
Tick Activity, Host Range, and Tick-Borne Pathogen Prevalence in Mountain Habitats of the Western Carpathians, Poland
 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Materials and Methods
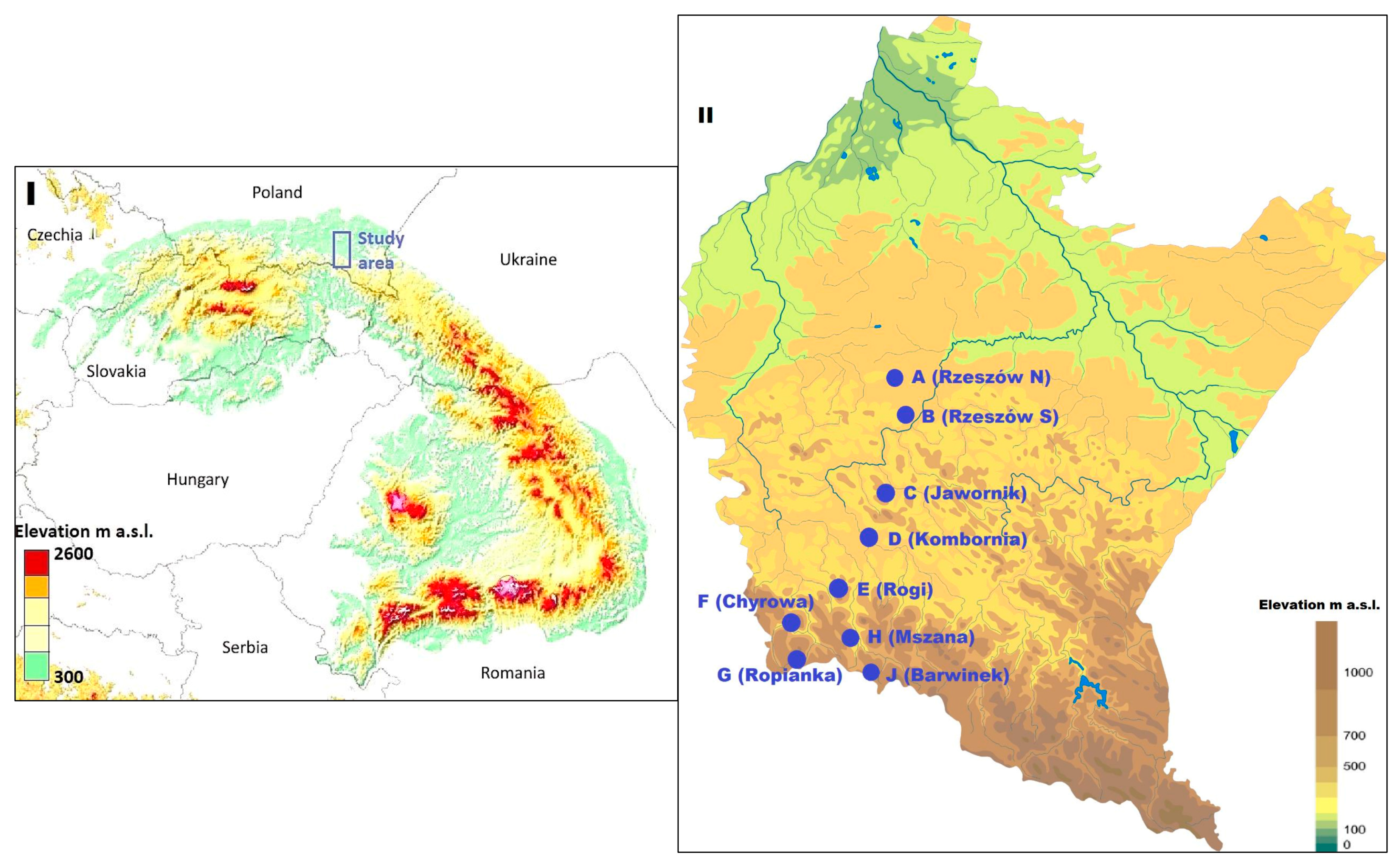
2.1. Study Area
2.2. Occurrence and Seasonal Activity of Ticks
2.3. Circadian Tick Activity
2.4. Host Range
2.5. Blood Collection Protocol
2.6. Molecular Analysis
2.6.1. DNA Extraction
2.6.2. DNA Preamplification
2.6.3. Microfluidic Real-Time PCR for High-Throughput Detection of Microorganisms
2.6.4. Validation of Microfluidic Real-Time PCR and Phylogenetic Analysis
2.7. Statistical Analysis
2.7.1. Prevalence of TBPs in Pooled Samples
2.7.2. Analysis of the Significance of Results
3. Results
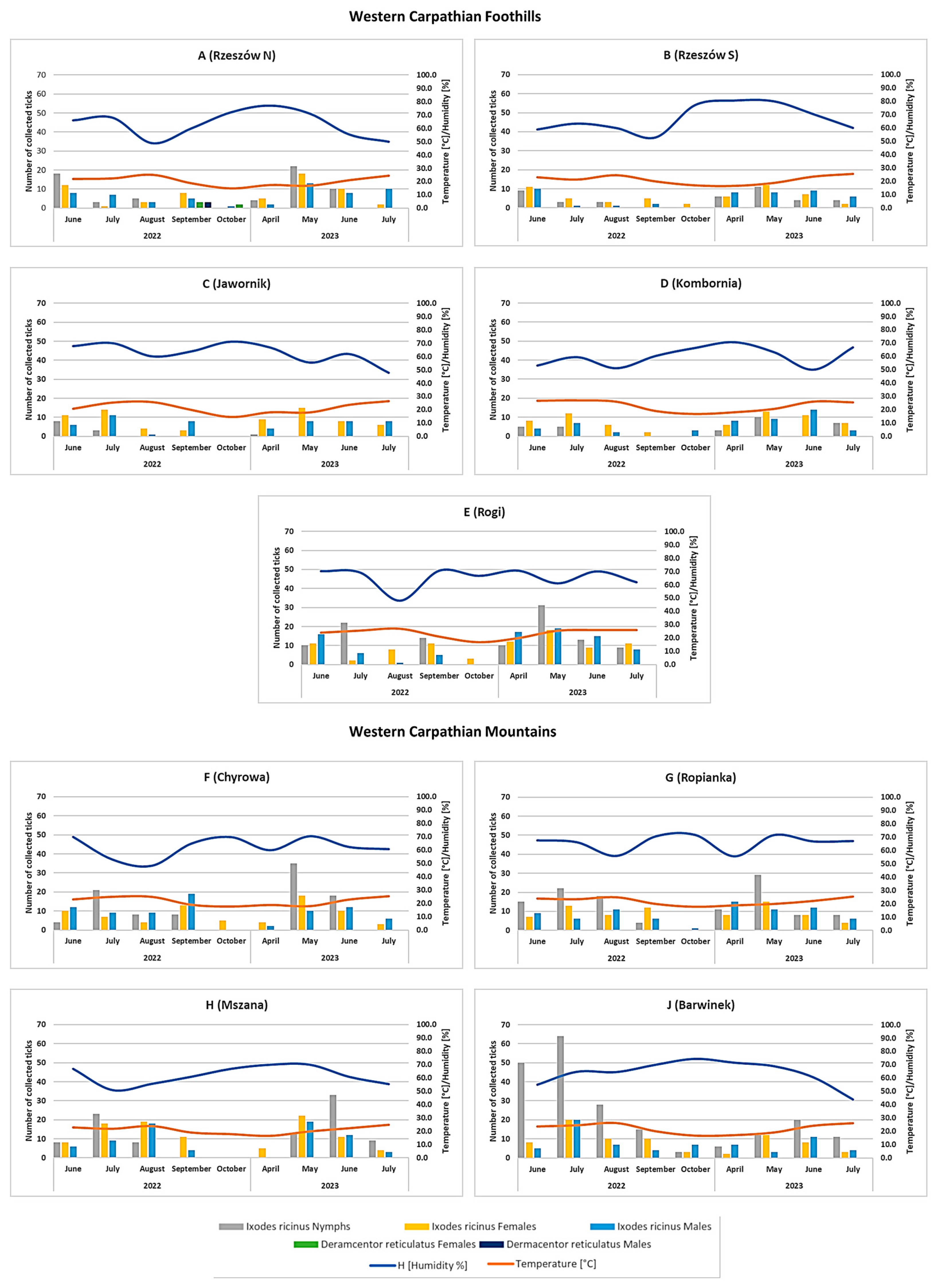
3.1. Occurrence and Seasonal Activity of Ticks
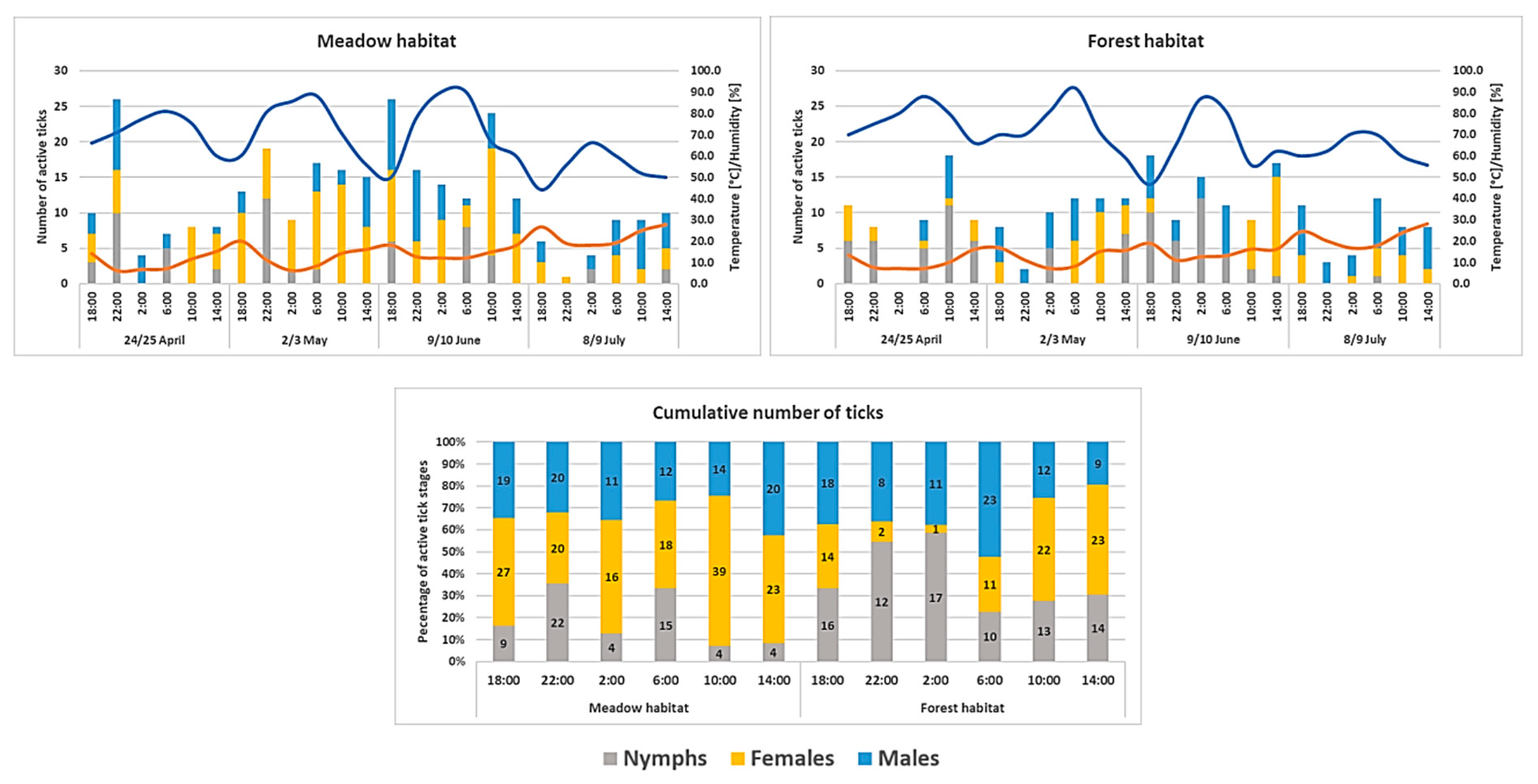
3.2. Circadian Activity of Ixodes ricinus Ticks
3.3. Host Range
3.4. Prevalence of TBPs in Questing Ixodes ricinus Ticks
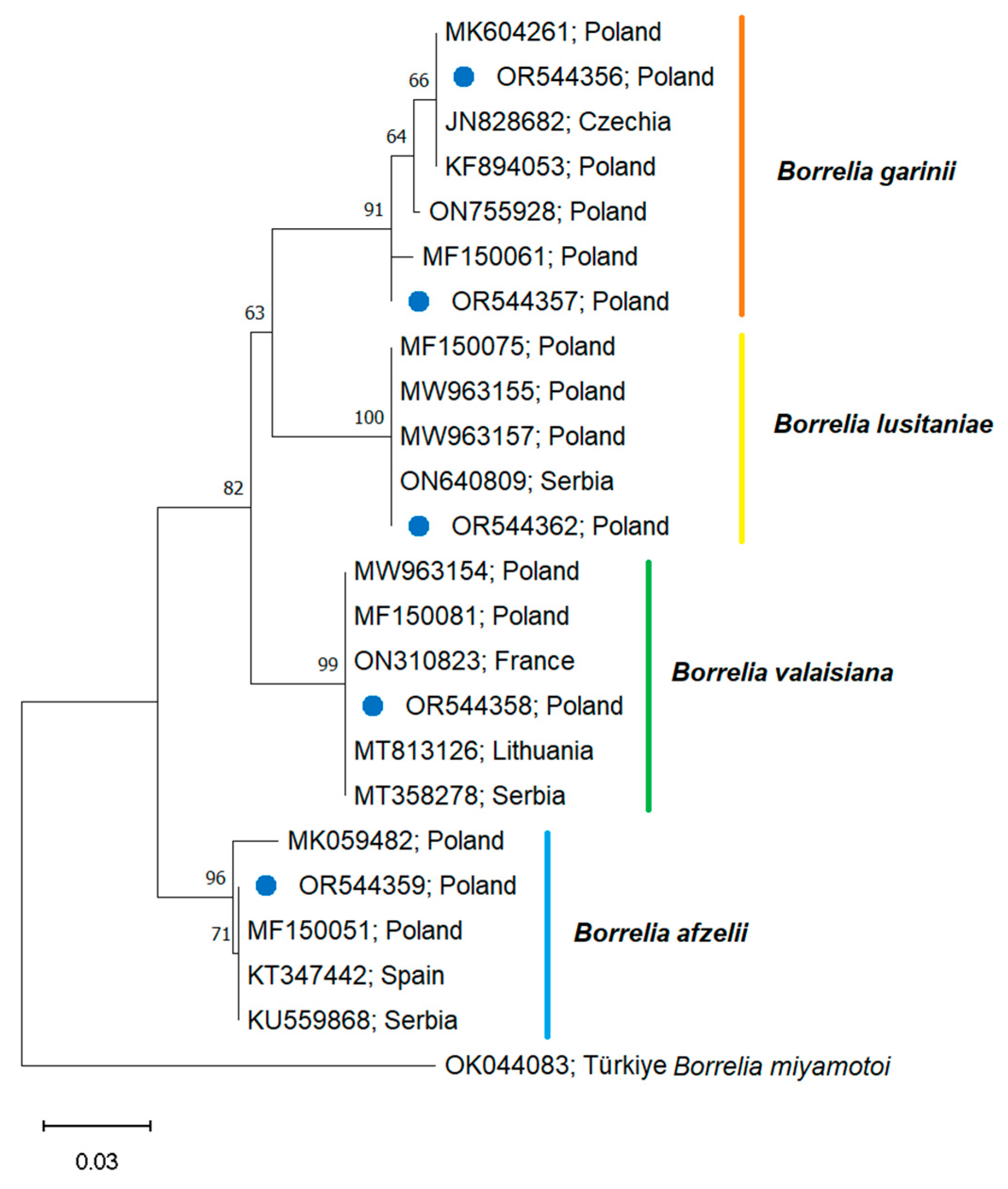
3.4.1. Prevalence and Phylogeny of Spirochetes of the Borreliaceae Family
3.4.2. Prevalence of Pathogens of the Anaplasmataceae Family
3.4.3. Prevalence of Bacteria of the Rickettsiaceae Family
3.4.4. Prevalence of the Protozoan Parasite Babesia venatorum
3.4.5. Coinfection with TBPs in Questing Ixodes ricinus Ticks
3.5. Rodents as a Reservoir of TBPs in Nature
3.5.1. Prevalence of TBPs in Juvenile Ticks Collected from Rodents
3.5.2. Prevalence of TBPs in Rodent Blood Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zachos, F.E.; Habel, J.C. Biodiversity Hotspots; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Estrada-Peña, A.; Cutler, S.; Potkonjak, A.; Vassier-Tussaut, M.; Van Bortel, W.; Zeller, H.; Fernández-Ruiz, N.; Mihalca, A.D. An updated meta-analysis of the distribution and prevalence of Borrelia burgdorferi s.l. in ticks in Europe. Int. J. Health Geograph. 2018, 17, 41. [Google Scholar] [CrossRef]
- Vikse, R.; Paulsen, K.M.; Edgar, K.S.; Pettersson, J.H.-O.; Ottesen, P.S.; Okbaldet, Y.B.; Kiran, N.; Lamsal, A.; Lindstedt, H.E.H.; Pedersen, B.N.; et al. Geographical istribution and prevalence of tick-borne encephalitis virus in questing Ixodes ricinus ticks and phylogeographic structure of the Ixodes ricinus vector in Norway. Zoonoses Pub. Health 2020, 67, 370–381. [Google Scholar] [CrossRef]
- Zając, Z.; Kulisz, J.; Bartosik, K.; Woźniak, A.; Dzierżak, M.; Khan, A. Environmental determinants of the occurrence and activity of Ixodes ricinus ticks and the prevalence of tick-borne diseases in Eastern Poland. Sci. Rep. 2021, 11, 15472. [Google Scholar] [CrossRef]
- Burn, L.; Pilz, A.; Vyse, A.; Gutiérrez Rabá, A.V.; Angulo, F.J.; Tran, T.M.P.; Fletcher, M.A.; Gessner, B.D.; Moïsi, J.C.; Stark, J.H. Seroprevalence of Lyme borreliosis in Europe: Results from a systematic literature review (2005–2020). Vector Borne Zoonot. Dis. 2023, 23, 195–220. [Google Scholar] [CrossRef]
- Rubel, F.; Brugger, K.; Pfeffer, M.; Chitimia-Dobler, L.; Didyk, Y.M.; Leverenz, S.; Dautel, H.; Kahl, O. Geographical distribution of Dermacentor marginatus and Dermacentor reticulatus in Europe. Ticks Tick. Dis. 2016, 7, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Bilbija, B.; Spitzweg, C.; Papoušek, I.; Fritz, U.; Földvári, G.; Mullett, M.; Ihlow, F.; Sprong, H.; Civáňová Křížová, K.; Anisimov, N.; et al. Dermacentor reticulatus—A tick on its way from glacial refugia to a panmictic eurasian population. Int. J. Parasitol. 2023, 53, 91–101. [Google Scholar] [CrossRef]
- Zając, Z.; Obregon, D.; Foucault-Simonin, A.; Wu-Chuang, A.; Moutailler, S.; Galon, C.; Kulisz, J.; Woźniak, A.; Bartosik, K.; Cabezas-Cruz, A. Disparate dynamics of pathogen prevalence in Ixodes ricinus and Dermacentor reticulatus ticks occurring sympatrically in diverse habitats. Sci. Rep. 2023, 13, 10645. [Google Scholar] [CrossRef]
- Banović, P.; Díaz-Sánchez, A.A.; Foucault-Simonin, A.; Mateos-Hernandez, L.; Wu-Chuang, A.; Galon, C.; Simin, V.; Mijatović, D.; Bogdan, I.; Corona-González, B.; et al. Emerging tick-borne spotted fever group rickettsioses in the Balkans. Infect. Gen. Evol. 2023, 107, 105400. [Google Scholar] [CrossRef] [PubMed]
- Ličková, M.; Fumačová Havlíková, S.; Sláviková, M.; Slovák, M.; Drexler, J.F.; Klempa, B. Dermacentor reticulatus is a vector of tick-borne Encephalitis Virus. Ticks Tick. Dis. 2020, 11, 101414. [Google Scholar] [CrossRef]
- Földvári, G.; Široký, P.; Szekeres, S.; Majoros, G.; Sprong, H. Dermacentor reticulatus: A vector on the rise. Parasit. Vect. 2016, 9, 314. [Google Scholar] [CrossRef]
- Ionita, M.; Mitrea, I.L.; Pfister, K.; Hamel, D.; Silaghi, C. Molecular evidence for bacterial and protozoan pathogens in hard ticks from Romania. Vet. Parasitol. 2013, 196, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Peña, A.; Mihalca, A.D.; Petney, T.N. Ticks of Europe and North Africa; Springer: Cham, Switzerland, 2018. [Google Scholar]
- Nowak-Chmura, M.; Siuda, K. Ticks of Poland. Review of contemporary issues and latest research. Ann. Parasitol. 2012, 58, 125–155. [Google Scholar] [PubMed]
- Dwużnik-Szarek, D.; Mierzejewska, E.J.; Rodo, A.; Goździk, K.; Behnke-Borowczyk, J.; Kiewra, D.; Kartawik, N.; Bajer, A. Monitoring the expansion of Dermacentor reticulatus and occurrence of canine babesiosis in Poland in 2016–2018. Parasit. Vect. 2021, 14, 267. [Google Scholar] [CrossRef]
- Siuda, K. Kleszcze Polski (Acari: Ixodida): Systematyka i Rozmieszczenie; Polskie Towarzystwo Parazytologiczne: Kraków, Poland, 1993. [Google Scholar]
- Kocoń, A.; Asman, M.; Nowak-Chmura, M.; Witecka, J.; Kłyś, M.; Solarz, K. Molecular detection of tick-borne pathogens in ticks collected from pets in selected muntainous areas of Tatra County (Tatra Mountains, Poland). Sci. Rep. 2020, 10, 15865. [Google Scholar] [CrossRef]
- Akimov, I.A.; Nebogatkin, I.V. Distribution of ticks from the Genus Dermacentor (Acari, Ixodidae) in Ukraine. Vestn. Zool. 2011, 45, 35–40. [Google Scholar] [CrossRef]
- Stanko, M.; Derdáková, M.; Špitalská, E.; Kazimírová, M. Ticks and their epidemiological role in Slovakia: From the past till present. Biologia 2021, 77, 1575–1610. [Google Scholar] [CrossRef] [PubMed]
- Rączkowska, Z. The Carpathians. In Periglacial Landscapes of Europe; Springer International Publishing: Cham, Switzerland, 2023. [Google Scholar]
- Available online: https://meteomodel.pl/dane/srednie-miesieczne/?imgwid=249210240&par=tm&max_empty=2 (accessed on 15 June 2023).
- Matuszkiewicz, J.M.; Wolski, J. Potencjalna Roślinność Naturalna Polski; IGiPZ PAN: Warszw, Poland, 2023; Available online: https://www.igipz.pan.pl/Roslinnosc-potencjalna-zgik.html (accessed on 15 June 2023).
- Skuratowicz, W. Pchły-Siphonaptera (Aphaniptera). In Klucze do Oznaczania Owadów Polski; Państwowe Wydawnictwo Naukowe: Warsaw, Poland, 1967. [Google Scholar]
- Kowalski, K. Klucz do Oznaczania Ssaków Polski; Państwowe Wydawnictwo Naukowe: Warsaw, Poland, 1984. [Google Scholar]
- Michelet, L.; Delannoy, S.; Devillers, E.; Umhang, G.; Aspan, A.; Juremalm, M.; Chirico, J.; van der Wal, F.J.; Sprong, H.; Boye Pihl, T.P.; et al. High-throughput screening of tick-borne pathogens in Europe. Front. Cell. Inf. Microbiol. 2014, 4, 103. [Google Scholar] [CrossRef]
- Young, L.; Sung, J.; Stacey, G.; Masters, J.R. Detection of Mycoplasma in cell cultures. Nature Protocol. 2010, 5, 929–934. [Google Scholar] [CrossRef] [PubMed]
- Karbowiak, G. Changes in the occurrence range of hosts cause the expansion of the ornate dog tick Dermacentor reticulatus (Fabricius, 1794) in Poland. Biologia 2021, 77, 1513–1522. [Google Scholar] [CrossRef]
- Zając, Z.; Kulisz, J.; Woźniak, A.; Bartosik, K.; Khan, A. Seasonal activity of Dermacentor reticulatus ticks in the era of progressive climate change in Eastern Poland. Sci. Rep. 2021, 11, 20382. [Google Scholar] [CrossRef] [PubMed]
- Dwużnik-Szarek, D.; Mierzejewska, E.J.; Bajer, A. Occurrence of juvenile Dermacentor reticulatus ticks in three regions in Poland: The final evidence of the conquest. Parasit. Vect. 2021, 14, 536. [Google Scholar] [CrossRef]
- Kahl, O.; Gray, J.S. The biology of Ixodes ricinus with emphasis on its ecology. Ticks Tick. Dis. 2023, 14, 102114. [Google Scholar] [CrossRef]
- Medlock, J.M.; Hansford, K.M.; Bormane, A.; Derdakova, M.; Estrada-Peña, A.; George, J.-C.; Golovljova, I.; Jaenson, T.G.T.; Jensen, J.-K.; Jensen, P.M.; et al. Driving forces for changes in geographical distribution of Ixodes ricinus ticks in Europe. Parasit. Vect. 2013, 6, 1. [Google Scholar] [CrossRef]
- Mancini, F.; Di Luca, M.; Toma, L.; Vescio, F.; Bianchi, R.; Khoury, C.; Marini, L.; Rezza, G.; Ciervo, A. Prevalence of tick-borne pathogens in an urban park in Rome, Italy. Ann. Agric. Environ. Med. 2014, 21, 723–727. [Google Scholar] [CrossRef]
- Halos, L.; Bord, S.; Cotté, V.; Gasqui, P.; Abrial, D.; Barnouin, J.; Boulouis, H.-J.; Vayssier-Taussat, M.; Vourc’h, G. Ecological factors characterizing the prevalence of bacterial tick-borne pathogens in Ixodes ricinus ticks in pastures and woodlands. App. Environ. Microbiol. 2010, 76, 4413–4420. [Google Scholar] [CrossRef] [PubMed]
- Randolph, S.E. Tick ecology: Processes and patterns behind the epidemiological risk posed by Ixodid ticks as vectors. Parasitology 2004, 129, S37–S65. [Google Scholar] [CrossRef]
- Kiewra, D.; Sobczyński, M. Biometrical analysis of the common tick, Ixodes ricinus, in the Ślęża Massif (Lower Silesia, Poland). J. Vector Ecol. 2006, 31, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Pietzsch, M.E.; Rice, N.V.P.; Jones, L.; Kerrod, E.; Avenell, D.; Los, S.; Ratcliffe, N.; Leach, S.; Butt, T. Investigation of ecological and environmental determinants for the presence of questing Ixodes ricinus (Acari: Ixodidae) on Gower, South Wales. J. Med. Entomol. 2008, 45, 314–325. [Google Scholar] [CrossRef] [PubMed]
- Lees, A.D. The water balance in Ixodes ricinus L. and certain other species of ticks. Parasitology 1946, 37, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.; Kahl, O.; Zintl, A. What do we still need to know about Ixodes ricinus? Ticks Tick. Dis. 2021, 12, 101682. [Google Scholar] [CrossRef]
- Materna, J.; Daniel, M.; Metelka, L.; Harčarik, J. The vertical distribution, density and the development of the tick Ixodes ricinus in mountain areas influenced by climate changes (The Krkonoše Mts., Czech Republic). Int. J. Med. Microbiol. 2008, 298, 25–37. [Google Scholar] [CrossRef]
- Garcia-Vozmediano, A.; Krawczyk, A.I.; Sprong, H.; Rossi, L.; Ramassa, E.; Tomassone, L. Ticks climb the mountains: Ixodid tick infestation and infection by tick-borne pathogens in the Western Alps. Ticks Tick. Dis. 2020, 11, 101489. [Google Scholar] [CrossRef]
- Danielová, V.; Rudenko, N.; Daniel, M.; Holubová, J.; Materna, J.; Golovchenko, M.; Schwarzová, L. Extension of Ixodes ricinus ticks and agents of tick-borne diseases to mountain areas in the Czech Republic. Int. J. Med. Microbiol. 2006, 296, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Rosický, B. Notes on ecology of the tick Ixodes ricinus L. in Central Europe with respect to natural foci. Věst. Českoslov. Spol. Zool. 1954, 18, 41–70. [Google Scholar]
- Černý, V.; Rosický, B.; Ašmera, J.; Kadlčík, K.; Kobík, V.; Kratochvílová, E.; Lauterer, P.; Novák, D.; Pauleová, E.; Šebek, Z.; et al. Results of investigations of phenology of the common tick Ixodes ricinus (L.) in the Czech lands in the years 1960–1962. Čs. Parasitol. 1965, 12, 125–131. [Google Scholar]
- Gray, J.S. Ixodes ricinus seasonal activity: Implications of global warming indicated by revisiting tick and weather data. Int. J. Med. Microbiol. 2008, 298, 19–24. [Google Scholar] [CrossRef]
- Schulz, M.; Mahling, M.; Pfister, K. Abundance and seasonal activity of questing Ixodes ricinus ticks in their natural habitats in Southern Germany in 2011. J. Vec. Ecol. 2014, 39, 56–65. [Google Scholar] [CrossRef]
- Stafford, K.C., III. Survival of immature Ixodes scapularis (Acari: Ixodidae) at different relative humidities. J. Med. Entomol. 1994, 31, 310–314. [Google Scholar] [CrossRef]
- Perret, J.-L.; Guerin, P.M.; Diehl, P.A.; Vlimant, M.; Gern, L. Darkness induces mobility, and saturation deficit limits questing duration, in the tick Ixodes ricinus. J. Exp. Biol. 2003, 206, 1809–1815. [Google Scholar] [CrossRef]
- Mihalca, A.D.; Sándor, A.D. The role of rodents in the ecology of Ixodes ricinus and associated pathogens in Central and Eastern Europe. Front. Cell. Infec. Microbiol. 2013, 3, 56. [Google Scholar] [CrossRef]
- Khanal, S.; Taank, V.; Anderson, J.F.; Sultana, H.; Neelakanta, G. Rickettsial pathogen perturbs tick circadian gene to infect the vertebrate host. Int. J. Mol. Sci. 2022, 23, 3545. [Google Scholar] [CrossRef] [PubMed]
- Opalińska, P.; Wierzbicka, A.; Asman, M.; Rączka, G.; Dyderski, M.K.; Nowak-Chmura, M. Fivefold higher abundance of ticks (Acari: Ixodida) on the European Roe Deer (Capreolus capreolus L.) forest than field ecotypes. Sci. Rep. 2021, 11, 10649. [Google Scholar] [CrossRef] [PubMed]
- Vor, T.; Kiffner, C.; Hagedorn, P.; Niedrig, M.; Rühe, F. Tick burden on European Roe Deer (Capreolus Capreolus). Exp. App. Acarol. 2010, 51, 405–417. [Google Scholar] [CrossRef]
- Takumi, K.; Hofmeester, T.R.; Sprong, H. Red and Fallow Deer determine the density of Ixodes ricinus nymphs containing Anaplasma phagocytophilum. Parasit. Vect. 2021, 14, 59. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, L.; Maffey, G.L.; Ramsay, S.L.; Hester, A.J. The effect of deer management on the abundance of Ixodes ricinusin Scotland. Ecol. Appl. 2012, 22, 658–667. [Google Scholar] [CrossRef]
- Krcmar, S. Hard ticks (Acari, Ixodidae) of Croatia. Zoo Keys 2012, 234, 19–57. [Google Scholar] [CrossRef] [PubMed]
- Steigedal, H.H.; Loe, L.E.; Grøva, L.; Mysterud, A. The effect of sheep (Ovis aries) presence on the abundance of ticks (Ixodes ricinus). Acta Agric. Scand. Sect. A-Animal Sci. 2013, 63, 111–120. [Google Scholar] [CrossRef]
- Karbowiak, G.; Miklisová, D.; Stanko, M.; Werszko, J.; Hajdul-Marwicz, M.; Szewczyk, T.; Rychlik, L. The competition between immatures of Ixodes ricinus and Dermacentor reticulatus (Ixodida: Ixodidae) ticks for rodent hosts. J. Med. Entomol. 2018, 56, 448–452. [Google Scholar] [CrossRef]
- Cayol, C.; Koskela, E.; Mappes, T.; Siukkola, A.; Kallio, E.R. Temporal dynamics of the tick Ixodes ricinus in Northern Europe: Epidemiological implications. Parasit. Vect. 2017, 10, 166. [Google Scholar] [CrossRef]
- Paulauskas, A.; Radzijevskaja, J.; Rosef, O.; Turcinaviciene, J.; Ambrasiene, D. Infestation of mice and voles with Ixodes ricinus ticks in Lithuania and Norway. Estonian J. Ecol. 2009, 58, 112. [Google Scholar] [CrossRef]
- Sawczyn-Domańska, A.; Zwoliński, J.; Kloc, A.; Wójcik-Fatla, A. Prevalence of Borrelia, Neoehrlichia Mikurensis and Babesia in ticks collected from vegetation in Eastern Poland. Exp. App. Acarol. 2023, 90, 409–428. [Google Scholar] [CrossRef]
- Strnad, M.; Hönig, V.; Růžek, D.; Grubhoffer, L.; Rego, R.O.M. Europe-wide meta-analysis of Borrelia burgdorferi sensu lato prevalence in questing Ixodes ricinus ticks. App. Environ. Microbiol. 2017, 83, e00609-17. [Google Scholar] [CrossRef] [PubMed]
- Severinsson, K.; Jaenson, T.G.; Pettersson, J.; Falk, K.; Nilsson, K. Detection and prevalence of Anaplasma phagocytophilum and Rickettsia helvetica in Ixodes ricinus ticks in seven study areas in Sweden. Parasit. Vect. 2010, 3, 66. [Google Scholar] [CrossRef]
- Klitgaard, K.; Kjær, L.J.; Isbrand, A.; Hansen, M.F.; Bødker, R. Multiple infections in questing nymphs and adult female Ixodes ricinus ticks collected in a recreational forest in Denmark. Ticks Tick. Dis. 2019, 10, 1060–1065. [Google Scholar] [CrossRef]
- Sprong, H.; Wielinga, P.R.; Fonville, M.; Reusken, C.; Brandenburg, A.H.; Borgsteede, F.; Gaasenbeek, C.; van der Giessen, J.W. Ixodes ricinus ticks are reservoir hosts for Rickettsia helvetica and potentially carry Flea-borne Rickettsia species. Parasit. Vect. 2009, 2, 41. [Google Scholar] [CrossRef] [PubMed]
- Christova, I.; Schouls, L.; van de Pol, I.; Park, J.; Panayotov, S.; Lefterova, V.; Kantardjiev, T.; Dumler, J.S. High prevalence of granulocytic Ehrlichiae and Borrelia burgdorferi sensu lato in Ixodes ricinus ticks from Bulgaria. J. Clin. Microbiol. 2001, 39, 4172–4174. [Google Scholar] [CrossRef]
- Egyed, L.; Élő, P.; Sréter-Lancz, Z.; Széll, Z.; Balogh, Z.; Sréter, T. Seasonal activity and tick-borne pathogen infection rates of Ixodes ricinus ticks in Hungary. Ticks Tick. Dis. 2012, 3, 90–94. [Google Scholar] [CrossRef]
- Matei, I.A.; Kalmár, Z.; Magdaş, C.; Magdaş, V.; Toriay, H.; Dumitrache, M.O.; Ionică, A.M.; D’Amico, G.; Sándor, A.D.; Mărcuţan, D.I.; et al. Anaplasma phagocytophilum in questing Ixodes ricinus ticks from Romania. Ticks Tick. Dis. 2015, 6, 408–413. [Google Scholar] [CrossRef]
- Diuk-Wasser, M.A.; Vannier, E.; Krause, P.J. Coinfection by Ixodes tick-borne pathogens: Ecological, epidemiological, and clinical consequences. Trends Parasitol. 2016, 32, 30–42. [Google Scholar] [CrossRef]
- Hakimi, H.; Sarani, A.; Takeda, M.; Kaneko, O.; Asada, M. Epidemiology, risk factors, and co-infection of vector-borne pathogens in goats from Sistan and Baluchestan province, Iran. PLoS ONE 2019, 14, e0218609. [Google Scholar] [CrossRef] [PubMed]
- Lommano, E.; Bertaiola, L.; Dupasquier, C.; Gern, L. Infections and coinfections of questing Ixodes ricinus ticks by emerging zoonotic pathogens in Western Switzerland. App. Environ. Microbiol. 2012, 78, 4606–4612. [Google Scholar] [CrossRef]
- Milutinović, M.; Masuzawa, T.; Tomanović, S.; Radulović, Ž.; Fukui, T.; Okamoto, Y. Borrelia burgdorferi sensu lato, Anaplasma phagocytophilum, Francisella tularensis and their co-infections in host-seeking Ixodes ricinus ticks collected in Serbia. Exp. Appl. Acarol. 2008, 45, 171–183. [Google Scholar] [CrossRef]
- Krawczyk, A.I.; van Duijvendijk, G.L.A.; Swart, A.; Heylen, D.; Jaarsma, R.I.; Jacobs, F.H.H.; Fonville, M.; Sprong, H.; Takken, W. Effect of rodent density on tick and tick-borne pathogen populations: Consequences for infectious disease risk. Parasit. Vect. 2020, 13, 34. [Google Scholar] [CrossRef] [PubMed]
- Burri, C.; Dupasquier, C.; Bastic, V.; Gern, L. Pathogens of emerging tick-borne diseases, Anaplasma phagocytophilum, Rickettsia spp., and Babesia spp., in Ixodes ticks collected from rodents at four sites in Switzerland (Canton of Bern). Vec. Borne Zoo. Dis. 2011, 11, 939–944. [Google Scholar] [CrossRef] [PubMed]
- Brandenburg, P.J.; Obiegala, A.; Schmuck, H.M.; Dobler, G.; Chitimia-Dobler, L.; Pfeffer, M. Seroprevalence of tick-borne encephalitis (TBE) virus antibodies in wild rodents from two natural TBE foci in Bavaria, Germany. Pathogens 2023, 12, 185. [Google Scholar] [CrossRef] [PubMed]
- Burri, C.; Schumann, O.; Schumann, C.; Gern, L. Are Apodemus spp. mice and Myodes glareolus reservoirs for Borrelia miyamotoi, Candidatus Neoehrlichia mikurensis, Rickettsia helvetica, R. monacensis and Anaplasma phagocytophilum? Ticks Tick. Dis. 2014, 5, 245–251. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Habitat Type | Host Species | Number of Tested Animals | Number of Infested Animals | Tick Species/Number of Collected Ticks | Prevalence (%) | Mean Tick Infestation per Animal | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ixodes ricinus | Dermacentor reticulatus | ||||||||||||
| L | N | F | M | L | N | F | M | ||||||
| Forest | Myodes glareolus | 18 | 16 | 30 | 49 | 0 | 0 | 0 | 0 | 0 | 0 | 88.9 | 4.9 |
| Apodemus flavicollis | 3 | 1 | 0 | 0 | 0 | 0 | 0 | 6 | 0 | 0 | 33.3 | 6.0 | |
| Microtus spp. | 2 | 1 | 0 | 6 | 0 | 0 | 0 | 0 | 0 | 0 | 50.0 | 6.0 | |
| Meadow | Apodemus agrarius | 24 | 14 | 6 | 36 | 0 | 0 | 0 | 6 | 0 | 0 | 58.3 | 3.4 |
| Microtus spp. | 2 | 1 | 0 | 0 | 0 | 0 | 0 | 7 | 0 | 0 | 50.0 | 7.0 | |
| Open range | Capreolus capreolus | 7 | 7 | 0 | 0 | 43 | 5 | 0 | 0 | 0 | 5 | 100.0 | 7.5 |
| Pasture | Ovis aries | 75 | 5 | 0 | 0 | 17 | 0 | 0 | 0 | 0 | 0 | 6.7 | 3.4 |
| Tick Collection Sites | Tick Stages | Tick-Borne Pathogens/Number of Positive Samples and Percentage Rate (%) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Borrelia spp. | Borrelia burgdorferi s.s | Borrelia garinii | Borrelia afzelii | Borrelia valaisiana | Borrelia lusitaniae | Borrelia spielmanii | Borrelia miyamotoi | Anaplasma phagocytophilum | Ehrlichia spp. | Rickettsia spp. | Rickettsia helvetica | Babesia venatorum | ||
| Meadow habitat of the Western Carpathians | Females n = 42 | 4 (9.52) | 1 (2.38) | 9 (21.42) | 4 (9.52) | 2 (4.76) | 0 (0.00) | 1 (2.38) | 1 (2.38) | 0 (0.00) | 3 (7.14) | 0 (0.00) | 4 (9.52) | 0 (0.00) |
| Males n = 14 | 1 (7.14) | 0 (0.00) | 1 (7.14) | 4 (28.57) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 1 (7.14) | 0 (0.00) | 1 (7.14) | 0 (0.00) | 4 (28.57) | 0 (0.00) | |
| Nymphs pn = 4 | 0 (0.00) | 0 (0.00) | 0 (0.00) | 1 * (4.17) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 1 * (4.17) | 0 (0.00) | |
| Forest habitat of the Western Carpathians | Females n = 42 | 1 (2.38) | 0 (0.00) | 6 (14.29) | 2 (4.76) | 0 (0.00) | 4 (9.52) | 0 (0.00) | 0 (0.00) | 1 (2.38) | 2 (4.76) | 0 (0.00) | 9 (21.43) | 1 (2.38) |
| Males n = 14 | 1 (7.14) | 0 (0.00) | 1 (7.14) | 1 (7.14) | 0 (0.00) | 1 (7.14) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 1 (7.14) | 1 (7.14) | 0 (0.00) | 0 (0.00) | |
| Nymphs pn = 4 | 0 (0.00) | 0 (0.00) | 1 * (4.17) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 0 (0.00) | |
| Forest habitat of the foothills of the Western Carpathians | Females n = 42 | 1 (2.38) | 2 (4.76) | 7 (16.67) | 5 (11.90) | 1 (2.38) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 4 (9.52) | 0 (0.00) | 10 (23.81) | 1 (2.38) |
| Males n = 14 | 0 (0.00) | 0 (0.00) | 1 (7.14) | 3 (21.43) | 0 (0.00) | 0 (0.00) | 2 (14.29) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 5 (35.71) | 0 (0.00) | |
| Nymphs pn = 4 | 0 (0.00) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 0 (0.00) | |
| Habitat Type | Rodent Species | Tick Species | Tick Stage | Tick-Borne Pathogens/Minimum Infection Rate (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Borrelia spp. | Borrelia afzelii | Borrelia miyamotoi | Rickettsia spp. | Rickettsia helvetica | Bartonella spp. | Mycoplasma spp. | ||||
| Forest | Myodes glareolus | Ixodes ricinus | Larvae n = 5 | 0.00 | 0.00 | 0.00 | 10.00 | 0.00 | 10.00 | 0.00 |
| Nymphs n = 8 | 0.00 | 0.00 | 2.08 | 4.17 | 4.17 | 2.08 | 6.25 | |||
| Microtus spp. | Ixodes ricinus | Nymphs n = 1 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 16.67 | |
| Apodemus flavicollis | Dermacentor reticulatus | Nymphs n = 1 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Meadow | Apodemus agrarius | Ixodes ricinus | Larvae n = 1 | 0.00 | 0.00 | 0.00 | 0.00 | 16.67 | 0.00 | 0.00 |
| Nymphs n = 10 | 3.33 | 1.67 | 0.00 | 6.67 | 3.33 | 1.67 | 8.33 | |||
| Dermacentor reticulatus | Nymphs n = 1 | 0.00 | 0.00 | 0.00 | 0.00 | 16.67 | 0.00 | 0.00 | ||
| Microtus spp. | Dermacentor reticulatus | Nymphs n = 1 | 0.00 | 0.00 | 0.00 | 16.67 | 0.00 | 0.00 | 0.00 | |
| Vector-Borne Pathogens | Forest Biotope | Meadow Biotope |
|---|---|---|
| Myodes glareolus n = 16 | Apodemus agrarius n = 14 | |
| Number of Positive Samples and Percentage Rate (%) | ||
| Bartonella spp. | 4 (25.00) | 0 (0.00) |
| Mycoplasma spp. | 6 (37.50) | 7 (50.00) |
| Borrelia spp. + Mycoplasma spp. | 1 (6.25) | 0 (0.00) |
| Bartonella spp. + Mycoplasma spp. | 3 (18.75) | 2 (14.29) |
| Ehrlichia spp. + Mycoplasma spp. | 0 (0.00) | 4 (28.57) |
| Ehrlichia spp. + Bartonella spp. + Mycoplasma spp. | 0 (0.00) | 1 (7.14) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zając, Z.; Kulisz, J.; Woźniak, A.; Bartosik, K.; Foucault-Simonin, A.; Moutailler, S.; Cabezas-Cruz, A. Tick Activity, Host Range, and Tick-Borne Pathogen Prevalence in Mountain Habitats of the Western Carpathians, Poland. Pathogens 2023, 12, 1186. https://doi.org/10.3390/pathogens12091186
Zając Z, Kulisz J, Woźniak A, Bartosik K, Foucault-Simonin A, Moutailler S, Cabezas-Cruz A. Tick Activity, Host Range, and Tick-Borne Pathogen Prevalence in Mountain Habitats of the Western Carpathians, Poland. Pathogens. 2023; 12(9):1186. https://doi.org/10.3390/pathogens12091186
Chicago/Turabian StyleZając, Zbigniew, Joanna Kulisz, Aneta Woźniak, Katarzyna Bartosik, Angélique Foucault-Simonin, Sara Moutailler, and Alejandro Cabezas-Cruz. 2023. "Tick Activity, Host Range, and Tick-Borne Pathogen Prevalence in Mountain Habitats of the Western Carpathians, Poland" Pathogens 12, no. 9: 1186. https://doi.org/10.3390/pathogens12091186