

Ornithodoros sonrai Soft Ticks and Associated Bacteria in Senegal
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
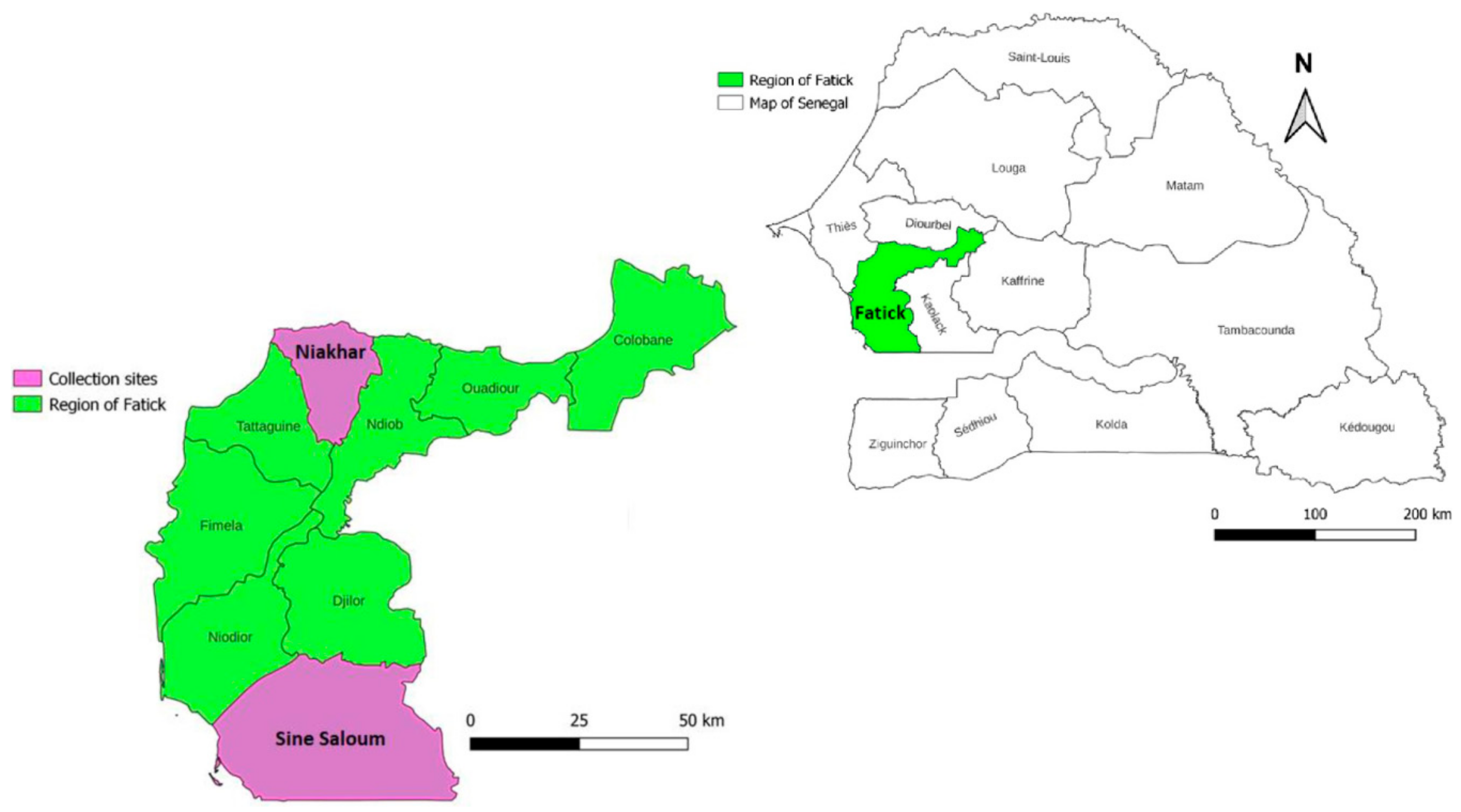
2.1. Study Sites and Tick Collection Periods
2.2. Morphological Identification of Ticks, Dissection and Sample Preparation
2.3. DNA Extraction and Molecular Identification of Ticks
2.4. Sample Homogenisation for MALDI-TOF MS Analyses
2.5. MALDI-TOF Mass Spectrometry Parameters
2.6. MALDI-TOF Spectral Analysis, Database Creation
2.7. MALDI-TOF Blind Test Identification
2.8. Molecular Detection of Tick-Associated Bacteria
2.9. Standard PCR and Phylogenetic Analyses of Tick-Associated Bacteria
2.10. Statistical Analysis
3. Results
3.1. Tick Collection and Morphological Identification
3.2. Molecular Identification of Ticks
3.3. MALDI-TOF MS Identification of O. sonrai Ticks
3.4. Molecular Detection of Micro-Organisms and Identification of Co-Infections
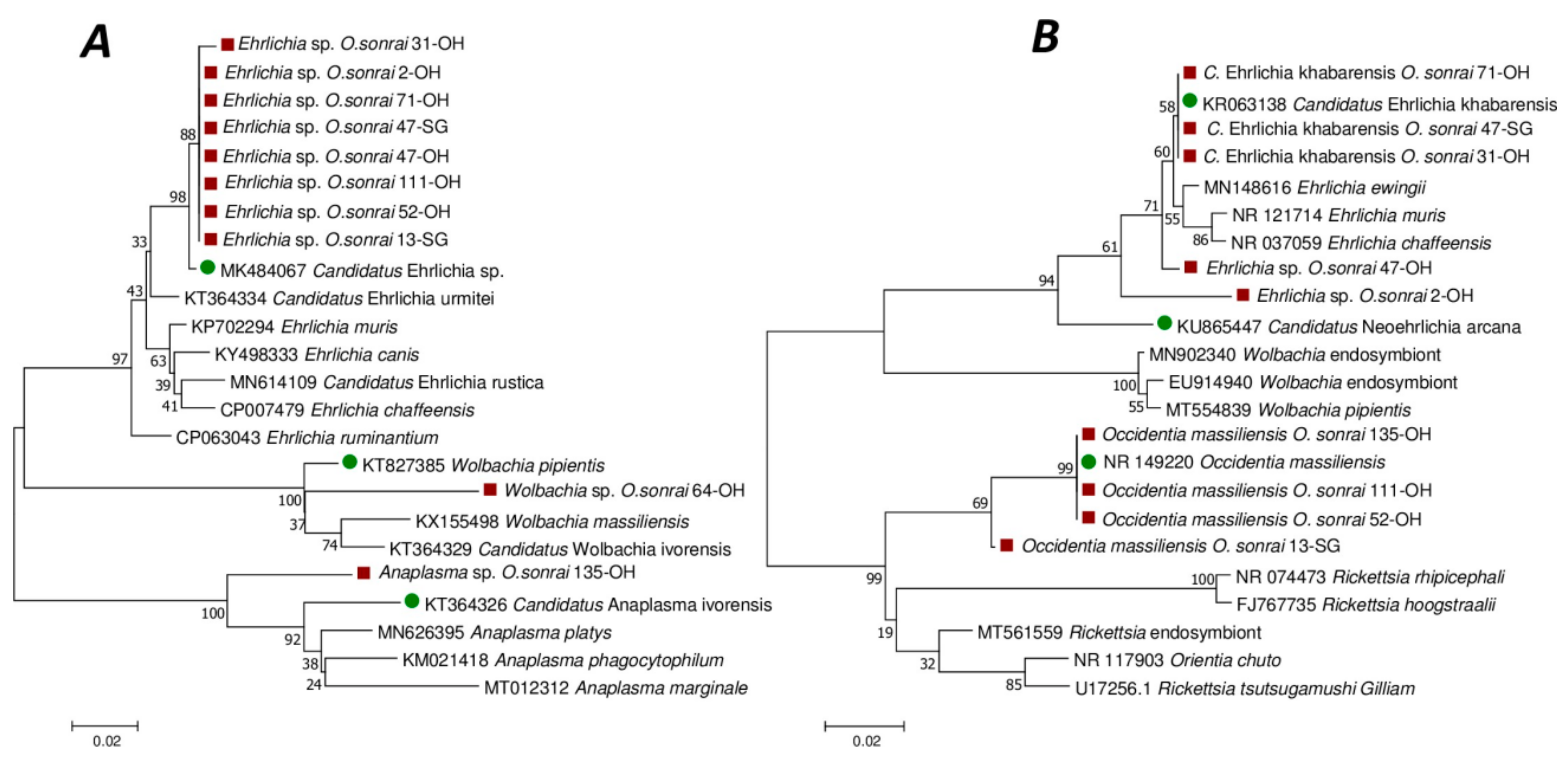
3.5. Standard PCR and Phylogenetic Analyses of Micro-Organisms
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Socolovschi, C.; Doudier, B.; Pages, F.; Parola, P. Ticks and human tick-borne diseases in Africa. Med. Trop. 2008, 68, 119–133. [Google Scholar]
- Parola, P.; Raoult, D. Ticks and Tickborne Bacterial Diseases in Humans: An Emerging Infectious Threat. Clin. Infect. Dis. 2001, 32, 897–928. [Google Scholar] [CrossRef] [PubMed]
- Oliver, J.H. Biology and Systematics of Ticks (Acari:Ixodida). Annu. Rev. Ecol. Syst. 1989, 20, 397–430. [Google Scholar] [CrossRef]
- Guglielmone, A.A.; Robbins, R.G.; Apanaskevich, D.A.; Petney, T.N.; Estrada-Peña, A.; Horak, I.G.; Shao, R.; Barker, S.C. The Argasidae, Ixodidae and Nuttalliellidae (Acari: Ixodida) of the World: A List of Valid Species Names. Zootaxa 2010, 2528, 1–28. [Google Scholar] [CrossRef]
- Trape, J.-F.; Diatta, G.; Arnathau, C.; Bitam, I.; Sarih, M.; Belghyti, D.; Bouattour, A.; Elguero, E.; Vial, L.; Mané, Y.; et al. The Epidemiology and Geographic Distribution of Relapsing Fever Borreliosis in West and North Africa, with a Review of the Ornithodoros Erraticus Complex (Acari: Ixodida). PLoS ONE 2013, 8, e78473. [Google Scholar] [CrossRef]
- Jumpertz, M.; Sevestre, J.; Luciani, L.; Houhamdi, L.; Fournier, P.-E.; Parola, P. Bacterial Agents Detected in 418 Ticks Removed from Humans during 2014–2021, France. Emerg. Infect. Dis. 2023, 29, 701. [Google Scholar] [CrossRef] [PubMed]
- Vial, L.; Diatta, G.; Tall, A.; Hadj Ba, E.; Bouganali, H.; Durand, P.; Sokhna, C.; Rogier, C.; Renaud, F.; Trape, J.-F. Incidence of Tick-Borne Relapsing Fever in West Africa: Longitudinal Study. Lancet 2006, 368, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Ndiaye, E.H.I.; Diouf, F.S.; Ndiaye, M.; Bassene, H.; Raoult, D.; Sokhna, C.; Parola, P.; Diatta, G. Tick-Borne Relapsing Fever Borreliosis, a Major Public Health Problem Overlooked in Senegal. PLoS Negl. Trop. Dis. 2021, 15, e0009184. [Google Scholar] [CrossRef] [PubMed]
- Edlinger, E.; Rodhain, F.; Perez, C. Lyme Disease in Patients Previously Suspected of Arbovirus Infection. Lancet 1985, 2, 93. [Google Scholar] [CrossRef]
- Diatta, G.; Vial, L.; Niang, O.; Bouganali, C.; Trape, J.-F. Enquête sur la Borréliose à Tiques à Borrelia Crocidurae au Sénégal; Laboratoire de Paludologie et Zoologie médicale, Institut de Recherche pour le Développement: Dakar, Senegal, 2005; p. 33. [Google Scholar]
- Trape, J.F.; Duplantier, J.M.; Bouganali, H.; Godeluck, B.; Legros, F.; Cornet, J.P.; Camicas, J.L. Tick-Borne Borreliosis in West Africa. Lancet 1991, 337, 473–475. [Google Scholar] [CrossRef]
- Trape, J.F.; Godeluck, B.; Diatta, G.; Rogier, C.; Legros, F.; Albergel, J.; Pepin, Y.; Duplantier, J.M. Tick-Borne Borreliosis in West Africa: Recent Epidemiological Studies. Rocz. Akad. Med. Bialymst. 1996, 41, 136–141. [Google Scholar] [PubMed]
- Ba, E.H. Epidémiologie Clinique de La Borréliose à Tiques Dans Une Zone Rurale Du Sénégal; Mémoire de Fin d’Études d’Ingénieur; University Cheikh Anta Diop de Dakar: Dakar, Senegal, 2001; p. 58. [Google Scholar]
- Sokhna, C.; Mediannikov, O.; Fenollar, F.; Bassene, H.; Diatta, G.; Tall, A.; Trape, J.-F.; Drancourt, M.; Raoult, D. Point-of-Care Laboratory of Pathogen Diagnosis in Rural Senegal. PLoS Negl. Trop. Dis. 2013, 7, e1999. [Google Scholar] [CrossRef] [PubMed]
- Abat, C.; Colson, P.; Chaudet, H.; Rolain, J.-M.; Bassene, H.; Diallo, A.; Mediannikov, O.; Fenollar, F.; Raoult, D.; Sokhna, C. Implementation of Syndromic Surveillance Systems in Two Rural Villages in Senegal. PLoS Negl. Trop. Dis. 2016, 10, e0005212. [Google Scholar] [CrossRef] [PubMed]
- Ndiaye, E.H.I.; Diatta, G.; Zan, D.A.; Bassene, H.; Sokhna, C.; Parola, P. Quantitative Polymerase Chain Reaction from Malaria Rapid Diagnostic Tests to Detect Borrelia Crocidurae, the Agent of Tick-Borne Relapsing Fever, in Febrile Patients in Senegal. Am. J. Trop. Med. Hyg. 2023, 108, 968–976. [Google Scholar] [CrossRef]
- Mediannikov, O.; Fenollar, F.; Socolovschi, C.; Diatta, G.; Bassene, H.; Molez, J.-F.; Sokhna, C.; Trape, J.-F.; Raoult, D. Coxiella Burnetii in Humans and Ticks in Rural Senegal. PLoS Negl. Trop. Dis. 2010, 4, e654. [Google Scholar] [CrossRef]
- Mediannikov, O.; Diatta, G.; Kasongo, K.; Raoult, D. Identification of Bartonellae in the Soft Tick Species Ornithodoros Sonrai in Senegal. Vector Borne Zoonotic Dis. 2014, 14, 26–32. [Google Scholar] [CrossRef]
- Medkour, H.; Lo, C.I.; Anani, H.; Fenollar, F.; Mediannikov, O. Bartonella massiliensis sp. nov., a New Bacterial Species Isolated from an Ornithodoros Sonrai Tick from Senegal. New Microbes New Infect. 2019, 32, 100596. [Google Scholar] [CrossRef]
- Mediannikov, O.; Nguyen, T.-T.; Bell-Sakyi, L.; Padmanabhan, R.; Fournier, P.-E.; Raoult, D. High Quality Draft Genome Sequence and Description of Occidentia massiliensis gen. nov., sp. nov., a New Member of the Family Rickettsiaceae. Stand. Genom. Sci. 2014, 9, 9. [Google Scholar] [CrossRef]
- Raoult, D.; Marrie, T. Q Fever. Clin. Infect. Dis. 1995, 20, 489–496. [Google Scholar] [CrossRef]
- Mediannikov, O.; El Karkouri, K.; Diatta, G.; Robert, C.; Fournier, P.-E.; Raoult, D. Non-Contiguous Finished Genome Sequence and Description of Bartonella senegalensis sp. nov. Stand. Genom. Sci. 2013, 8, 279–289. [Google Scholar] [CrossRef]
- Diarra, A.Z.; Almeras, L.; Laroche, M.; Berenger, J.-M.; Koné, A.K.; Bocoum, Z.; Dabo, A.; Doumbo, O.; Raoult, D.; Parola, P. Molecular and MALDI-TOF identification of ticks and tick-associated bacteria in Mali. PLoS Negl. Trop. Dis. 2017, 11, e0005762. [Google Scholar] [CrossRef]
- Mediannikov, O.; Fenollar, F. Looking in Ticks for Human Bacterial Pathogens. Microb. Pathog. 2014, 77, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Sevestre, J.; Diarra, A.Z.; Laroche, M.; Almeras, L.; Parola, P. Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry: An Emerging Tool for Studying the Vectors of Human Infectious Diseases. Future Microbiol. 2021, 16, 323–340. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Juanes, F.; Calvo Sánchez, N.; Belhassen García, M.; Vieira Lista, C.; Román, R.M.; Álamo Sanz, R.; Muro Álvarez, A.; Muñoz Bellido, J.L. Applications of MALDI-TOF Mass Spectrometry to the Identification of Parasites and Arthropod Vectors of Human Diseases. Microorganisms 2022, 10, 2300. [Google Scholar] [CrossRef] [PubMed]
- Ndiaye, E.H.I.; Diatta, G.; Diarra, A.Z.; Berenger, J.M.; Bassene, H.; Mediannikov, O.; Bouganali, C.; Sokhna, C.; Parola, P. Morphological, Molecular and MALDI-TOF MS Identification of Bedbugs and Associated Wolbachia Species in Rural Senegal. J. Med. Entomol. 2022, 59, 1019–1032. [Google Scholar] [CrossRef] [PubMed]
- Mediannikov, O.; Socolovschi, C.; Bassene, H.; Diatta, G.; Ratmanov, P.; Fenollar, F.; Sokhna, C.; Raoult, D. Borrelia Crocidurae Infection in Acutely Febrile Patients, Senegal. Emerg. Infect. Dis. 2014, 20, 1335–1338. [Google Scholar] [CrossRef]
- Camicas, J.-L.; Morel, P.C. Position systématique et classification des tiques (Acarida:Ixodida). Acarologia (FRA) 1977, 18, 410–420. [Google Scholar]
- Hoogstraal, H. Argasid and Nuttalliellid Ticks as Parasites and Vectors. Adv. Parasitol. 1985, 24, 135–238. [Google Scholar] [CrossRef]
- Tahir, D.; Socolovschi, C.; Marié, J.L.; Ganay, G.; Berenger, J.M.; Bompar, J.M.; Blanchet, D.; Cheuret, M.; Mediannikov, O.; Raoult, D.; et al. New Rickettsia Species in Soft Ticks Ornithodoros Hasei Collected from Bats in French Guiana. Ticks Tick Borne Dis. 2016, 7, 1089–1096. [Google Scholar] [CrossRef]
- Diarra, A.Z.; Laroche, M.; Berger, F.; Parola, P. Use of MALDI-TOF MS for the Identification of Chad Mosquitoes and the Origin of Their Blood Meal. Am. J. Trop. Med. Hyg. 2019, 100, 47–53. [Google Scholar] [CrossRef]
- Roux, V.; Raoult, D. Body Lice as Tools for Diagnosis and Surveillance of Reemerging Diseases. J. Clin. Microbiol. 1999, 37, 596–599. [Google Scholar] [CrossRef]
- Dahmani, M.; Sambou, M.; Scandola, P.; Raoult, D.; Fenollar, F.; Mediannikov, O. Bartonella Bovis and Candidatus Bartonella Davousti in Cattle from Senegal. Comp. Immunol. Microbiol. Infect. Dis. 2017, 50, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Dillon, B.; Valenzuela, J.; Don, R.; Blanckenberg, D.; Wigney, D.I.; Malik, R.; Morris, A.J.; Robson, J.M.; Iredell, J. Limited Diversity among Human Isolates of Bartonella Henselae. J. Clin. Microbiol. 2002, 40, 4691–4699. [Google Scholar] [CrossRef] [PubMed]
- Inokuma, H.; Raoult, D.; Brouqui, P. Detection of Ehrlichia Platys DNA in Brown Dog Ticks (Rhipicephalus Sanguineus) in Okinawa Island, Japan. J. Clin. Microbiol. 2000, 38, 4219–4221. [Google Scholar] [CrossRef]
- Fotso Fotso, A.; Mediannikov, O.; Diatta, G.; Almeras, L.; Flaudrops, C.; Parola, P.; Drancourt, M. MALDI-TOF Mass Spectrometry Detection of Pathogens in Vectors: The Borrelia Crocidurae/Ornithodoros Sonrai Paradigm. PLoS Negl. Trop. Dis. 2014, 8, e2984. [Google Scholar] [CrossRef] [PubMed]
- Benkacimi, L.; Gazelle, G.; El Hamzaoui, B.; Bérenger, J.-M.; Parola, P.; Laroche, M. MALDI-TOF MS Identification of Cimex Lectularius and Cimex Hemipterus Bedbugs. Infect. Genet. Evol. 2020, 85, 104536. [Google Scholar] [CrossRef] [PubMed]
- Yssouf, A.; Flaudrops, C.; Drali, R.; Kernif, T.; Socolovschi, C.; Berenger, J.-M.; Raoult, D.; Parola, P. Matrix-Assisted Laser Desorption Ionization–Time of Flight Mass Spectrometry for Rapid Identification of Tick Vectors. J. Clin. Microbiol. 2013, 51, 522–528. [Google Scholar] [CrossRef]
- Ahamada, S.M.; Diarra, A.Z.; Almeras, L.; Parola, P. Identification of Ticks from an Old Collection by MALDI-TOF MS. J. Proteom. 2022, 264, 104623. [Google Scholar] [CrossRef]
- Elbir, H.; Henry, M.; Diatta, G.; Mediannikov, O.; Sokhna, C.; Tall, A.; Socolovschi, C.; Cutler, S.J.; Bilcha, K.D.; Ali, J.; et al. Multiplex Real-Time PCR Diagnostic of Relapsing Fevers in Africa. PLoS Negl. Trop. Dis. 2013, 7, e2042. [Google Scholar] [CrossRef]
- Medkour, H.; Castaneda, S.; Amona, I.; Fenollar, F.; André, C.; Belais, R.; Mungongo, P.; Muyembé-Tamfum, J.-J.; Levasseur, A.; Raoult, D.; et al. Potential Zoonotic Pathogens Hosted by Endangered Bonobos. Sci. Rep. 2021, 11, 6331. [Google Scholar] [CrossRef]
- Diatta, G.; Duplantier, J.-M.; Granjon, L.; Bâ, K.; Chauvancy, G.; Ndiaye, M.; Trape, J.-F. Borrelia Infection in Small Mammals in West Africa and Its Relationship with Tick Occurrence inside Burrows. Acta Trop. 2015, 152, 131–140. [Google Scholar] [CrossRef]
- Diatta, G.; Mediannikov, O.; Boyer, S.; Sokhna, C.; Bassène, H.; Fenollar, F.; Chauvancy, G.; Ndiaye, A.A.; Diene, F.; Parola, P.; et al. An Alternative Strategy of Preventive Control of Tick-Borne Relapsing Fever in Rural Areas of Sine-Saloum, Senegal. Am. J. Trop. Med. Hyg. 2016, 95, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Trape, J.F.; Godeluck, B.; Diatta, G.; Rogier, C.; Legros, F.; Albergel, J.; Pepin, Y.; Duplantier, J.M. The Spread of Tick-Borne Borreliosis in West Africa and Its Relationship to Sub-Saharan Drought. Am. J. Trop. Med. Hyg. 1996, 54, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Fall, F.K.; Laroche, M.; Bossin, H.; Musso, D.; Parola, P. Performance of MALDI-TOF Mass Spectrometry to Determine the Sex of Mosquitoes and Identify Specific Colonies from French Polynesia. Am. J. Trop. Med. Hyg. 2021, 104, 1907. [Google Scholar] [CrossRef]
- Vial, L.; Durand, P.; Arnathau, C.; Halos, L.; Diatta, G.; Trape, J.F.; Renaud, F. Molecular Divergences of the Ornithodoros Sonrai Soft Tick Species, a Vector of Human Relapsing Fever in West Africa. Microbes Infect. 2006, 8, 2605–2611. [Google Scholar] [CrossRef]
- Benyahia, H.; Ouarti, B.; Diarra, A.Z.; Boucheikhchoukh, M.; Meguini, M.N.; Behidji, M.; Benakhla, A.; Parola, P.; Almeras, L. Identification of Lice Stored in Alcohol Using MALDI-TOF MS. J. Med. Entomol. 2021, 58, 1126–1133. [Google Scholar] [CrossRef]
- Ngoy, S.; Diarra, A.Z.; Laudisoit, A.; Gembu, G.-C.; Verheyen, E.; Mubenga, O.; Mbalitini, S.G.; Baelo, P.; Laroche, M.; Parola, P. Using MALDI-TOF Mass Spectrometry to Identify Ticks Collected on Domestic and Wild Animals from the Democratic Republic of the Congo. Exp. Appl. Acarol. 2021, 84, 637–657. [Google Scholar] [CrossRef]
- Dahmana, H.; Granjon, L.; Diagne, C.; Davoust, B.; Fenollar, F.; Mediannikov, O. Rodents as Hosts of Pathogens and Related Zoonotic Disease Risk. Pathogens 2020, 9, E202. [Google Scholar] [CrossRef]
- Meerburg, B.G.; Singleton, G.R.; Kijlstra, A. Rodent-Borne Diseases and Their Risks for Public Health. Crit. Rev. Microbiol. 2009, 35, 221–270. [Google Scholar] [CrossRef] [PubMed]
- Ouarti, B.; Sall, M.; Ndiaye, E.H.I.; Diatta, G.; Zan Diarra, A.; Berenger, J.M.; Sokhna, C.; Granjon, L.; Le Fur, J.; Parola, P. Pathogen Detection in Ornithodoros Sonrai Ticks and Invasive House Mice Mus Musculus Domesticus in Senegal. Microorganisms 2022, 10, 2367. [Google Scholar] [CrossRef]
- Angelakis, E.; Billeter, S.A.; Breitschwerdt, E.B.; Chomel, B.B.; Raoult, D. Potential for Tick-Borne Bartonelloses. Emerg. Infect. Dis. 2010, 16, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Eldin, C.; Mélenotte, C.; Mediannikov, O.; Ghigo, E.; Million, M.; Edouard, S.; Mege, J.-L.; Maurin, M.; Raoult, D. From Q Fever to Coxiella Burnetii Infection: A Paradigm Change. Clin. Microbiol. Rev. 2017, 30, 115–190. [Google Scholar] [CrossRef] [PubMed]
- Dumler, J.S.; Barbet, A.F.; Bekker, C.P.; Dasch, G.A.; Palmer, G.H.; Ray, S.C.; Rikihisa, Y.; Rurangirwa, F.R. Reorganization of Genera in the Families Rickettsiaceae and Anaplasmataceae in the Order Rickettsiales: Unification of Some Species of Ehrlichia with Anaplasma, Cowdria with Ehrlichia and Ehrlichia with Neorickettsia, Descriptions of Six New Species Combinations and Designation of Ehrlichia Equi and “HGE Agent” as Subjective Synonyms of Ehrlichia Phagocytophila. Int. J. Syst. Evol. Microbiol. 2001, 51, 2145–2165. [Google Scholar] [CrossRef] [PubMed]
- Stuen, S.; Granquist, E.G.; Silaghi, C. Anaplasma Phagocytophilum—A Widespread Multi-Host Pathogen with Highly Adaptive Strategies. Front. Cell Infect. Microbiol. 2013, 3, 31. [Google Scholar] [CrossRef]
- Sy, I.; Margardt, L.; Ngbede, E.O.; Adah, M.I.; Yusuf, S.T.; Keiser, J.; Rehner, J.; Utzinger, J.; Poppert, S.; Becker, S.L. Identification of Adult Fasciola Spp. Using Matrix-Assisted Laser/Desorption Ionization Time-of-Flight (MALDI-TOF) Mass Spectrometry. Microorganisms 2021, 9, 82. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| System Name | Assay Specificity | Targeted Gene | Primer Sequences | Reference |
|---|---|---|---|---|
| Real Time Quantitative PCR | Borrelia spp. | ITS4 | Bor_ITS4_ F (5’-GGCTTCGGGTCTACCACATCTA-3’) Bor_ITS4_ R (5’-CCGGGAGGGGAGTGAAATAG-3’) Bor_ITS4_ P (6FAM-TGCAAAAGGCACGCCATCACC) | [32] |
| B. crocidurae | GlpQ | Bcroci_glpQ_F (5’-CCTTGGATACCCCAAATCATC-3’) Bcroci_glpQ_R (5’-GGCAATGCATCAATTCTAAAC-3’) Bcroci_glpQ_P (6FAM-ATGGACAAATGACAGGTCTTAC –MGB | ||
| Bartonella spp. | ITS2 | Barto_ITS2_F (5’- GGGGCCGTAGCTCAGCTG-3’) Barto_ITS2_R (5’- TGAATATATCTTCTCTTCACAATTTC-3’) Barto_ITS2_P (6FAM- CGATCCCGTCCGGCTCCACCA) | [32] | |
| Anaplasmataceae | qANA | TtAna_F (5’-TGACAGCGTACCTTTTGCAT-3’) TtAna_R (5’- GTAACAGGTTCGGTCCTCCA-3’) TtAna_P (6FAM- GGATTAGACCCGAAACCAAG) | [32] | |
| Coxiella burnetii | ISS11 | CB_IS1111_0706F (5’-CAAGAAACGTATCGCTGTGGC-3’) CB_IS1111_0706R (5’-CACAGAGCCACCGTATGAATC-3’) CB_IS1111_0706P (6FAM-CCGAGTTCGAAACAATGAGGGCTG) | [32] | |
| IS30A | CB_IS30A_3F (5’- CGCTGACCTACAGAAATATGTCC-3’) CB_IS30A_3R (5’-GGGGTAAGTAAATAATACCTTCTGG-3’) CB_IS30A_3P (6FAM- CATGAAGCGATTTATCAATACGTGTATGC) | [32] | ||
| Rickettsia | RKND | RKND03_F (5’-GTGAATGAAAGATTACACTATTTAT-3’) RKND03_R (5’-GTATCTTAGCAATCATTCTAATAGC-3’) RKND03_R (6FAM- CTATTATGCTTGCGGCTGTCGGTTC) | [32] | |
| Standard PCR | Ticks | 16S | 16S_Tick_F (5′-CCGGTCTGAACTCAGATCAAGT -3′) 16S_Tick_R (5′-GCTCAATGATTTTTTAAATTGCTGT-3′) | [23] |
| Borrelia spp. | 16S | B_F1 (5’-GCTGGCAGTGCGTCTTAAGC -3’) B_R1 (5’-GCTTCGGGTATCCTCAACTC -3’) | [33] | |
| Anaplasmataceae | 23S rRNA (rrl) | Ana23S-212_F (5’-ATAAGCTGCGGGGAATTGTC-3’) Ana23S-753_R (5’-TGCAAAAGGTACGCTGTCAC-3’) | [27] | |
| Bartonella spp. | FtsZ | FTSZDIR-F (5’-CCGTGAATAATATGATTAATGC-3’) FTSZREV-R (5’-TTGAAATGGCTTTGTCACAAC-3’) | [34] | |
| GltA | GltA-F (5’- TTACTTATGATCCKGGYTTTA-3’) GltA-R (5’-AATGCAAAAAGAACAGTAAACA-3’) | [35] | ||
| Wolbachia spp. | FtsZ | Wol-99F (5’-TTGTAGCCTGCTATGGTATAACT-3’) Wol-994R (5’-GAATAGGTATGATTTTCATGT-3’) | [27] | |
| Ehrlichia spp. | 16S | Ehr-16S-F (5’-GGTACCYACAGAAGAAGTCC-3’) Ehr-16S-R (5’-TAGCACTCATCGTTTACAGC-3’) | [36] |
| Area of Collection | Villages Infested/Prospected | Households Investigated | Positive Households (%) | Burrows Prospected | Positive Burrows (%) | Tick Storing Medium | Number of Specimens Collected |
|---|---|---|---|---|---|---|---|
| Niakhar | 17/17 | 173 | 92 (53%) | 541 | 219 (41%) | Alcohol | 338 |
| Silica gel | 481 | ||||||
| Sine-Saloum | 10/15 | 148 | 34 (23%) | 334 | 58 (17%) | Alcohol | 322 |
| Silica gel | 134 | ||||||
| Total | 32 | 321 | 126 (39%) | 875 | 277 (32%) | 1275 | |
| Storage Medium | Area | Number of Villages | % MALDI-TOF MS Specimens Tested/Collected | % Good Spectrum | Spectra in Data | % Identification [LSVs Range] |
|---|---|---|---|---|---|---|
| Alcohol | Niakhar | 2/17 | 10/338 | 50% (5/10) | 1 | 100% (4/4) [1.8–2.5] |
| Sine-Saloum | 6/15 | 149/322 | 89% (134/149) | 9 | 100% (125/125) [1.6–2.5] | |
| Silica gel | Niakhar | 9/17 | 100/481 | 95% (95/100) | 4 | 100% (91/91) [1.8–2.6] |
| Total | 17/32 | 23% (259/1141) | 90% (234/259) | 14 | 100% (220/220) [1.6–2.6] | |
| qPCR Pathogen Tested | Proportion of Positive Samples Per Village | Total | |
|---|---|---|---|
| Sine-Saloum | Niakhar | ||
| Prevalence of pathogen detected in O. sonrai | |||
| Borrelia crocidurae | 33%; 43/130 | 19%; 23/119 | 26%, 66/249 |
| Coxiella burnetii | 0%; 0/130 | 1.7%; 2/119 | 0.8%, 2/249 |
| Bartonella spp. | 19%; 25/130 | 51%; 61/119 | 35%, 86/249 |
| Anaplasmataceae | 23%; 30/130 | 33%; 39/119 | 28%, 69/249 |
| Rickettsia spp. | 0%; 0/130 | 0%; 0/119 | 0% |
| Prevalence of double co-infections detected | |||
| B. crocidurae + Anaplasmataceae | 8.5%; 11/130 | 4.2%; 5/119 | 6.4%, 16/249 |
| B. crocidurae + Bartonella spp. | 6.2%; 8/130 | 1.7%; 2/119 | 4%, 10/249 |
| B. crocidurae + C. burnetii | 0%; 0/130 | 0.8%; 1/119 | 0.4%, 1/249 |
| Anaplasmataceae + Bartonella spp. | 2.3%; 3/130 | 10%; 12/119 | 6%, 15/249 |
| Prevalence of triple co-infections detected | |||
| Anaplasmataceae + B. crocidurae + Bartonella spp. | 4.6%; 6/130 | 6.7%; 8/119 | 5.6%, 14/249 |
| Anaplasmataceae + B. crocidurae + C. burnetii | 0%; 0/130 | 0.8%; 1/119 | 0.4%, 1/249 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ndiaye, E.H.I.; Diarra, A.Z.; Diouf, F.S.; Bouganali, C.; Almeras, L.; Sokhna, C.; Diatta, G.; Parola, P. Ornithodoros sonrai Soft Ticks and Associated Bacteria in Senegal. Pathogens 2023, 12, 1078. https://doi.org/10.3390/pathogens12091078
Ndiaye EHI, Diarra AZ, Diouf FS, Bouganali C, Almeras L, Sokhna C, Diatta G, Parola P. Ornithodoros sonrai Soft Ticks and Associated Bacteria in Senegal. Pathogens. 2023; 12(9):1078. https://doi.org/10.3390/pathogens12091078
Chicago/Turabian StyleNdiaye, El Hadji Ibrahima, Adama Zan Diarra, Fatou Samba Diouf, Charles Bouganali, Lionel Almeras, Cheikh Sokhna, Georges Diatta, and Philippe Parola. 2023. "Ornithodoros sonrai Soft Ticks and Associated Bacteria in Senegal" Pathogens 12, no. 9: 1078. https://doi.org/10.3390/pathogens12091078