Characterization of Salmonella Isolates from Wastewater Treatment Plant Influents to Estimate Unreported Cases and Infection Sources of Salmonellosis



Abstract
:1. Introduction
2. Results
2.1. Prevalence of Salmonella Isolates from Sewage Influent Samples and Humans
2.2. Antimicrobial Susceptibility
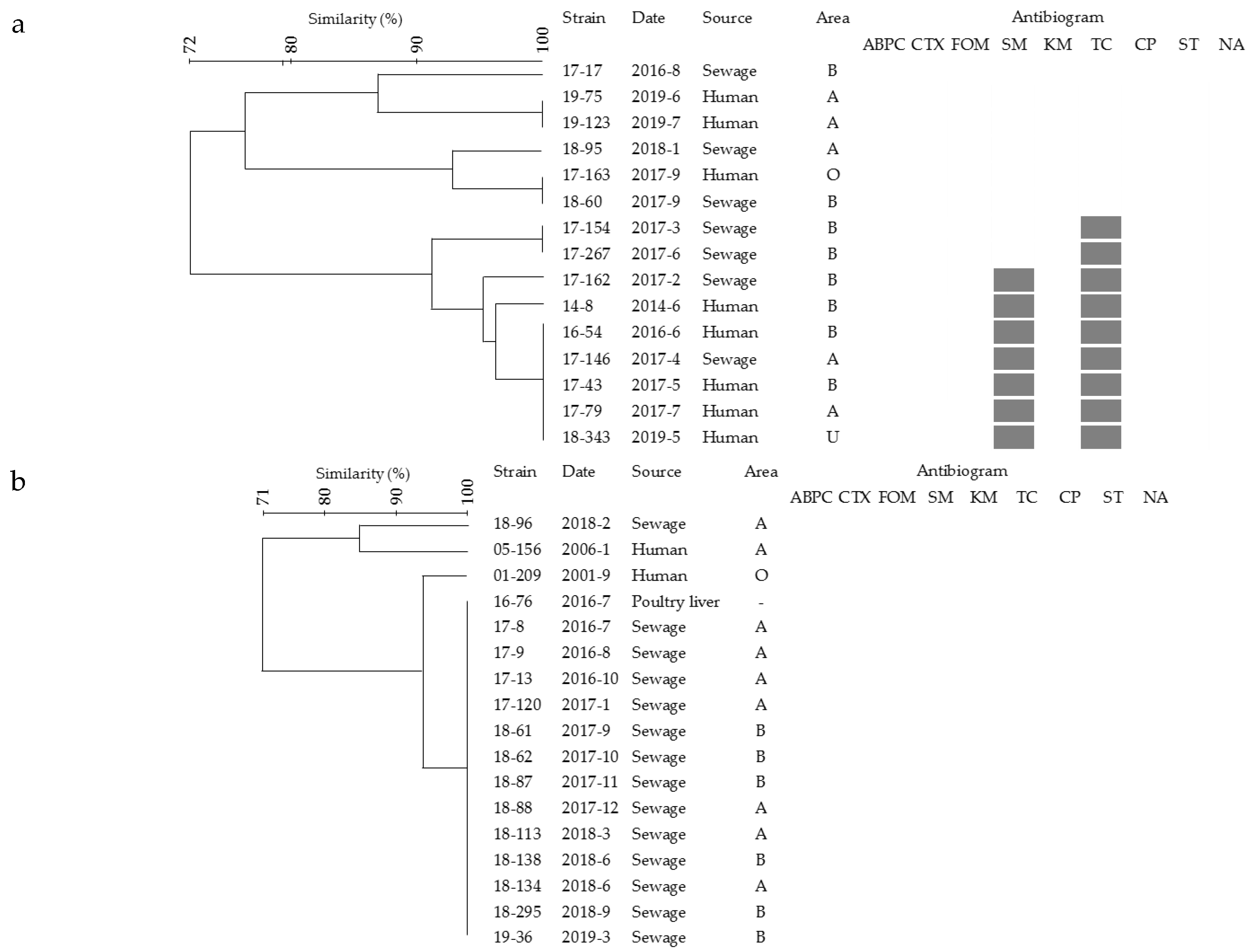
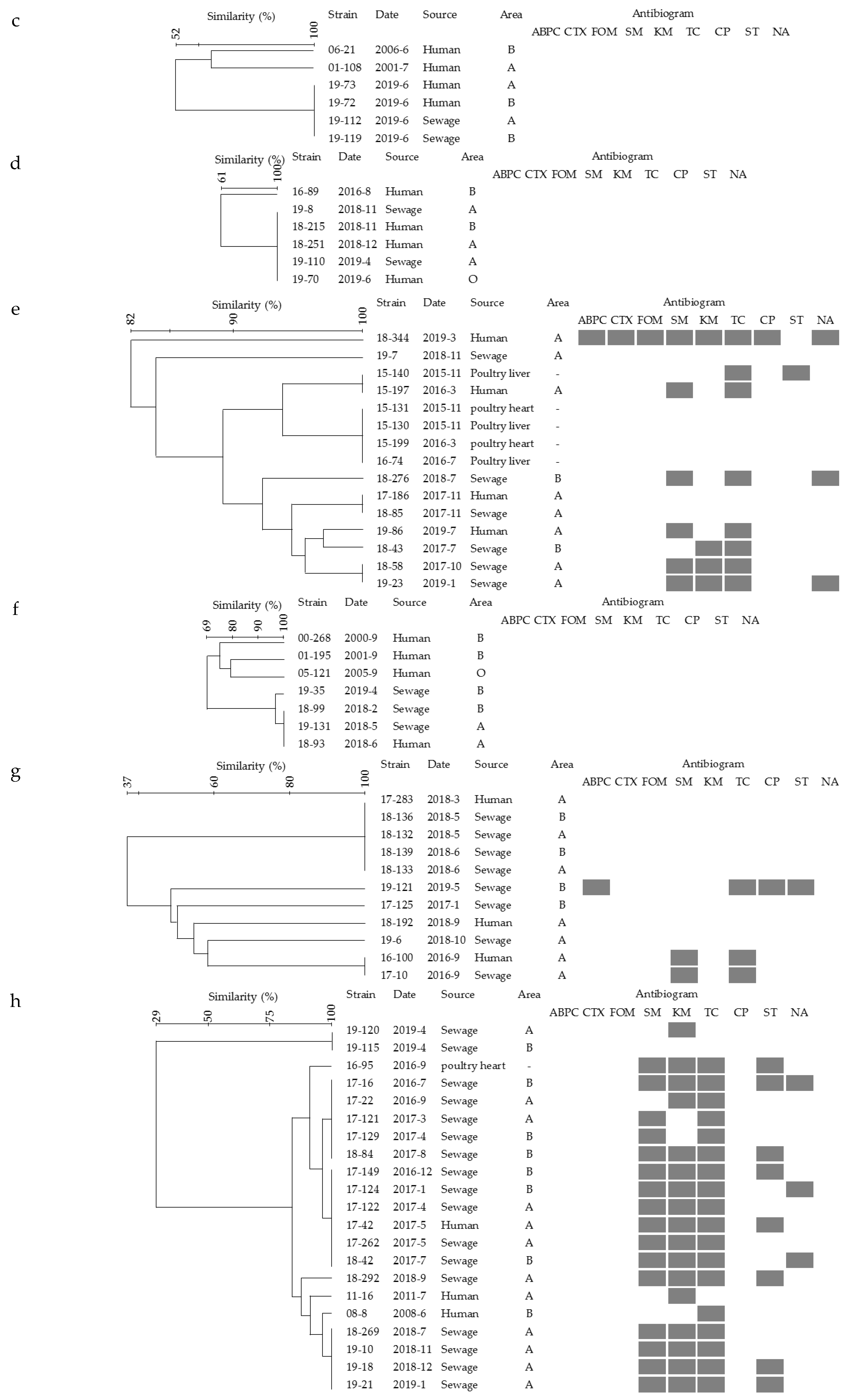
2.3. Pulsed-Field Gel Electrophoresis (PFGE) Analysis
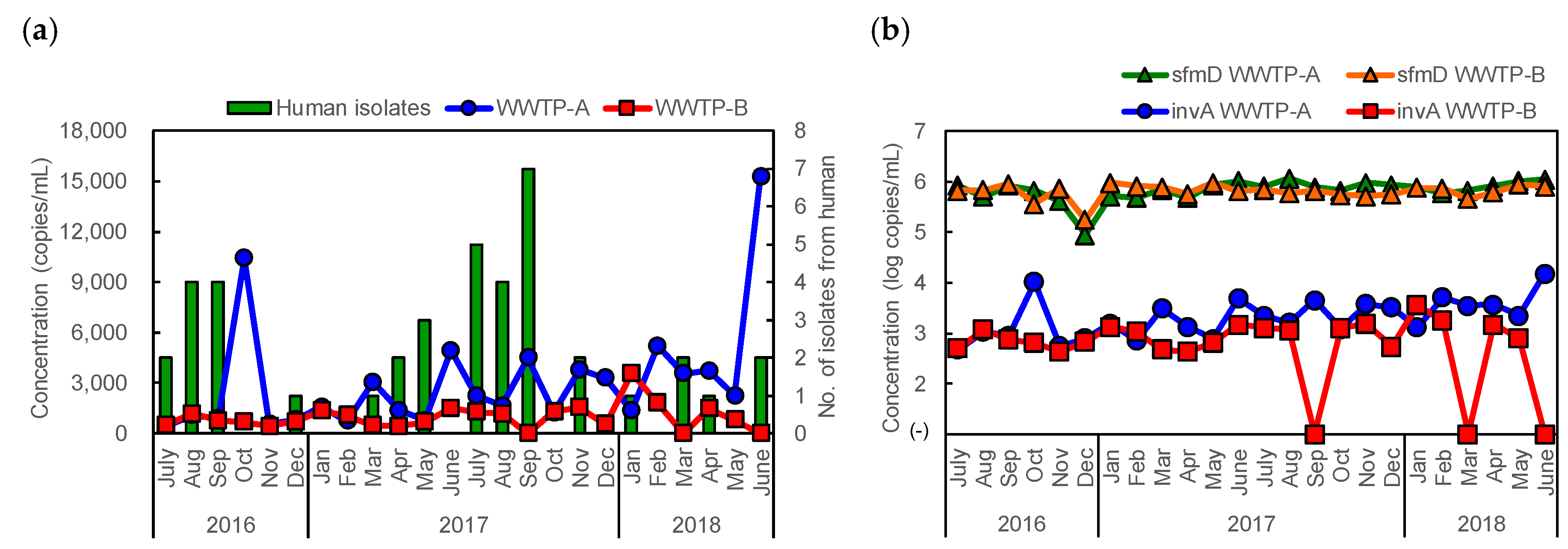
2.4. Quantification of Salmonella from Sewage Influent Samples
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Isolation of Salmonella
4.3. Strains
4.4. Antimicrobial Susceptibility Test
4.5. PFGE Analysis
4.6. Quantification of Salmonella
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Majowicz, S.E.; Musto, J.; Scallan, E.; Angulo, F.J.; Kirk, M.; O’brien, S.J.; Jones, T.F.; Fazil, A.; Hoekstra, R.M. The global burden of non-typhoidal Salmonella gastroenteritis. Clin Infect Dis. 2010, 50, 882–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Issenhuth-Jeanjean, S.; Roggentin, P.; Mikoleit, M.; Guibourdenche, M.; de Pinna, E.; Nair, S.; Fields, P.I.; Weill, F.X. Supplement 2008–2010 (no. 48) to the White–Kauffmann–Le Minor scheme. Res. Microbiol. 2014, 165, 526–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrari, R.G.; Rosario, D.K.A.; Cunha-Neto, A.; Mano, S.B.; Figueiredo, E.E.S.; Conte-Junior, C.A. Worldwide epidemiology of Salmonella serovars in animal-based foods: A meta-analysis. Appl. Environ. Microbiol. 2019, 85, e00591-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ministry of Health, Labor and Welfare (MHLW). Statistics of Food Poisoning in Japan. Available online: http://www.mhlw.go.jp/stf/seisakunitsuite/bunya/kenkou_iryou/shokuhin/syokuchu/04.html (accessed on 4 October 2019). (In Japanese).
- Pires, S.M.; Evers, E.G.; van Pelt, W.; Ayers, T.; Scallan, E.; Angulo, F.J.; Havelaar, A.; Hald, T. The Med-Vet-Net Workpackage 28 Working Group. Attributing the human disease burden of foodborne infections to specific sources. Foodborne Pathog. Dis. 2009, 6, 417–424. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, I.; Ishihara, T.; Teranishi, H.; Saito, S.; Yatsuyanagi, J.; Wada, E.; Wada, E.; Kumagai, Y.; Takahashi, S.; Konno, T.; et al. Prevalence and Characteristics of Salmonella and Campylobacter in retail poultry meat in Japan. Jpn. J. Infect. Dis. 2017, 70, 239–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiroi, M.; Kawamori, F.; Harada, T.; Sano, Y.; Miwa, N.; Sugiyama, K.; Hara-Kudo, Y.; Masuda, T. Antibiotic resistance in bacterial pathogens from retail raw meats and food-producing animals in Japan. J. Food Prot. 2012, 75, 1774–1782. [Google Scholar] [CrossRef]
- Hara-Kudo, Y.; Konuma, H.; Kamata, Y.; Miyahara, M.; Takatori, K.; Onoue, Y.; Sugita-Konishi, Y.; Ohnishi, T. Prevalence of the main food-borne pathogens in retail food under the national food surveillance system in Japan. Food Addit. Contam. Part A 2012, 30, 1450–1458. [Google Scholar] [CrossRef]
- Tauxe, R.V. Emerging foodborne diseases: An evolving public health challenge. Emerg. Infect. Dis. 1997, 3, 425–434. [Google Scholar] [CrossRef] [Green Version]
- Berge, A.C.B.; Dueger, E.L.; Sischo, W.M. Comparison of Salmonella enterica serovar distribution and antibiotic resistance patterns in wastewater at municipal water treatment plants in two California cities. J. Appl. Microbiol. 2006, 101, 1309–1316. [Google Scholar] [CrossRef]
- Haramoto, E.; Katayama, H.; Oguma, K.; Yamashita, H.; Tajima, A.; Nakajima, H.; Ohgaki, S. Seasonal profiles of human noroviruses and indicator bacteria in wastewater treatment plant in Tokyo, Japan. Water Sci. Technol. 2006, 54, 301–308. [Google Scholar] [CrossRef] [Green Version]
- Haramoto, E.; Katayama, H.; Oguma, K.; Ohgaki, S. Quantitative analysis of human enteric adenoviruses in aquatic environments. J. Appl. Microbiol. 2007, 103, 2153–2159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haramoto, E.; Katayama, H.; Phanuwan, C.; Ohgaki, S. Quantitative detection of sapoviruses in wastewater and river water in Japan. Lett. Appl. Microbiol. 2008, 46, 408–413. [Google Scholar] [CrossRef] [PubMed]
- Bisseux, M.; Colombet, J.; Mirand, A.; Roque-Afonso, A.M.; Abravanel, F.; Izopet, J.; Archimbaud, C.; Peigue-Lafeuille, H.; Debroas, D.; Bailly, J.-L.; et al. Monitoring human enteric viruses in wastewater and relevance to infections encountered in the clinical setting: A one-year experiment in central France, 2014 to 2015. Euro. Surveill. 2018, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, T.; O’Brien, P.; Shelton, J.M.; Whelen, A.C.; Pagaling, E. Municipal wastewater as a microbial surveillance platform for enteric diseases: A case study for Salmonella and salmonellosis. Environ. Sci. Technol. 2018, 52, 4869–4877. [Google Scholar] [CrossRef] [PubMed]
- Diemert, S.; Yan, T. Clinically unreported salmonellosis outbreak detected via comparative genomic analysis of municipal wastewater Salmonella isolates. Appl. Environ. Microbiol. 2019, 85, e00139-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reilly, W.J.; Oboegbulem, S.I.; Munro, D.S.; Forbes, G.I. The epidemiological relationship between Salmonella isolated from poultry meat and sewage effluents at a long-stay hospital. Epidemiol. Infect. 1991, 106, 1–10. [Google Scholar] [CrossRef]
- Ostoich, M.; Aimo, E.; Frate, R.; Vazzoler, M.; Stradella, S.; Osti, P. Integrated approach for microbiological impact assessment of public wastewater treatment plants. Chem. Ecol. 2007, 23, 43–62. [Google Scholar] [CrossRef]
- Yang, K.; Pagaling, E.; Yan, T. Estimating the Prevalence of Potential Enteropathogenic Escherichia coli and Intimin Gene Diversity in a Human Community by Monitoring Sanitary Sewage. Appl. Environ. Microbiol. 2014, 80, 119–127. [Google Scholar] [CrossRef] [Green Version]
- Kacprzak, M.; Fijalkowski, K.; Grobelak, A.; Rosikon, K.; Rorat, A. Eschrichia coli and Salmonella spp. early diagnosis and seasonal monitoring in the sewage treatment process by EMA-qPCR method. Pol. J. Microbiol. 2015, 64, 143–148. [Google Scholar] [CrossRef] [Green Version]
- Nair, A.; Balasaravanan, T.; Malik, S.V.; Mohan, V.; Kumar, M.; Vergis, J.; Rawool, D.B. Isolation and identification of Salmonella from diarrheagenic infants and young animals, sewage waste and fresh vegetables. Vet. World 2015, 8, 669–673. [Google Scholar] [CrossRef]
- Bonetta, S.; Pignata, C.; Lorenzi, E.; De Ceglia, M.; Meucci, L.; Bonetta, S.; Gilli, G.; Carraro, E. Detection of pathogenic Campylobacter, E. coli O157:H7 and Salmonella spp. in wastewater by PCR assay. Environ. Sci. Pollut. Res. 2016, 23, 15302–15309. [Google Scholar] [CrossRef] [PubMed]
- Zang, C.M.; Xu, L.M.; Mou, X.; Xu, H.; Liu, J.; Miao, Y.H.; Wang, X.C.; Li, X. Characterization and evolution of antibiotic resistance of Salmonella in municipal wastewater treatment plants. J. Environ. Manag. 2019, 251, 109547. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, E.; Torii, Y.; Shigemura, H.; Ishige, T.; Yanagimoto, K.; Uematsu, K.; Ando, N.; Murakami, S. Isolation of Salmonella enterica serovar Agona strains and their similarities to strains derived from a clone caused a serovar shift in broilers. J. Infect. Chemother. 2019, 25, 71–74. [Google Scholar] [CrossRef] [Green Version]
- Torii, Y.; Yokoyama, E.; Seki, M.; Shigemura, H.; Ishige, T.; Yanagimoto, K.; Uematsu, K.; Ando, N.; Fujimaki, T.; Murakami, S. Genetic characteristics of emerging Salmonella enterica serovar Agona strains isolated from humans in the prior period to occurrence of the serovar shift in broilers. J. Vet. Med. Sci. 2019, 81, 1117–1120. [Google Scholar] [CrossRef] [Green Version]
- Gutema, F.D.; Agga, G.E.; Abdi, R.D.; De Zutter, L.; Duchateau, L.; Gabriël, S. Prevalence and Serotype Diversity of Salmonella in Apparently Healthy Cattle: Systematic Review and Meta-Analysis of Published Studies, 2000–2017. Front. Vet. Sci. 2019, 6, 102. [Google Scholar] [CrossRef]
- Watanabe, R.; Ogawa, K.; Kubota, A.; Morimoto, Y. Serotype frequency of Salmonella isolates from human in Hokkaido. Rep. Hokkaido Inst. Pub. Health 2016, 66, 65–67. (In Japanese) [Google Scholar]
- Kurogi, M.; Yoshino, S.; Ganmyo, H.; Nagano, K. Distribution of serovars of Salmonella strains isolated during 2008-2014 in Miyazaki prefecture. Annu. Rep. Miyazaki Pref. Inst. Pub. Health Environ. 2014, 26, 67–71. (In Japanese) [Google Scholar]
- National Institute of Infectious Diseases. Infectious Agents Surveillance Report. Available online: https://nesid4g.mhlw.go.jp/Byogentai/Pdf/data81j.pdf (accessed on 24 October 2019). (In Japanese)
- Dione, M.M.; Ikumapayi, U.N.; Saha, D.; Mohammed, N.I.; Geerts, S.; Ieven, M.; Adegbola, R.A.; Antonio, M. Clonal differences between Non-Typhoidal Salmonella (NTS) recovered from children and animals living in close contact in the Gambia. PLoS Negl. Trop. Dis. 2011, 5, e1148. [Google Scholar] [CrossRef]
- Traoré, O.; Nyholm, O.; Siitonen, A.; Bonkoungou, I.J.; Traoré, A.S.; Barro, N.; Haukka, K. Prevalence and diversity of Salmonella enterica in water, fish and lettuce in Ouagadougou, Burkina Faso. BMC Microbiol. 2015, 15, 151. [Google Scholar] [CrossRef]
- Kaneko, M.; Noda, H.; Ohnuma, M. Serovars and epidemiological properties of Salmonella isolated from patients with sporadic diarrhea in Yamanashi prefecture during the last 22 years (1985-2006). Kansenshogaku Zasshi 2007, 81, 394–402. (In Japanese) [Google Scholar] [CrossRef]
- Yanagimoto, K.; Ohnuma, M.; Uematsu, K. Antimicrobial resistance of Salmonella strains isolated from patients with sporadic diarrhea in Yamanashi prefecture, 2007-2010. Annu. Rep. Yamanashi Inst. Pub. Health 2011, 55, 65–70. (In Japanese) [Google Scholar]
- Oberreuter, H.; Rau, J. Artificial neural network-assisted Fourier transform infrared spectroscopy for differentiation of Salmonella serogroups and its application on epidemiological tracing of Salmonella Bovismorbificans outbreak isolates from fresh sprouts. FEMS Microbiol. Lett. 2019, 366, fnz193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandwagt, D.; van den Wijngaard, C.; Tulen, A.D.; Mulder, A.C.; Hofhuis, A.; Jacobs, R.; Heck, M.; Verbruggen, A.; van den Kerkhof, H.; Slegers-Fitz-James, I.; et al. Outbreak of Salmonella Bovismorbificans associated with the consumption of uncooked ham products, the Netherlands, 2016 to 2017. Euro Surveill. 2018, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murakami, S.; Seki, M.; Suzuki, Y.; Takano, A.; Suzuki, K.; Okazaki, M.; Kobayashi, T.; Torii, Y.; Fujimaki, T.; Yokoyama, E. Investigation of Salmonella Agona colonized in broiler chickens and Salmonella contamination in minced chicken. J. Jpn. Soc. Poult. Dis. 2016, 52, 180–184. (In Japanese) [Google Scholar]
- Centers for Disease Control and Prevention. Multistate Outbreak of Salmonella Mbandaka Infection Linked to Kellogg’s Honey Smacks Cereal (Final Update). Available online: https://www.cdc.gov/salmonella/mbandaka-06-18/index.html (accessed on 11 October 2019).
- Dickter, J.K.; Cai, L.; Snyder, D.S. Endocarditis following Consumption of Cereal Associated with Salmonella enterica Subtype Mbandaka Outbreak. Case Rep. Infect. Dis. 2019, 2019. [Google Scholar] [CrossRef] [Green Version]
- Onwuezobe, I.A.; Oshun, P.O.; Odigwe, C.C. Antimicrobials for treating symptomatic non-typhoidal Salmonella infection. Cochrane Database Syst. Rev. 2019, 11. [Google Scholar] [CrossRef]
- Ono, K. Quantitative contamination level of Campylobacter and Salmonella species in commercial chicken meat and their drug susceptibility. J. Jpn. Vet. Med. Assoc. 2014, 67, 442–448. (In Japanese) [Google Scholar] [CrossRef]
- Noda, H.; Ohnuma, M.; Mikami, M.; Kaneko, M. Salmonella cell count in feces of patients with sporadic diarrhea. Annu. Rep. Yamanashi Inst. Pub. Health Environ. 2002, 46, 23–26. (In Japanese) [Google Scholar]
- Imanishi, M.; Rotstein, D.S.; Reimschuessel, R.; Schwensohn, C.A.; Woody, D.H., Jr.; Davis, S.W.; Hunt, A.D.; Arends, K.D.; Achen, M.; Cui, J. Outbreak of Salmonella enterica serotype Infantis infection in humans linked to dry dog food in the United States and Canada, 2012. J. Am. Vet. Med. Assoc. 2014, 244, 545–553. [Google Scholar] [CrossRef]
- Uematsu, K.; Yanagimoto, K. Health hazard risk of infection caused by raw horseflesh, chicken, and beef offal contaminated with organisms Salmonella, Campylobacter jejuni/coli and enterohemorrhagic Escherichia coli. Annu. Rep. Yamanashi Inst. Pub. Health Environ. 2017, 61, 41–43. (In Japanese) [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Disk Susceptibility Tests, 13th ed.; M02; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Ribot, E.M.; Fair, M.A.; Gautom, R.; Cameron, D.N.; Hunter, S.B.; Swaminathan, B.; Barrett, T.J. Standardization of pulsed-field gel electrophoresis protocols for the subtyping of Escherichia coli O157:H7, Salmonella, and Shigella for PulseNet. Foodborne Pathog Dis. 2006, 3, 59–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iijima, Y.; Asako, N.T.; Aihara, M.; Hayashi, K. Improvement in the detection rate of diarrhoeagenic bacteria in human stool specimens by a rapid real-time PCR assay. J. Med. Microbiol. 2004, 53, 617–622. [Google Scholar] [CrossRef] [PubMed]
- Kaclíková, E.; Pangallo, D.; Oravcová, K.; Drahovská, H.; Kuchta, T. Quantification of Escherichia coli by kinetic 5’-nuclease polymerase chain reaction (real-time PCR) oriented to sfmD gene. Lett. Appl. Microbiol. 2005, 41, 132–135. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Serotype | 2016 | 2017 | 2018 | 2019 | Total (%) | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 7 | 8 | 9 | 10 | 11 | 12 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 1 | 2 | 3 | 4 | 5 | 6 | |||
| Schwarz-engrund | B | - | A | - | - | B | B | - | A | A, B | A | - | B | B | - | - | - | - | - | - | - | - | - | - | A | - | A | - | A | A | A | - | - | A, B | - | - | 17 | (11) |
| Anatum | A | A | - | A | - | - | A | - | - | - | - | - | - | - | B | B | B | A | - | A | A | - | - | A, B | - | - | B | - | - | - | - | - | B | - | - | - | 14 | (9) |
| Newport | - | A | - | - | - | - | B | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | A, B | A, B | - | - | - | A | - | - | - | - | - | - | B | - | 8 | (5) |
| Saintpaul | - | - | - | - | A | - | B | - | A | - | - | B | - | B | - | A | - | B | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 7 | (5) |
| Agona | - | B | - | - | - | - | - | B | B | A | - | B | - | - | B | - | - | - | A | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 7 | (5) |
| Thompson | - | - | - | - | B | B | - | - | - | - | - | - | - | - | - | - | - | - | - | - | A | - | - | - | - | - | B | - | B | A | - | A | - | - | - | - | 7 | (5) |
| Bareilly | - | - | - | A | - | - | - | - | - | - | - | - | - | - | A | - | - | - | - | - | - | A | - | - | A | - | A | B | - | - | - | B | - | - | - | - | 7 | (5) |
| Infantis | - | - | - | - | - | - | - | - | - | - | - | - | B | - | - | A | A | - | - | - | - | - | - | - | B | - | - | - | A | - | A | - | - | - | - | - | 6 | (4) |
| Braenderup | - | - | - | - | - | - | - | - | - | - | - | - | - | B | - | - | - | - | - | - | - | - | - | - | - | - | - | - | B | B | A, B | - | - | - | - | - | 5 | (3) |
| Chester | - | - | - | - | - | B | - | B | B | - | - | A | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | B | - | - | 5 | (3) |
| Stanley | - | - | A | - | - | - | A | - | A | - | - | - | - | A | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 4 | (3) |
| Mikaw-ashima | - | - | - | - | - | - | - | - | - | B | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | B | B | - | - | - | - | B | - | - | - | - | 4 | (3) |
| Corvallis | - | - | - | - | - | - | A | - | A | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | A | B | - | - | - | - | - | - | - | - | - | - | 4 | (3) |
| Hvittingfoss | - | - | B | - | - | - | - | - | - | - | A | - | A | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 3 | (2) |
| Blockley | - | - | - | - | - | - | - | - | B | B | B | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 3 | (2) |
| London | - | - | - | B | - | - | - | - | B | - | - | - | - | - | - | - | - | - | - | - | - | B | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 3 | (2) |
| Mbandaka | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | B | - | - | A | - | - | - | - | - | - | - | - | - | B | - | - | - | 3 | ( 2) |
| Cerro | - | - | - | - | - | - | - | - | - | - | - | - | - | A | A | - | - | - | - | - | - | - | - | - | - | A | - | - | - | - | - | - | - | - | - | - | 3 | ( 2) |
| Altona | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | A | A | - | B | - | 3 | ( 2) |
| Rissen | - | - | - | - | - | A | - | - | - | - | - | B | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 2 | ( 1) |
| Vitkin | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | B | B | - | - | - | - | - | - | - | - | - | - | - | - | - | 2 | ( 1) |
| Colindale | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | A, B | - | - | - | - | - | - | - | - | - | - | 2 | ( 1) |
| Brande-nburg | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | A | - | - | - | - | A | - | - | 2 | ( 1) |
| Bovismo-rbificans | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | A, B | 2 | ( 1) |
| Litchfield | - | - | A | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | ( 1) |
| Aberdeen | - | - | - | - | - | - | B | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | ( 1) |
| Typhim-urium | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | A | - | - | - | - | 1 | ( 1) |
| Derby | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | B | - | - | - | - | 1 | ( 1) |
| Ebrie | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | B | - | - | 1 | ( 1) |
| Javiana | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | A | - | - | 1 | ( 1) |
| Untypable (O13:z29:-) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | B | - | B | A, B | B | B | B | - | - | - | - | - | - | - | - | - | - | - | 7 | ( 5) |
| Untypable (O7:HNM) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | B | B | - | - | - | - | - | - | - | - | - | - | A | - | - | - | - | - | - | - | 3 | ( 2) |
| Untypable (O8:-:1,5) | - | - | - | - | - | - | - | - | - | - | - | - | - | B | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | ( 1) |
| Others | - | - | A | - | - | - | B | - | - | B | - | A | B | - | - | - | - | B | B | - | A | - | - | B | A | - | - | - | - | A, B | - | B | B | B | A | A | 17 | (11) |
| Total | 2 | 3 | 5 | 3 | 2 | 4 | 8 | 2 | 8 | 6 | 3 | 5 | 4 | 6 | 4 | 3 | 3 | 4 | 3 | 2 | 4 | 5 | 5 | 6 | 6 | 5 | 5 | 2 | 6 | 5 | 4 | 7 | 4 | 7 | 3 | 3 | 157 | |
| Serotype | Sources | |||||||
|---|---|---|---|---|---|---|---|---|
| Sewage | Human | |||||||
| No. of Isolates (%) | No. of Antimicrobial-Resistant Isolates (Rate of Resistance; %) | No. of Isolates (%) | No. of Antimicrobial-Resistant Isolates (Rate of Resistance; %) | |||||
| Agona | 7 | (5) | 4 | (57) | 7 | (9) | 4 | (57) |
| Stanley | 4 | (3) | 0 | 6 | (8) | 0 | ||
| Saintpaul | 7 | (5) | 0 | 5 | (7) | 2 | (40) | |
| Brandenburg | 2 | (1) | 0 | 5 | (7) | 0 | ||
| Thompson | 4 | (3) | 1 | (25) | 4 | (5) | 0 | |
| Typhimurium | 1 | (1) | 1 | (100) | 4 | (5) | 2 | (50) |
| Enteritidis | 0 | (0) | 0 | 4 | (5) | 0 | ||
| Bareilly | 7 | (5) | 0 | 4 | (5) | 0 | ||
| Infantis | 6 | (4) | 4 | (67) | 4 | (5) | 3 | (75) |
| Nagoya | 0 | (0) | 0 | 3 | (4) | 0 | ||
| Newport | 8 | (5) | 2 | (25) | 3 | (4) | 1 | (33) |
| Blockley | 3 | (2) | 3 | (100) | 2 | (3) | 2 | (100) |
| Mikawashima | 1 | (1) | 0 | 2 | (3) | 0 | ||
| Litchfield | 1 | (1) | 0 | 2 | (3) | 1 | (50) | |
| Bovismorbificans | 2 | (1) | 0 | 2 | (3) | 0 | ||
| Give | 0 | (0) | 0 | 1 | (1) | 1 | (100) | |
| Havana | 1 | (1) | 1 | (100) | 1 | (1) | 0 | |
| Weltevreden | 0 | (0) | 0 | 1 | (1) | 0 | ||
| Braenderup | 5 | (3) | 1 | (20) | 1 | (1) | 0 | |
| Manhattan | 0 | (0) | 0 | 1 | (1) | 1 | (100) | |
| Schwarzengrund | 17 | (11) | 16 | (94) | 1 | (1) | 1 | (100) |
| Mbandaka | 3 | (2) | 0 | 1 | (1) | 0 | ||
| Colindale | 2 | (1) | 0 | 1 | (1) | 0 | ||
| Narashino | 0 | (0) | 0 | 1 | (1) | 1 | (100) | |
| Senftenberg | 0 | (0) | 0 | 1 | (1) | 0 | ||
| Willemstad | 0 | (0) | 0 | 1 | (1) | 1 | (100) | |
| Untypable (O4:i:-) | 0 | (0) | 0 | 6 | (8) | 6 | (100) | |
| Untypable (O8:-:1,5) | 1 | (1) | 0 | 1 | (1) | 0 | ||
| Untypable (O8:e,h:-) | 0 | (0) | 0 | 1 | (1) | 0 | ||
| Untypable (OUT:r:1,5) | 0 | (0) | 0 | 1 | (1) | 1 | (100) | |
| Others | 75 | (48) | 11 | (15) | 0 | 0 | ||
| Total | 157 | 44 | (28) | 77 | 27 | (35) | ||
| Serotype | Sources | 2016 | 2017 | 2018 | 2019 | Total | ||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 6 | 7 | 8 | 9 | 10 | 11 | 12 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | |||
| Agona | Sewage | - | - | 1 | - | - | - | - | - | 1 | 1 | 1 | - | 1 | - | - | 1 | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 7 |
| Human | 1 | - | - | - | - | - | - | - | - | - | - | 1 | - | 1 | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | 1 | 1 | 7 | |
| Bareilly | Sewage | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | - | 1 | - | - | 1 | - | 1 | 1 | - | - | - | 1 | - | - | - | - | - | 7 |
| Human | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | 1 | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | 1 | - | 4 | |
| Bovismorbificans | Sewage | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 2 | - | 2 |
| Human | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 2 | - | 2 | |
| Brandenburg | Sewage | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | 1 | - | - | - | 2 |
| Human | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | 1 | - | - | - | - | - | 1 | - | 4 | |
| Colindale | Sewage | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 2 | - | - | - | - | - | - | - | - | - | - | - | 2 |
| Human | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - | 1 | |
| Infantis | Sewage | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | 1 | 1 | - | - | - | - | - | - | - | 1 | - | - | - | 1 | - | 1 | - | - | - | - | - | - | 6 |
| Human | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | 1 | 3 | |
| Litchfield | Sewage | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Human | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | 2 | |
| Mbandaka | Sewage | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | 1 | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | 3 |
| Human | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | |
| Newport | Sewage | - | - | 1 | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 2 | 2 | - | - | - | 1 | - | - | - | - | - | - | 1 | - | - | 8 |
| Human | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | 3 | |
| Saintpaul | Sewage | - | - | - | - | - | 1 | - | 1 | - | 1 | - | - | 1 | - | 1 | - | 1 | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 7 |
| Human | - | - | 2 | - | - | - | - | - | - | - | - | - | - | - | 2 | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | 5 | |
| Schwarzengrund | Sewage | - | 1 | - | 1 | - | - | 1 | 1 | - | 1 | 2 | 1 | - | 1 | 1 | - | - | - | - | - | - | - | - | - | - | 1 | - | 1 | - | 1 | 1 | 1 | - | - | 1 | - | 1 | - | 17 |
| Human | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | |
| Stanley | Sewage | - | - | - | 1 | - | - | - | 1 | - | 1 | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 4 |
| Human | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | 1 | 2 | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - | 1 | 6 | |
| Untypable (O8:-:1,5) | Sewage | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Human | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yanagimoto, K.; Yamagami, T.; Uematsu, K.; Haramoto, E. Characterization of Salmonella Isolates from Wastewater Treatment Plant Influents to Estimate Unreported Cases and Infection Sources of Salmonellosis. Pathogens 2020, 9, 52. https://doi.org/10.3390/pathogens9010052
Yanagimoto K, Yamagami T, Uematsu K, Haramoto E. Characterization of Salmonella Isolates from Wastewater Treatment Plant Influents to Estimate Unreported Cases and Infection Sources of Salmonellosis. Pathogens. 2020; 9(1):52. https://doi.org/10.3390/pathogens9010052
Chicago/Turabian StyleYanagimoto, Keita, Takaya Yamagami, Kosei Uematsu, and Eiji Haramoto. 2020. "Characterization of Salmonella Isolates from Wastewater Treatment Plant Influents to Estimate Unreported Cases and Infection Sources of Salmonellosis" Pathogens 9, no. 1: 52. https://doi.org/10.3390/pathogens9010052