Establishment of an In Vitro Model of Persistent Chicken Anemia Virus Infection

 and
and
Abstract
:1. Introduction
2. Results
2.1. Cell Viability
2.2. Detection of CAV Antigens
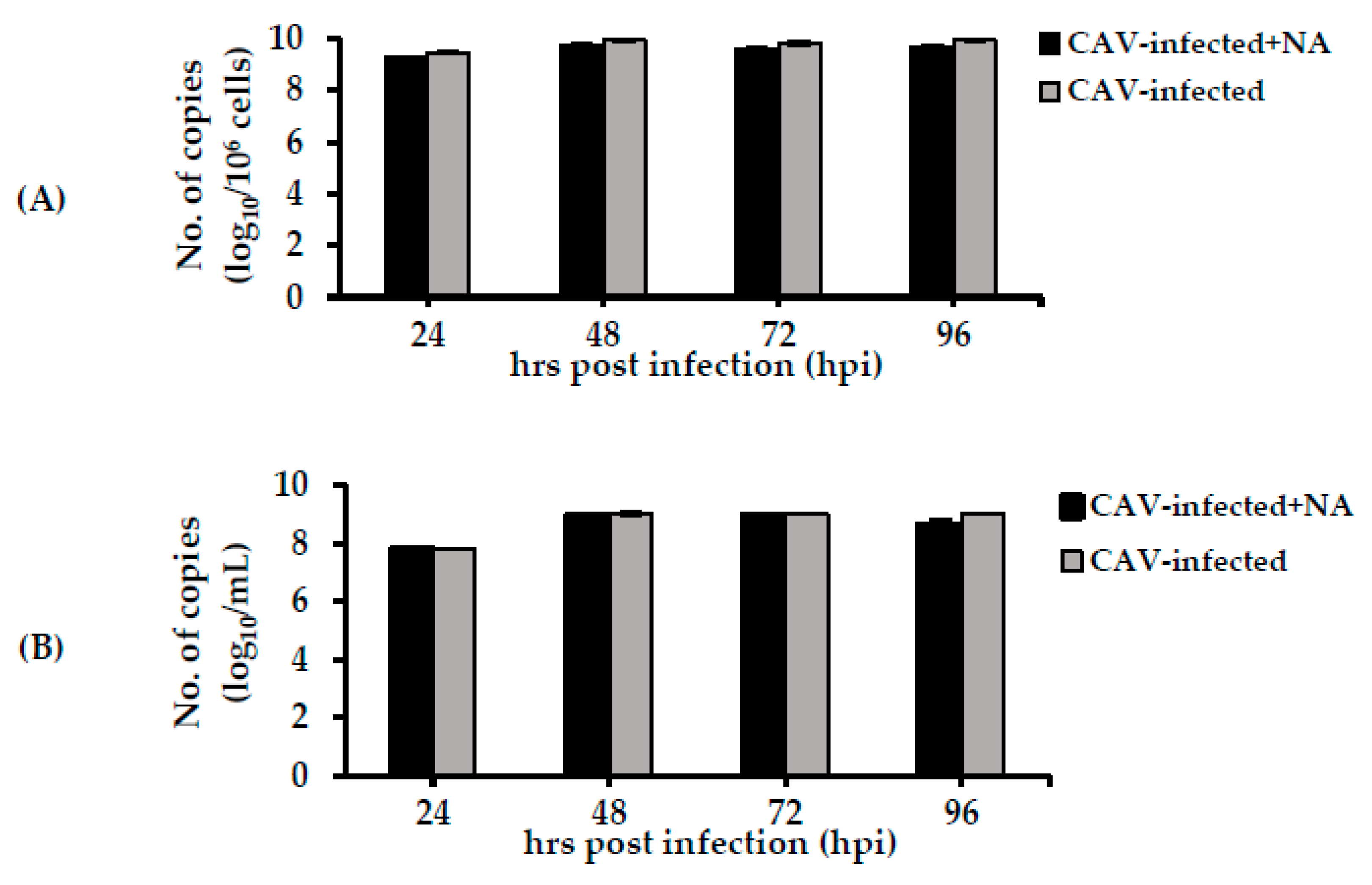
2.3. Detection of the CAV Genome within Infected Cells and Cell Supernatants
2.4. Recovery of CAV during Passage
2.5. Genetic Analyses of Virus Recovered from Passaged Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture, Virus, Titration, and Virus Isolation
4.2. Antiserum, Monoclonal Antibodies, and Anti-Peptide Antibodies
4.3. Detection of Cell Viability
4.4. DNA Extraction and Real-Time PCR
4.5. IFAT
4.6. Experimental Design for In Vitro Persistent Infection
4.7. Nucleotide Sequencing and Genetic Analyses
4.8. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rosario, K.; Breitbart, M.; Harrach, B.; Segales, J.; Delwart, E.; Biagini, P.; Varsani, A. Revisiting the taxonomy of the family Circoviridae: Establishment of the genus Cyclovirus and removal of the genus Gyrovirus. Arch. Virol. 2017, 162, 1447–1463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noteborn, M.H.; de Boer, G.F.; van Roozelaar, D.J.; Karreman, C.; Kranenburg, O.; Vos, J.G.; Jeurissen, S.H.; Hoeben, R.C.; Zantema, A.; Koch, G.; et al. Characterization of cloned chicken anemia virus DNA that contains all elements for the infectious replication cycle. J. Virol. 1991, 65, 3131–3139. [Google Scholar] [CrossRef] [Green Version]
- Trinh, D.Q.; Ogawa, H.; Bui, V.N.; Baatartsogt, T.; Kizito, M.K.; Yamaguchi, S.; Imai, K. Characterization of mAbs to chicken anemia virus and epitope mapping on its viral protein, VP1. J. Gen. Virol. 2015, 96, 1086–1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeurissen, S.H.; Wagenaar, F.; Pol, J.M.; van der Eb, A.J.; Noteborn, M.H. Chicken anemia virus causes apoptosis of thymocytes after in vivo infection and of cell lines after in vitro infection. J. Virol. 1992, 66, 7383–7388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noteborn, M.H.; Todd, D.; Verschueren, C.A.; de Gauw, H.W.; Curran, W.L.; Veldkamp, S.; Douglas, A.J.; McNulty, M.S.; van der, E.A.; Koch, G. A single chicken anemia virus protein induces apoptosis. J. Virol. 1994, 68, 346–351. [Google Scholar] [CrossRef] [Green Version]
- Lai, G.H.; Lien, Y.Y.; Lin, M.K.; Cheng, J.H.; Tzen, J.T.; Sun, F.C.; Lee, M.S.; Chen, H.J. VP2 of chicken anaemia virus interacts with apoptin for down-regulation of apoptosis through de-phosphorylated threonine 108 on apoptin. Sci. Rep. 2017, 7, 14799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuasa, N.; Taniguchi, T.; Yoshida, I. Isolation and some characteristics of an agent inducing anemia in chicks. Avian Dis. 1979, 23, 366–385. [Google Scholar] [CrossRef]
- McNulty, M.S. Chicken anaemia agent: A review. Avian Pathol. 1991, 20, 187–203. [Google Scholar] [CrossRef] [PubMed]
- Hoop, R.K. Persistence and vertical transmission of chicken anaemia agent in experimentally infected laying hens. Avian Pathol. 1992, 21, 493–501. [Google Scholar] [CrossRef]
- Rosenberger, J.K.; Cloud, S.S. The effects of age, route of exposure, and coinfection with infectious bursal disease virus on the pathogenicity and transmissibility of chicken anemia agent (CAA). Avian Dis. 1989, 33, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Yuasa, N.; Yoshida, I. Experimental egg transmission of chicken anemia agent. Natl. Inst. Anim. Healthq. (Tokyo) 1983, 23, 99–100. [Google Scholar] [PubMed]
- Adair, B.M. Immunopathogenesis of chicken anemia virus infection. Dev. Comp. Immunol. 2000, 24, 247–255. [Google Scholar] [CrossRef]
- De Boer, G.F.; Van Roozelaar, D.J.; Moormann, R.J.; Jeurissen, S.H.; Wijngaard, J.C.; Hilbink, F.; Koch, G. Interaction between chicken anaemia virus and live Newcastle disease vaccine. Avian Pathol. 1994, 23, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Cui, N.; Han, N.; Wu, J.; Cui, Z.; Su, S. Depression of vaccinal immunity to Marek’s disease by infection with chicken infectious anemia virus. Front. Microbiol. 2017, 8, 1863. [Google Scholar] [CrossRef] [Green Version]
- Chettle, N.J.; Eddy, R.K.; Wyeth, P.J.; Lister, S.A. An outbreak of disease due to chicken anaemia agent in broiler chickens in England. Vet. Rec. 1989, 124, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Hoop, R.K.; Guscetti, F.; Keller, B. An outbreak of infectious chicken anemia in fattening chickens in Switzerland. Schweiz. Arch. Tierheilkd. 1992, 134, 485–489. [Google Scholar] [PubMed]
- Yuasa, N.; Imai, K.; Watanabe, K.; Saito, F.; Abe, M.; Komi, K. Aetiological examination of an outbreak of haemorrhagic syndrome in a broiler flock in Japan. Avian Pathol. 1987, 16, 521–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardona, C.J.; Oswald, W.B.; Schat, K.A. Distribution of chicken anaemia virus in the reproductive tissues of specific-pathogen-free chickens. J. Gen. Virol. 2000, 81, 2067–2075. [Google Scholar] [CrossRef]
- McNulty, M.S.; Connor, T.J.; McNeilly, F. A survey of specific pathogen-free chicken flocks for antibodies to chicken anaemia agent, avian nephritis virus and group a rotavirus. Avian Pathol. 1989, 18, 215–220. [Google Scholar] [CrossRef] [Green Version]
- Miles, M.M.; Oswald, W.B.; Scarlet, J.; Chat, K.A. Patterns chicken infectious anemia virus (CIAV) seroconversion in three Cornell SPF flock. In Proceeding of the Second International Symposium on Infectious Bursa Disease and Chicken Infectious Anemia, Rauischholzhausen, Germany, 21–24 June 2001; pp. 410–417. [Google Scholar]
- Schat, K.A.; Schukken, Y.H. An 8-year longitudinal survey for the presence of antibodies to chicken infectious anemia virus in two specific-pathogen-free strains of chickens. Avian Dis. 2010, 54, 46–52. [Google Scholar] [CrossRef]
- Brentano, L.; Lazzarin, S.; Bassi, S.S.; Klein, T.A.; Schat, K.A. Detection of chicken anemia virus in the gonads and in the progeny of broiler breeder hens with high neutralizing antibody titers. Vet. Microbiol. 2005, 105, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Imai, K.; Mase, M.; Tsukamoto, K.; Hihara, H.; Yuasa, N. Persistent infection with chicken anaemia virus and some effects of highly virulent infectious bursal disease virus infection on its persistency. Res. Vet. Sci. 1999, 67, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Notkins, A.L. Immune mechanisms by which the spread of viral infections is stopped. Cell. Immunol. 1974, 11, 478–483. [Google Scholar] [CrossRef]
- Schat, K.A.; van Santen, V.L. Chicken Infectious Anemia Virus and Other Circovirus Infections. In Diseases of Poultry, 12th ed.; Saif, J.R., McDougald, L.R., Nolan, L.K., Swayne, D.E., Eds.; John Wiley & Sons, Inc.: Blackwel, IA, USA; Hoboken, NJ, USA, 2008; pp. 211–223. [Google Scholar]
- Akiyama, Y.; Kato, S. Two cell lines from lymphomas of Marek’s disease. Biken. J. 1974, 17, 105–116. [Google Scholar]
- Yuasa, N.; Imai, K. Pathogenicity and antigenicity of eleven isolates of chicken anaemia agent (CAA). Avian Pathol. 1986, 15, 639–645. [Google Scholar] [CrossRef]
- Imai, K.; Yuasa, N. Development of a microtest method for serological and virological examinations of chicken anemia agent. Jpn. J. Vet. Sci. 1990, 52, 873–875. [Google Scholar] [CrossRef]
- Yuasa, N. Propagation and infectivity titration of the Gifu-1 strain of chicken anemia agent in a cell line (MDCC-MSB1) derived from Marek’s disease lymphoma. Natl. Inst. Anim. Healthq. 1983, 23, 13–20. [Google Scholar]
- Behrens, B.; Kärber, G. Wie sind Reihenversuche für biologishe Auswertungen am zweckmäßigsten anzuordnen? Naunyn. Schmiedebergs. Arch. Exp. Pathol. Pharmakol. 1934, 18, 379–388. [Google Scholar] [CrossRef]
- Dong, H.V.; Tran, G.T.H.; Nguyen, G.V.; Dao, T.D.; Bui, V.N.; Huynh, L.T.M.; Takeda, Y.; Ogawa, H.; Imai, K. Chicken anemia virus in northern Vietnam: Molecular characterization reveals multiple genotypes and evidence of recombination. Virus Genes 2019, 55, 643–653. [Google Scholar] [CrossRef] [PubMed]
- Olszewska-Tomczyk, M.; Swieton, E.; Minta, Z.; Smietanka, K. Occurrence and phylogenetic studies of chicken anemia virus from Polish broiler flocks. Avian Dis. 2016, 60, 70–74. [Google Scholar] [CrossRef]
- Yuasa, N.; Imai, K.; Tezuka, H. Survey of antibody against chicken anaemia agent (CAA) by an indirect immunofluorescent antibody technique in breeder flocks in Japan. Avian Pathol. 1985, 14, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, Y.; Wu, B.; Sun, B.; Chen, F.; Ji, J.; Ma, J.; Xie, Q. Phylogenetic and molecular characterization of chicken anemia virus in southern China from 2011 to 2012. Sci. Rep. 2013, 3, 3519. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | CAV Isolation (TCID50/106 cells) | |||
|---|---|---|---|---|
| No. of Passages | ||||
| P1 | P5 | P10 | P14 | |
| cells | 5.32 | 6.04 | 5.9 | 6.28 |
| sup | <0.7 | <0.7 | <0.7 | <0.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Dong, H.; Tran, G.T.H.; Trinh, D.Q.; Takeda, Y.; Ogawa, H.; Imai, K. Establishment of an In Vitro Model of Persistent Chicken Anemia Virus Infection. Pathogens 2020, 9, 842. https://doi.org/10.3390/pathogens9100842
Van Dong H, Tran GTH, Trinh DQ, Takeda Y, Ogawa H, Imai K. Establishment of an In Vitro Model of Persistent Chicken Anemia Virus Infection. Pathogens. 2020; 9(10):842. https://doi.org/10.3390/pathogens9100842
Chicago/Turabian StyleVan Dong, Hieu, Giang Thi Huong Tran, Dai Quang Trinh, Yohei Takeda, Haruko Ogawa, and Kunitoshi Imai. 2020. "Establishment of an In Vitro Model of Persistent Chicken Anemia Virus Infection" Pathogens 9, no. 10: 842. https://doi.org/10.3390/pathogens9100842