Shwiff, S.A. Cross-species transmission potential between wild pigs, livestock, poultry, wildlife, and humans: Implications for disease risk management in North America. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef]
- Turner, L.H. Leptospirosis I. Trans. R. Soc. Trop. Med. Hyg. 1967, 61, 842–855. [Google Scholar] [CrossRef]
- Budihal, S.V.; Perwez, K. Leptospirosis diagnosis: Competancy of various laboratory tests. J. Clin. Diagn. Res. 2014, 8, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Kaltenboeck, B.; Wang, C. Advances in real-time PCR: Application to clinical laboratory diagnostics. Adv. Clin. Chem. 2005, 40, 219–259. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Loftis, A.; Ahluwalia, S.K.; Gao, G.; Verma, A.; Wang, C.; Kaltenboeck, B. Diagnosis of canine leptospirosis by a highly sensitive FRET-PCR targeting the lig genes. PLoS ONE 2014, 9, e89507. [Google Scholar] [CrossRef] [PubMed]
- Levett, P.N.; Morey, R.E.; Galloway, R.L.; Turner, D.E.; Steigerwalt, A.G.; Mayer, L.W. Detection of pathogenic Leptospires by real-time quantitative PCR. J. Med. Microbiol. 2005, 54, 45–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boonsilp, S.; Thaipadungpanit, J.; Amornchai, P.; Wuthiekanun, V.; Bailey, M.S.; Holden, M.T.; Zhang, C.; Jiang, X.; Koizumi, N.; Taylor, K.; et al. A single multilocus sequence typing (MLST) scheme for seven pathogenic Leptospira species. PLoS Negl. Trop. Dis. 2013, 7, e1954. [Google Scholar] [CrossRef] [Green Version]
- Lewis, J.S.; Corn, J.L.; Mayer, J.J.; Jordan, T.R.; Farnsworth, M.L.; Burdett, C.L.; Ver Cauteren, K.C.; Sweeney, S.J.; Miller, R.S. Historical, current and potential population size estimates of invasive wild pigs (Sus scrofa) in the United States. Biol. Invasions 2019, 21, 2373–2384. [Google Scholar] [CrossRef]
- Wei, L.; Kelly, P.; Zhang, J.; Yang, Y.; Zheng, X.; Tao, J.; Zhang, Z.; Wang, C. Use of a universal hydroxymethylbilane synthase (HMBS)-based PCR as an endogenous internal control and to enable typing of mammalian DNAs. Appl. Microbiol. Biotechnol. 2014, 98, 5579–5587. [Google Scholar] [CrossRef]


{kind=link}
| Specimen Type | Sample ID | Sampling Dates | Species Identified by PCR | Anti-Leptospira IgG ELISA (OD Value) |
|---|---|---|---|---|
| Kidney | EAPL-1-2 | July 19 | L. interrogans | 2.7 |
| EAPL-5-7 | July 19 | L. interrogans | 1.36 | |
| EAPL-25-5 | August 19 | L. interrogans | 1.33 | |
| EAPL-30-18 | August 19 | L. noguchii | 1.57 | |
| EAPL-30-20 | August 19 | L. noguchii | 1.18 | |
| EAPL-50-4 | August 19 | L. noguchii | 2.15 | |
| EAPL-20-5 | September 19 | L. noguchii | 2.29 | |
| EAPL-36-12 | September 19 | L. noguchii | 3.22 | |
| EAPL-25-17 | November 19 | L. interrogans | 1.23 | |
| EAPL-5-4-9 | January 20 | L. noguchii | 1.21 | |
| Soil sediment | EP-2B-8 | July 2019 | L. noguchii | Not applicable |
| Stream water | Tusk-1W | July 2019 | L. interrogans | Not applicable |
| Sampling Time | Total Assayed (N) | Positive (N) | Positivity (Confidence Interval) |
|---|---|---|---|
| July 2019 | 61 | 37 | 60.6% a (48.4%, 72.9%) |
| August 2019 | 35 | 20 | 57.1% a (40.8%, 73.5%) |
| September 2019 | 54 | 26 | 48.1% a (34.8%, 61.5%) |
| October 2019 | 46 | 16 | 34.8% ab (21.0%, 48.6%) |
| November 2019 | 16 | 5 | 31.2% ab (8.5%, 54.0%) |
| December 2019 | 14 | 5 | 35.7% ab (10.6%, 60.8%) |
| January 2020 | 25 | 9 | 36.0 ab (17.2%, 54.8%) |
| February 2020 | 7 | 2 | 28.6% ab (8.2%, 64.1%) |
| March 2020 | 18 | 3 | 16.7% b (5.8%, 39.2%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poudel, A.; Hoque, M.M.; Madere, S.; Bolds, S.; Price, S.; Barua, S.; Adekanmbi, F.; Kalalah, A.; Kitchens, S.; Brown, V.; et al. Molecular and Serological Prevalence of Leptospira spp. in Feral Pigs (Sus scrofa) and their Habitats in Alabama, USA. Pathogens 2020, 9, 857. https://doi.org/10.3390/pathogens9100857
Poudel A, Hoque MM, Madere S, Bolds S, Price S, Barua S, Adekanmbi F, Kalalah A, Kitchens S, Brown V, et al. Molecular and Serological Prevalence of Leptospira spp. in Feral Pigs (Sus scrofa) and their Habitats in Alabama, USA. Pathogens. 2020; 9(10):857. https://doi.org/10.3390/pathogens9100857
Chicago/Turabian StylePoudel, Anil, Md Monirul Hoque, Steven Madere, Sara Bolds, Stuart Price, Subarna Barua, Folasade Adekanmbi, Anwar Kalalah, Steven Kitchens, Vienna Brown, and et al. 2020. "Molecular and Serological Prevalence of Leptospira spp. in Feral Pigs (Sus scrofa) and their Habitats in Alabama, USA" Pathogens 9, no. 10: 857. https://doi.org/10.3390/pathogens9100857