
Effect of Mannan Oligosaccharides Extracts in Uropathogenic Escherichia coli Adhesion in Human Bladder Cells
 , , , , ,
, , , , ,  , and
, and 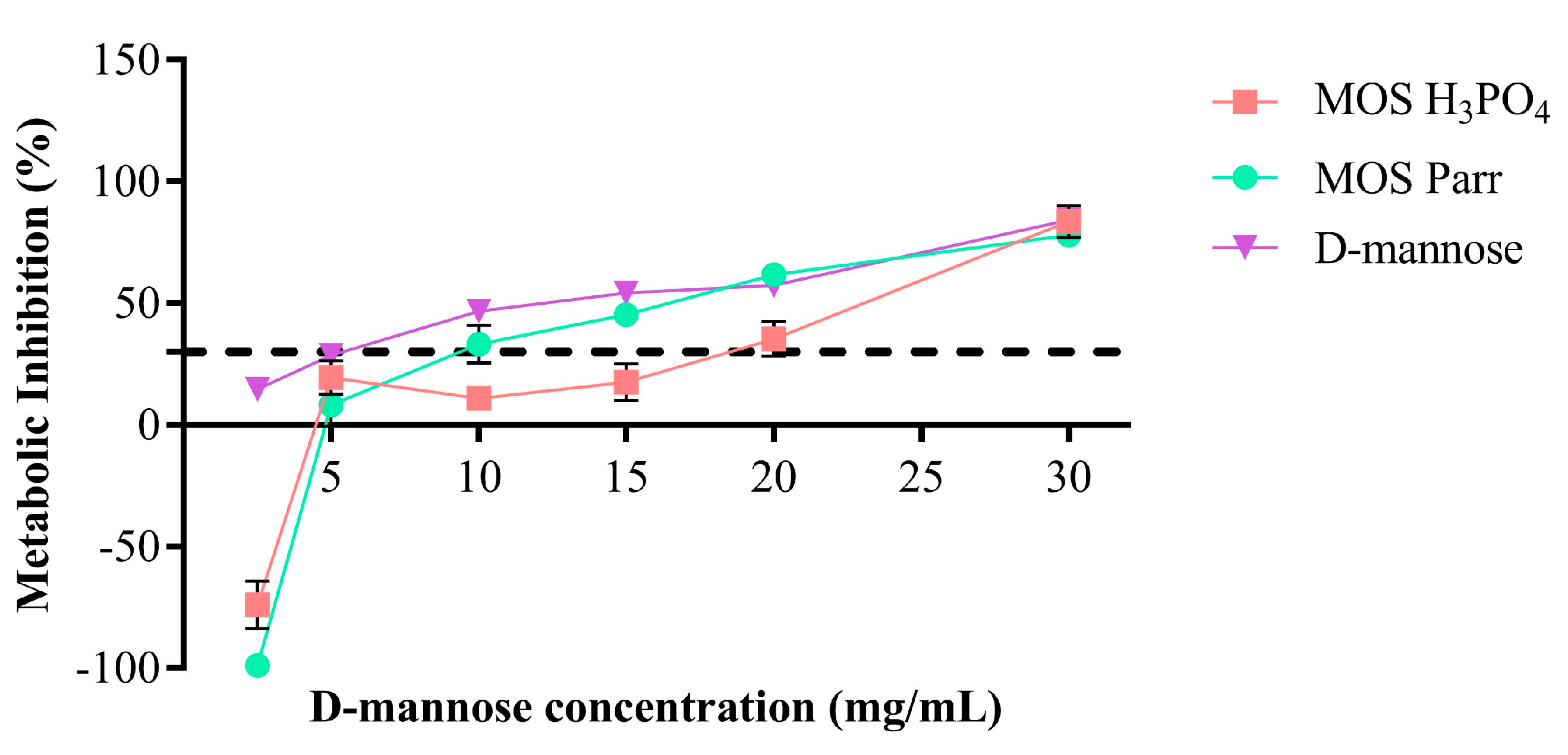{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Bacterial Strain and Cell Line
2.3. Cytotoxicity Assay
2.4. Immunological Response without Stimulus
2.5. UPEC Adhesion Assays
2.5.1. Simultaneous Exposure (Competition)
2.5.2. Prophylaxis Assessment
2.6. Total Viable Counts Determination
2.7. Protein Determination and Interleukin Evaluation
2.8. Antimicrobial Activity–Growth Inhibition Curves
2.9. Statistical Analysis
3. Results and Discussion
3.1. Cytotoxicity
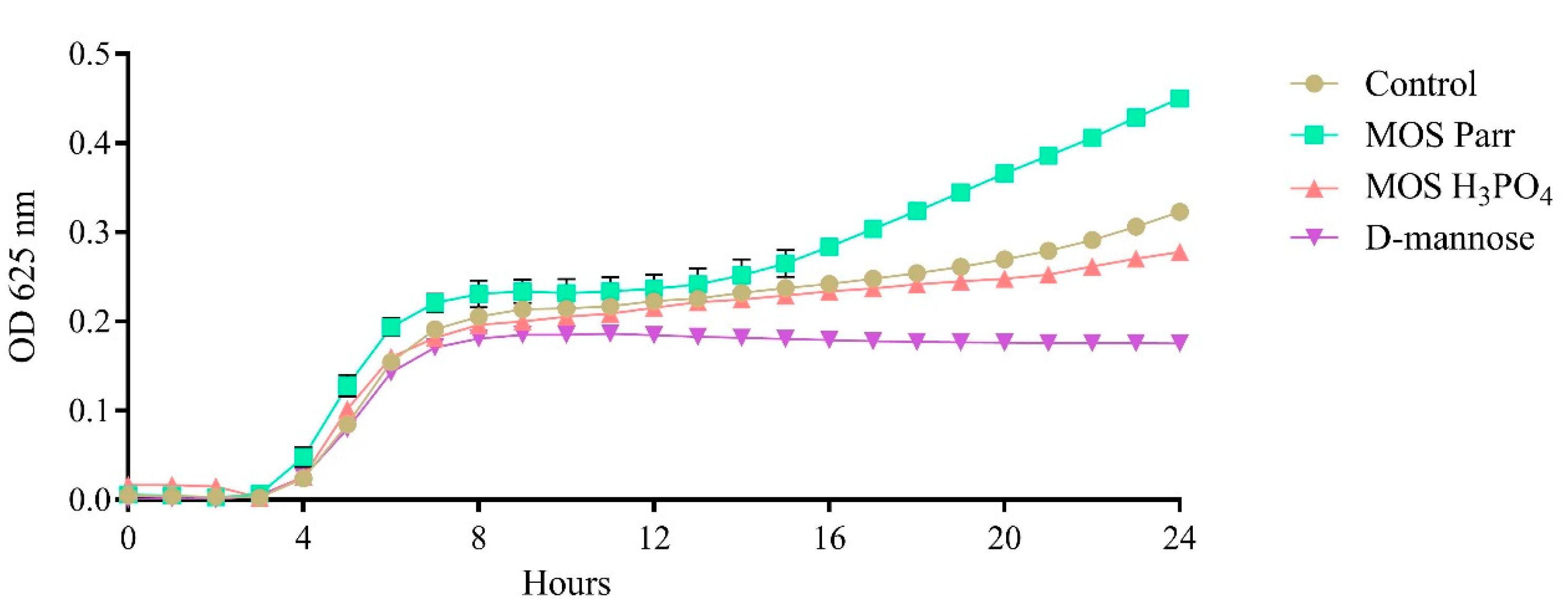
3.2. Impact upon UPEC Growth
3.3. Immunologic Response (1, 2 and 3 h Exposures to the Samples)
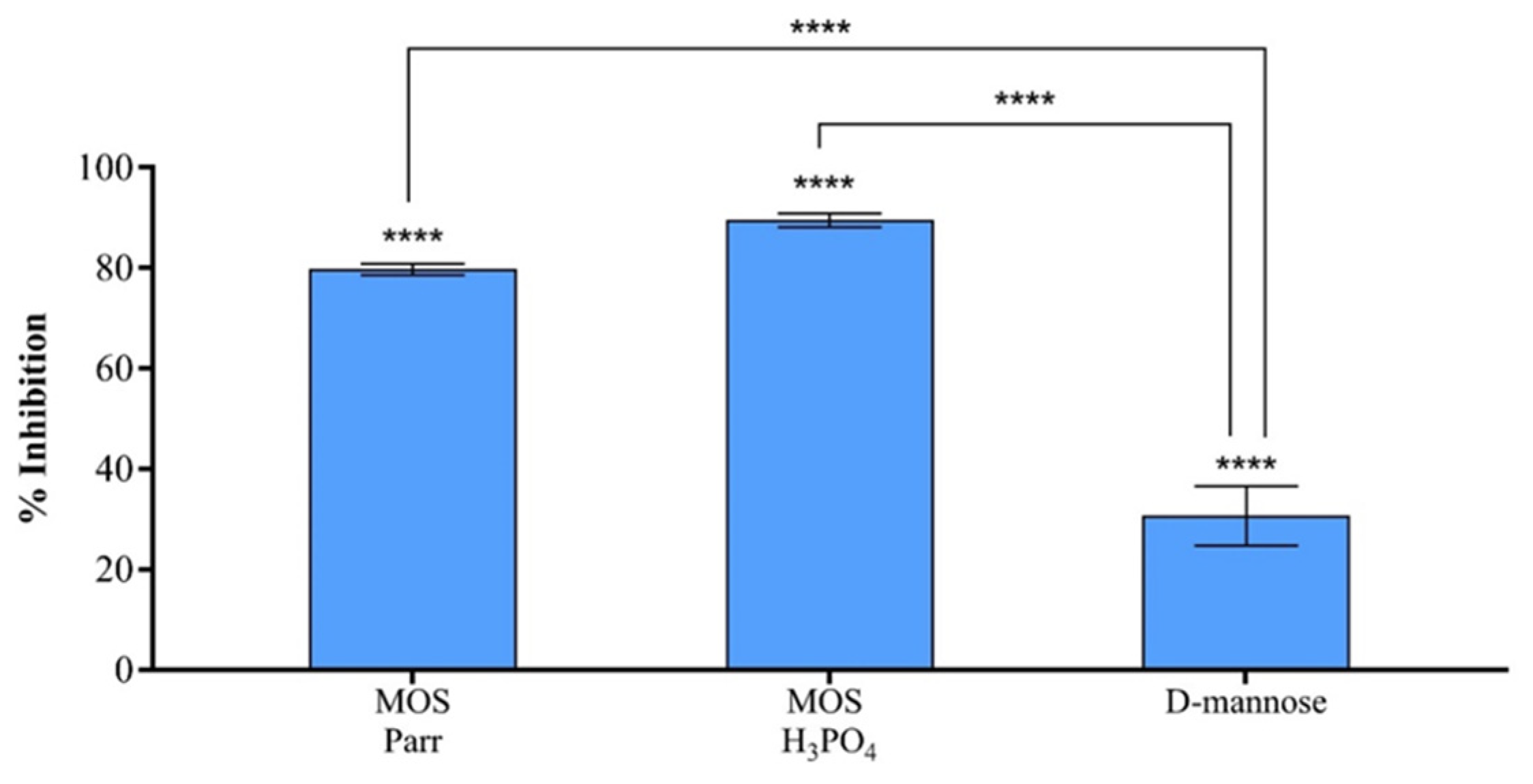
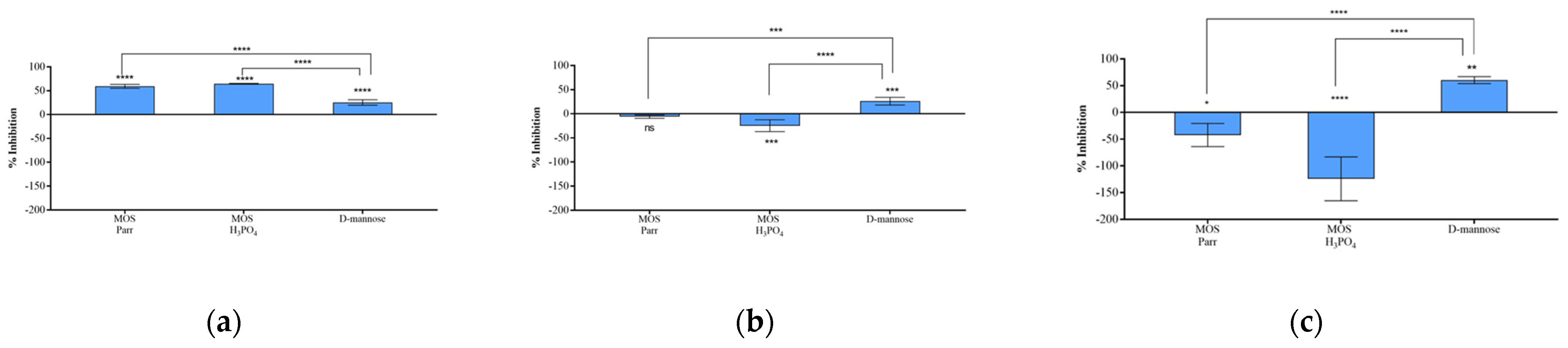
3.4. MOS Extracts Inhibitory Effect on UPEC Adhesion to HTB-9
3.5. Prophylactic Potential of MOS Extracts
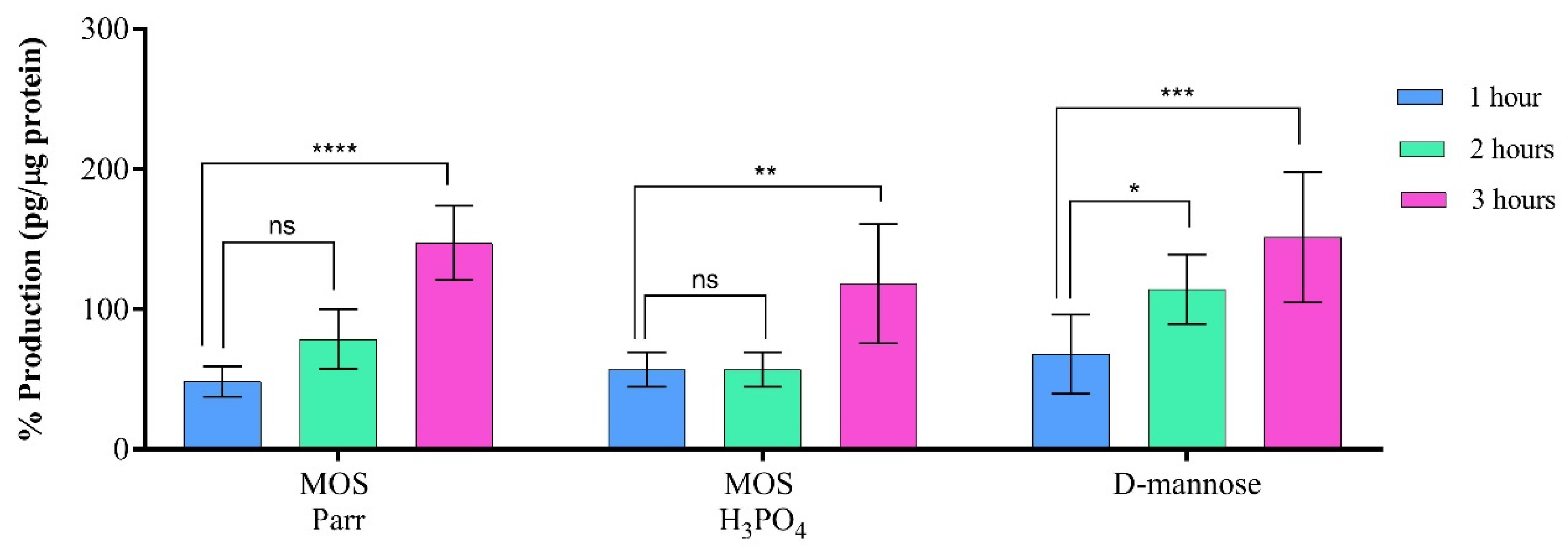
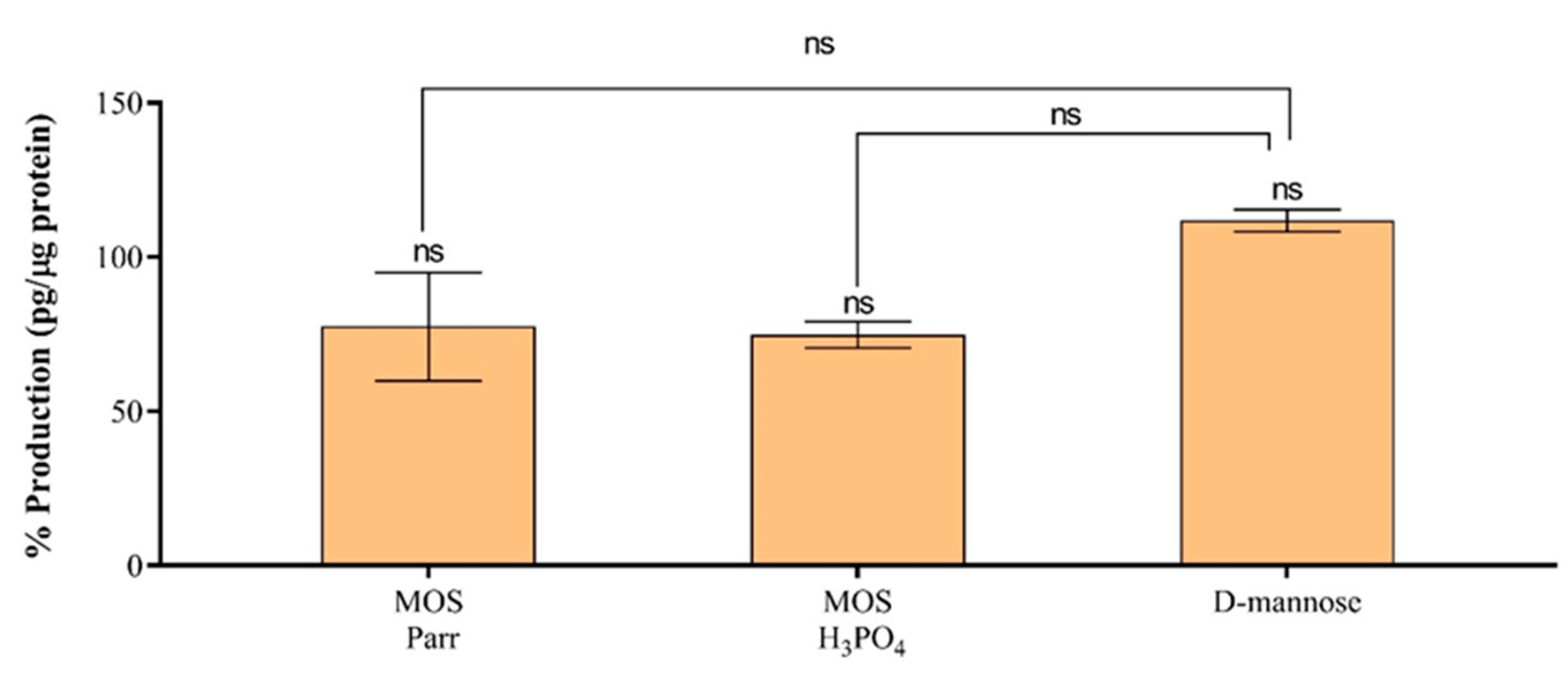
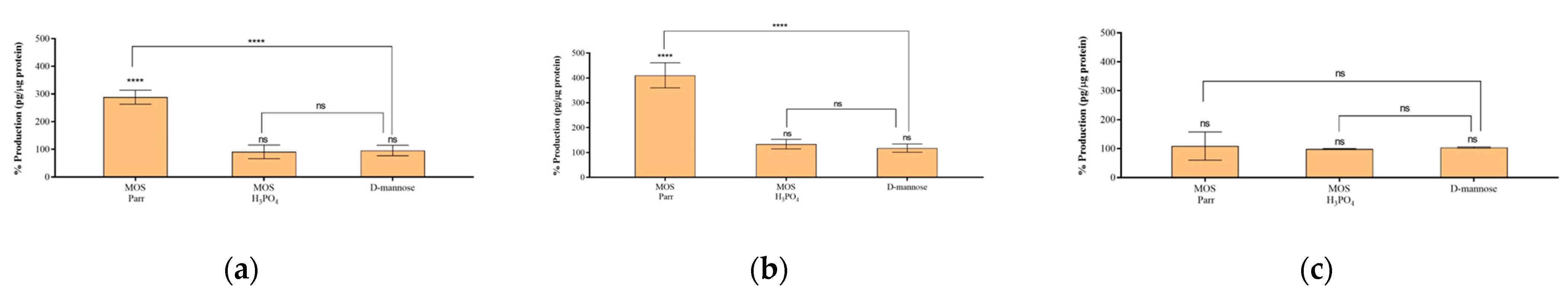
3.6. Cytokine Production in the Presence of UPEC
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Flores-Mireles, A.L.; Walker, J.N.; Caparon, M.; Hultgren, S.J. Urinary Tract Infections: Epidemiology, Mechanisms of Infection and Treatment Options. Nat. Rev. Microbiol. 2015, 13, 269–284. [Google Scholar] [CrossRef]
- Terlizzi, M.E.; Gribaudo, G.; Maffei, M.E. UroPathogenic Escherichia coli (UPEC) Infections: Virulence Factors, Bladder Responses, Antibiotic, and Non-Antibiotic Antimicrobial Strategies. Front. Microbiol. 2017, 8, 1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalas, V.; Hibbing, M.E.; Maddirala, A.R.; Chugani, R.; Pinkner, J.S.; Mydock-McGrane, L.K.; Conover, M.S.; Janetka, J.W.; Hultgren, S.J. Structure-Based Discovery of Glycomimetic FmlH Ligands as Inhibitors of Bacterial Adhesion during Urinary Tract Infection. Proc. Natl. Acad. Sci. USA 2018, 115, E2819–E2828. [Google Scholar] [CrossRef] [Green Version]
- Magliano, E.; Grazioli, V.; Deflorio, L.; Leuci, A.I.; Mattina, R.; Romano, P.; Cocuzza, C.E. Gender and Age-Dependent Etiology of Community-Acquired Urinary Tract Infections. Sci. World J. 2012, 2012, 349597. [Google Scholar] [CrossRef] [Green Version]
- Vyas, S.; Varshney, D.; Sharma, P.; Juyal, R.; Nautiyal, V.; Shrotriya, V. An Overview of the Predictors of Symptomatic Urinary Tract Infection among Nursing Students. Ann. Med. Health Sci. Res. 2015, 5, 54. [Google Scholar] [CrossRef] [Green Version]
- Bonkat, G.; Bartoletti, R.; Bruyère, F.; Cai, T.; Geerlings, S.E.; Köyes, B.; Kranz, J.; Schubert, S.; Pilatz, A.; Veeratterapillay, R.; et al. EAU Guidelines, Proceedings of the EAU Annual Congress, Milan, Italy, 10–13 March 2023; EAU Guidelines Office: Amhen, The Nertherlands, 2023; ISBN 978-94-92671-19-6. [Google Scholar]
- McLellan, L.K.; Hunstad, D.A. Urinary Tract Infection: Pathogenesis and Outlook. Trends Mol. Med. 2016, 22, 946–957. [Google Scholar] [CrossRef] [Green Version]
- Forsyth, V.S.; Armbruster, C.E.; Smith, S.N.; Pirani, A.; Springman, A.C.; Walters, M.S.; Nielubowicz, G.R.; Himpsl, S.D.; Snitkin, E.S.; Mobley, H.L.T. Rapid Growth of Uropathogenic Escherichia coli during Human Urinary Tract Infection. mBio 2018, 9, e00186-18. [Google Scholar] [CrossRef] [Green Version]
- Hannan, T.J.; Mysorekar, I.U.; Hung, C.S.; Isaacson-Schmid, M.L.; Hultgren, S.J. Early Severe Inflammatory Responses to Uropathogenic E. coli Predispose to Chronic and Recurrent Urinary Tract Infection. PLOS Pathog. 2010, 6, e1001042. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, D.J.; Chen, S.L.; Hultgren, S.J.; Seed, P.C. Population Dynamics and Niche Distribution of Uropathogenic Escherichia Coli during Acute and Chronic Urinary Tract Infection. Infect. Immun. 2011, 79, 4250–4259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribić, R.; Meštrović, T.; Neuberg, M.; Kozina, G. Effective Anti-Adhesives of Uropathogenic Escherichia coli. Acta Pharm. 2018, 68, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mydock-Mcgrane, L.K.; Cusumano, Z.T.; Janetka, J.W. Mannose-Derived FimH Antagonists: A Promising Anti-Virulence Therapeutic Strategy for Urinary Tract Infections and Crohn’s Disease. Expert Opin. Ther. Pat. 2016, 26, 175–197. [Google Scholar] [CrossRef]
- Mickiewicz, K.M.; Kawai, Y.; Drage, L.; Gomes, M.C.; Davison, F.; Pickard, R.; Hall, J.; Mostowy, S.; Aldridge, P.D.; Errington, J. Possible Role of L-Form Switching in Recurrent Urinary Tract Infection. Nat. Commun. 2019, 10, 4379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, G.G.; Palermo, J.J.; Schilling, J.D.; Roth, R.; Heuser, J.; Hultgren, S.J. Intracellular Bacterial Biofilm-like Pods in Urinary Tract Infections. Science 2003, 301, 105–107. [Google Scholar] [CrossRef] [Green Version]
- Snyder, J.A.; Lloyd, A.L.; Lockatell, C.V.; Johnson, D.E.; Mobley, H.L.T. Role of Phase Variation of Type 1 Fimbriae in a Uropathogenic Escherichia Coli Cystitis Isolate during Urinary Tract Infection. Infect. Immun. 2006, 74, 1387–1393. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.R.; Sun, T.T.; Medina, J.J. In Vitro Binding of Type 1-Fimbriated Escherichia coli to Uroplakins Ia and Ib: Relation to Urinary Tract Infections. Proc. Natl. Acad. Sci. USA 1996, 93, 9630–9635. [Google Scholar] [CrossRef] [Green Version]
- Wei, Z.; Huang, L.; Cui, L.; Zhu, X. Mannose: Good Player and Assister in Pharmacotherapy. Biomed. Pharmacother. 2020, 129, 110420. [Google Scholar] [CrossRef]
- Wu, H.; Zhang, W.; Mu, W. Recent Studies on the Biological Production of D-Mannose. Appl. Microbiol. Biotechnol. 2019, 103, 8753–8761. [Google Scholar] [CrossRef] [PubMed]
- Cusumano, C.K.; Pinkner, J.S.; Han, Z.; Greene, S.E.; Ford, B.A.; Crowley, J.R.; Henderson, J.P.; Janetka, J.W.; Hultgren, S.J. Treatment and Prevention of Urinary Tract Infection with Orally Active FimH Inhibitors. Sci. Transl. Med. 2011, 3, 109ra115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Firon, N.; Ofek, I.; Sharon, N. Interaction of Mannose-Containing Oligosaccharides with the Fimbrial Lectin of Escherichia coli. Biochem. Biophys. Res. Commun. 1982, 105, 1426–1432. [Google Scholar] [CrossRef]
- Klein, T.; Abgottspon, D.; Wittwer, M.; Rabbani, S.; Herold, J.; Jiang, X.; Kleeb, S.; Lüthi, C.; Scharenberg, M.; Bezençon, J.; et al. FimH Antagonists for the Oral Treatment of Urinary Tract Infections: From Design and Synthesis to in vitro and in vivo Evaluation. J. Med. Chem. 2010, 53, 8627–8641. [Google Scholar] [CrossRef] [PubMed]
- Ofek, I.; Mirelman, D.; Sharon, N. Adherence of Escherichia coli to Human Mucosal Cells Mediated by Mannose Receptors. Nature 1977, 265, 623–625. [Google Scholar] [CrossRef]
- O’Brien, V.P.; Hannan, T.J.; Nielsen, H.V.; Hultgren, S.J. Drug and Vaccine Development for the Treatment and Prevention of Urinary Tract Infections. Microbiol. Spectr. 2016, 4, 589–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zacchè, M.M.; Giarenis, I. Therapies in Early Development for the Treatment of Urinary Tract Inflammation. Expert Opin. Investig. Drugs 2016, 25, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Scribano, D.; Sarshar, M.; Prezioso, C.; Lucarelli, M.; Angeloni, A.; Zagaglia, C.; Palamara, A.T.; Ambrosi, C. D-Mannose Treatment Neither Affects Uropathogenic Escherichia coli Properties nor Induces Stable Fimh Modifications. Molecules 2020, 25, 316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pigrau, C.; Escolà-Vergé, L. Recurrent Urinary Tract Infections: From Pathogenesis to Prevention. Med. Clin. 2020, 155, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Serafini-Cessi, F.; Monti, A.; Cavallone, D. N-Glycans Carried by Tamm-Horsfall Glycoprotein Have a Crucial Role in the Defense against Urinary Tract Diseases. Glycoconj. J. 2005, 22, 383–394. [Google Scholar] [CrossRef]
- Faustino, M.; Durão, J.; Pereira, C.F.; Oliveira, A.S.; Pereira, J.O.; Pereira, A.M.; Ferreira, C.; Pintado, M.E.; Carvalho, A.P. Comparative Analysis of Mannans Extraction Processes from Spent Yeast Saccharomyces cerevisiae. Foods 2022, 11, 3753. [Google Scholar] [CrossRef]
- ISO 10993-5; Biological Evaluation of Medical Devices. Part 5: Tests for in vitro Cytotoxicity. International Organization for Standarrization (ISO): Geneve, Switzerland, 2009.
- Miles, A.A.; Misra, S.S.; Irwin, J.O. The Estimation of the Bactericidal Power of the Blood. J. Hyg. 1938, 38, 732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CLSI Methods for Dilution Antimicriobial Susceptibility Tests for Bacteria That Grow Aerobically, 11th ed.; Clinical & Laboratory Standards Institute: Wayne, PA, USA, 2012; Volume 112.
- Svanborg, C.; Godaly, G.; Hedlund, M. Cytokine Responses during Mucosal Infections: Role in Disease Pathogenesis and Host Defence. Curr. Opin. Microbiol. 1999, 2, 99–103. [Google Scholar] [CrossRef]
- Kishimoto, T. Interleukin-6: From Basic Science to Medicine—40 Years in Immunology. Annu. Rev. Immunol. 2005, 23, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in Inflammation, Immunity, and Disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef]
- Moriyama, K.; Nishida, O. Targeting Cytokines, Pathogen-Associated Molecular Patterns, and Damage-Associated Molecular Patterns in Sepsis via Blood Purification. Int. J. Mol. Sci. 2021, 22, 8882. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Jung, S.; Carey, J.B. Effects of a Commercial Mannan-Oligosaccharide Product on Growth Performance, Intestinal Histomorphology, and Amino Acid Digestibility in White Pekin Ducks. J. Appl. Poult. Res. 2019, 28, 72–77. [Google Scholar] [CrossRef]
- Zheng, C.; Zhou, J.; Zeng, Y.; Liu, T. Effects of Mannan Oligosaccharides on Growth Performance, Nutrient Digestibility, Ruminal Fermentation and Hematological Parameters in Sheep. PeerJ 2021, 9, e11631. [Google Scholar] [CrossRef]
- Hooge, D.M.; Sims, M.D.; Sefton, A.E.; Connolly, A.; Spring, P. Effect of Dietary Mannan Oligosaccharide, With or Without Bacitracin or Virginiamycin, on Live Performance of Broiler Chickens at Relatively High Stocking Density on New Litter. J. Appl. Poult. Res. 2003, 12, 461–467. [Google Scholar] [CrossRef]
- Agazzi, A.; Perricone, V.; Omodei Zorini, F.; Sandrini, S.; Mariani, E.; Jiang, X.-R.; Ferrari, A.; Crestani, M.; Nguyen, T.X.; Bontempo, V.; et al. Dietary Mannan Oligosaccharides Modulate Gut Inflammatory Response and Improve Duodenal Villi Height in Post-Weaning Piglets Improving Feed Efficiency. Animals 2020, 10, 1283. [Google Scholar] [CrossRef]
- Halas, V.; Nochta, I. Mannan Oligosaccharides in Nursery Pig Nutrition and Their Potential Mode of Action. Animals 2012, 2, 261–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, C.; Katz, R.; McCulloch, M. Yeast Mannan Oligosaccharide Dietary Supplement in the Treatment of Chronically Acute Urinary Tract Infections: A Case Series. UroToday Int. J. 2012, 5, 50. [Google Scholar] [CrossRef]
- Kim, D.Y.; Lee, J.C. Adherence Assay of Uropathogenic Escherichia coli in vivo and in vitro. Urogenit. Tract Infect. 2017, 12, 122. [Google Scholar] [CrossRef] [Green Version]
- Porru, D.; Parmigiani, A.; Tinelli, C.; Barletta, D.; Choussos, D.; Di Franco, C.; Bobbi, V.; Bassi, S.; Miller, O.; Gardella, B.; et al. Oral D-Mannose in Recurrent Urinary Tract Infections in Women: A Pilot Study. J. Clin. Urol. 2014, 7, 208–213. [Google Scholar] [CrossRef]
- Ruggieri, M.R.; Hanno, P.M.; Levin, R.M. Mannose Inhibition of Escherichia Coli Adherence to Urinary Bladder Epithelium: Comparison with Yeast Agglutination. Urol. Res. 1985, 13, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Taleb, N.S. A Study of Therapeutic Effect of D-Mannose on Urinary Infection Caused by Escherichia coli. MOJ Proteom. Bioinforma. 2018, 7, 172–175. [Google Scholar] [CrossRef]
- Crépin, S.; Houle, S.; Charbonneau, M.È.; Mourez, M.; Harel, J.; Dozois, C.M. Decreased Expression of Type 1 Fimbriae by a Pst Mutant of Uropathogenic Escherichia coli Reduces Urinary Tract Infection. Infect. Immun. 2012, 80, 2802–2815. [Google Scholar] [CrossRef] [Green Version]
- Schwan, W.R.; Beck, M.T.; Hung, C.S.; Hultgren, S.J. Differential Regulation of Escherichia coli Fim Genes Following Binding to Mannose Receptors. J. Pathog. 2018, 2018, 2897581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samuelsson, P.; Hang, L.; Wullt, B.; Irjala, H.; Svanborg, C. Toll-like Receptor 4 Expression and Cytokine Responses in the Human Urinary Tract Mucosa. Infect. Immun. 2004, 72, 3179–3186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores-Mireles, A.L.; Walker, J.N.; Bauman, T.M.; Potretzke, A.M.; Schreiber, H.L.; Park, A.M.; Pinkner, J.S.; Caparon, M.G.; Hultgren, S.J.; Desai, A. Fibrinogen Release and Deposition on Urinary Catheters Placed during Urological Procedures. J. Urol. 2016, 196, 416–421. [Google Scholar] [CrossRef] [Green Version]
- Sarshar, M.; Behzadi, P.; Ambrosi, C.; Zagaglia, C.; Palamara, A.T.; Scribano, D. FimH and Anti-Adhesive Therapeutics: A Disarming Strategy against Uropathogens. Antibiotics 2020, 9, 397. [Google Scholar] [CrossRef]
- Scaglione, F.; Musazzi, U.M.; Minghetti, P. Considerations on D-Mannose Mechanism of Action and Consequent Classification of Marketed Healthcare Products. Front. Pharmacol. 2021, 12, 330. [Google Scholar] [CrossRef]
- McAchran, S.; Rackley, R.; Vasavada, S. Neuromodulation for Voiding Dysfunction. Neuromodulation 2009, 2, 945–956. [Google Scholar] [CrossRef]
- Lukacz, E.S.; Whitcomb, E.L.; Lawrence, J.M.; Nager, C.W.; Luber, K.M. Urinary Frequency in Community-Dwelling Women: What Is Normal? Am. J. Obstet. Gynecol. 2009, 200, 552.e1–552.e7. [Google Scholar] [CrossRef] [Green Version]
- Faustino, M.; Durão, J.; Pereira, C.F.; Pintado, M.E.; Carvalho, A.P. Mannans and Mannan Oligosaccharides (MOS) from Saccharomyces cerevisiae—A Sustainable Source of Functional Ingredients. Carbohydr. Polym. 2021, 272, 118467. [Google Scholar] [CrossRef] [PubMed]
- Spencer, J.D.; Schwaderer, A.L.; Becknell, B.; Watson, J.; Hains, D.S. The Innate Immune Response during Urinary Tract Infection and Pyelonephritis. Pediatr. Nephrol. 2014, 29, 1139–1149. [Google Scholar] [CrossRef] [Green Version]
- Schilling, J.D.; Martin, S.M.; Hunstad, D.A.; Patel, K.P.; Mulvey, M.A.; Justice, S.S.; Lorenz, R.G.; Hultgren, S.J. CD14- and Toll-like Receptor-Dependent Activation of Bladder Epithelial Cells by Lipopolysaccharide and Type 1 Piliated Escherichia coli. Infect. Immun. 2003, 71, 1470–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedges, S.; Agace, W.; Svensson, M.; Sjogren, A.C.; Ceska, M.; Svanborg, C. Uroepithelial Cells Are Part of a Mucosal Cytokine Network. Infect. Immun. 1994, 62, 2315–2321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Man, P.; Van Kooten, C.; Aarden, L.; Engberg, I.; Linder, H.; Svanborg Eden, C. Interleukin-6 Induced at Mucosal Surfaces by Gram-Negative Bacterial Infection. Infect. Immun. 1989, 57, 3383–3388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wullt, B.; Bergsten, G.; Connell, H.; Röllano, P.; Gebratsedik, N.; Hang, L.; Svanborg, C. P-Fimbriae Trigger Mucosal Responses to Escherichia coli in the Human Urinary Tract. Cell. Microbiol. 2001, 3, 255–264. [Google Scholar] [CrossRef]
- Schilling, J.D.; Mulvey, M.A.; Vincent, C.D.; Lorenz, R.G.; Hultgren, S.J. Bacterial Invasion Augments Epithelial Cytokine Responses to Escherichia coli Through a Lipopolysaccharide-Dependent Mechanism. J. Immunol. 2001, 166, 1148–1155. [Google Scholar] [CrossRef] [Green Version]
- Graham, J.C.; Galloway, A. ACP Best Practice No 167: The Laboratory Diagnosis of Urinary Tract Infection. J. Clin. Pathol. 2001, 54, 911–919. [Google Scholar] [CrossRef]
- Jantausch, B.A.; O’Donnell, R.; Wiedermann, B.L. Urinary Interleukin-6 and Interleukin-8 in Children with Urinary Tract Infection. Pediatr. Nephrol. 2000, 15, 236–240. [Google Scholar] [CrossRef]
- Schwartz, D.J.; Conover, M.S.; Hannan, T.J.; Hultgren, S.J. Uropathogenic Escherichia coli Superinfection Enhances the Severity of Mouse Bladder Infection. PLOS Pathog. 2015, 11, e1004599. [Google Scholar] [CrossRef]
- Tramma, D.; Hatzistylianou, M.; Gerasimou, G.; Lafazanis, V. Interleukin-6 and Interleukin-8 Levels in the Urine of Children with Renal Scarring. Pediatr. Nephrol. 2012, 27, 1525–1530. [Google Scholar] [CrossRef] [PubMed]
- Sheu, J.-N.; Chen, M.-C.; Chen, S.-M.; Chen, S.-L.; Chiou, S.-Y.; Lue, K.-H. Relationship between Serum and Urine Interleukin-6 Elevations and Renal Scarring in Children with Acute Pyelonephritis. Scand. J. Urol. Nephrol. 2009, 43, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Sheu, J.N.; Chen, M.C.; Lue, K.H.; Cheng, S.L.; Lee, I.C.; Chen, S.M.; Tsay, G.J. Serum and Urine Levels of Interleukin-6 and Interleukin-8 in Children with Acute Pyelonephritis. Cytokine 2006, 36, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, L.M.; Robles, B.; Marugán, J.M.; Suárez, Á.; Santos, F. Urinary Interleukin-6 Is Useful in Distinguishing between Upper and Lower Urinary Tract Infections. Pediatr. Nephrol. 2008, 23, 429–433. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faustino, M.; Silva, S.; Costa, E.M.; Pereira, A.M.; Pereira, J.O.; Oliveira, A.S.; Ferreira, C.M.H.; Pereira, C.F.; Durão, J.; Pintado, M.E.; et al. Effect of Mannan Oligosaccharides Extracts in Uropathogenic Escherichia coli Adhesion in Human Bladder Cells. Pathogens 2023, 12, 885. https://doi.org/10.3390/pathogens12070885
Faustino M, Silva S, Costa EM, Pereira AM, Pereira JO, Oliveira AS, Ferreira CMH, Pereira CF, Durão J, Pintado ME, et al. Effect of Mannan Oligosaccharides Extracts in Uropathogenic Escherichia coli Adhesion in Human Bladder Cells. Pathogens. 2023; 12(7):885. https://doi.org/10.3390/pathogens12070885
Chicago/Turabian StyleFaustino, Margarida, Sara Silva, Eduardo M. Costa, Ana Margarida Pereira, Joana Odila Pereira, Ana Sofia Oliveira, Carlos M. H. Ferreira, Carla F. Pereira, Joana Durão, Manuela E. Pintado, and et al. 2023. "Effect of Mannan Oligosaccharides Extracts in Uropathogenic Escherichia coli Adhesion in Human Bladder Cells" Pathogens 12, no. 7: 885. https://doi.org/10.3390/pathogens12070885