
Insect Meals and Insect Antimicrobial Peptides as an Alternative for Antibiotics and Growth Promoters in Livestock Production

{kind=link}
Abstract
:1. Introduction
2. Antimicrobial Peptides
3. Source of Antimicrobial Peptides
4. Insect Antimicrobial Peptides
4.1. Defensins (Cysteine-Rich Peptides)
4.2. Cecropins (α-Helical AMPs)
4.3. Moricins
4.4. Proline-Rich Peptides
4.5. Glycine-Rich Peptides
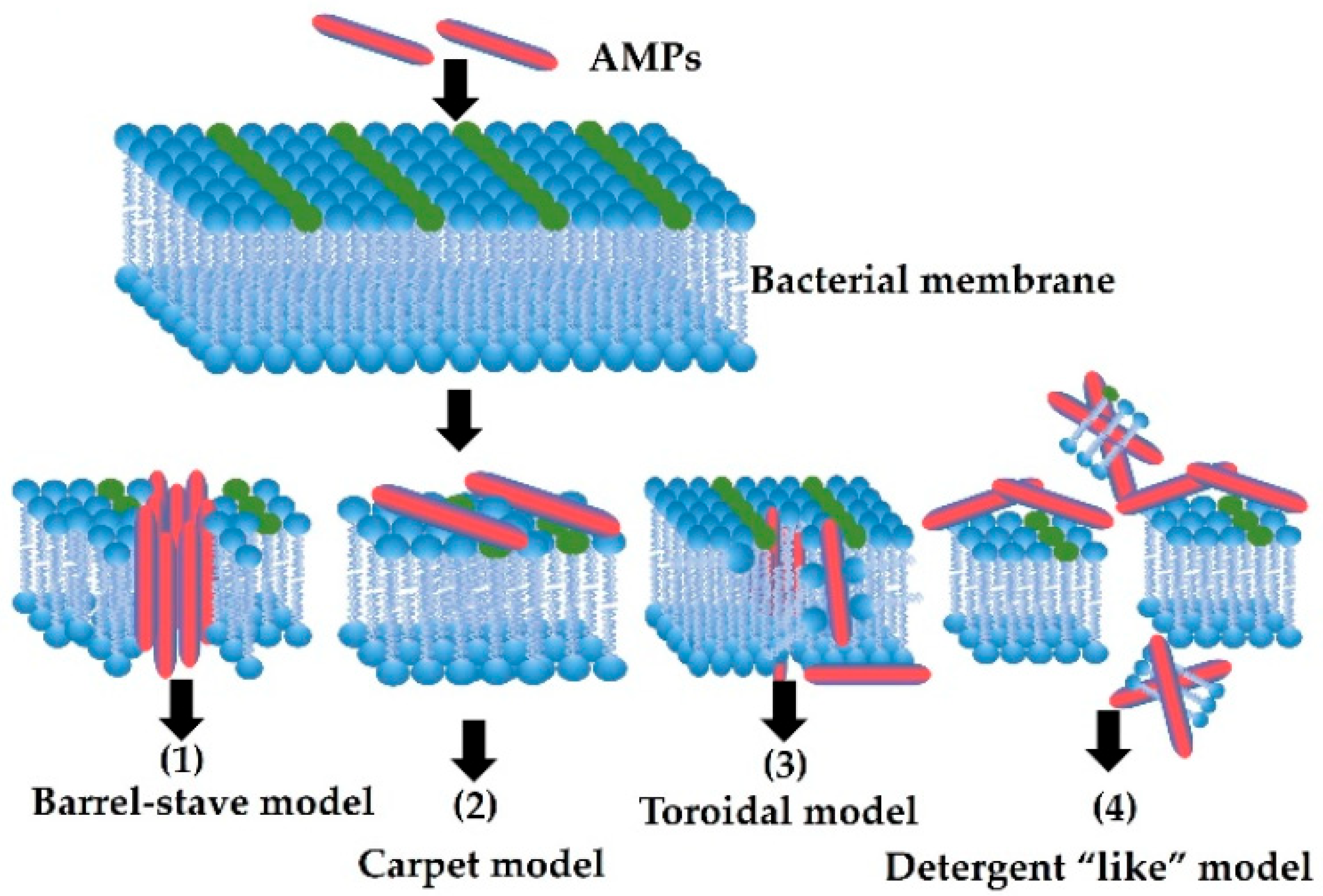
5. Mode of Action of AMPs
6. Mechanisms of Resistance to AMPs
7. Effect of Antimicrobial Peptides on Livestock Health
8. Legal Regulations Regarding the Use of Insects in Feeding Farm Animals
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Osiński, Z.; Patyra, E. Nawozy naturalne i organiczne jako źródło zanieczyszczenia środowiska substancjami przeciwbakteryjnymi. Med. Weter. 2022, 78, 173–183. [Google Scholar]
- Schulze, M.; Nitsche-Melkus, E.; Hensel, B.; Jung, M.; Jakop, U. Antibiotics and their alternatives in Artificial Breeding in livestock. Anim. Reprod. Sci. 2020, 220, 106284. [Google Scholar] [CrossRef] [PubMed]
- Mengistu, G.; Dejenu, G.; Tesema, C.; Arega, B.; Awoke, T.; Alemu, K.; Moges, F. Epidemiology of streptomycin resistant Salmonella from humans and animals in Ethiopia: A systematic review and meta-analysis. PLoS ONE 2020, 15, e0244057. [Google Scholar] [CrossRef] [PubMed]
- Molineri, A.I.; Camussone, C.; Zbrun, M.V.; Suárez Archilla, G.; Cristiani, M.; Neder, V.; Calvinho, L.; Signorini, M. Antimicrobial resistance of Staphylococcus aureus isolated from bovine mastitis: Systematic review and meta-analysis. Prev. Vet. Med. 2021, 188, 105261. [Google Scholar] [CrossRef]
- Józefiak, A.; Enberg, R.M. Insect proteins as a potential source of antimicrobial peptides in livestock production. A review. J. Anim. Feed Sci. 2017, 26, 87–99. [Google Scholar] [CrossRef]
- Wang, G. Human antimicrobial peptides and proteins. Pharmaceuticals 2014, 7, 545–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.K.; Kim, C.; Seo, C.H.; Park, Y. The therapeutic applications of antimicrobial peptides (AMPs): A patent review. J. Microbiol. 2017, 55, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lei, J.; Sun, L.; Huang, S.; Zhu, C.; Li, P.; He, J. The antimicrobial peptides and their potential clinical applications. Am. J. Transl. Res. 2019, 11, 3919–3931. [Google Scholar]
- Mohammed, I.; Said, D.G.; Dua, H.S. Human antimicrobial peptides in ocular surface defense. Prog Retin Eye Res. 2017, 61, 1–22. [Google Scholar] [CrossRef]
- Giuliani, A.; Pirri, G.; Nicoletto, S.F. Antimicrobial peptides: An overview of a promising class of therapeutics. Cent. Eur. J. Biol. 2007, 2, 1–33. [Google Scholar] [CrossRef]
- Yount, N.Y.; Yeaman, M.R. Structural congruence among membrane-active host defense polypeptides of diverse phylogeny. Biochim. Et Biophys. Acta (BBA)-Biomembr. 2006, 1758, 1373–1386. [Google Scholar] [CrossRef] [Green Version]
- Dathe, M.; Wieprecht, T. Structural features of helical antimicrobial peptides: Their potential to modulate activity on model membranes and biological cells. Biochim. Biophys. Acta. 1999, 1462, 71–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Available online: http://dramp.cpu-bioinfor.org/ (accessed on 18 May 2023).
- Kang, X.; Dong, F.; Shi, C.; Liu, S.; Sun, J.; Chen, J. DRAMP 2.0, An updated data repository of antimicrobial peptides. Sci. Data. 2019, 6, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.-Y.; Yan, Z.-B.; Meng, Y.-M.; Hong, X.-Y.; Shao, G.; Ma, J.-J.; Cheng, X.-R.; Liu, J.; Kang, J.; Fu, C.-Y. Antimicrobial peptides: Mechanism of action, activity and clinical potential. Mil. Med. Res. 2021, 8, 48. [Google Scholar] [CrossRef]
- Hancock, R.E.W.; Scott, G. The role of antimicrobial peptides in animal defense. Proc. Natl. Acad. Sci. USA 2000, 97, 8856–8861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hancock, R.E.; Brown, K.L.; Mookherjee, N. Host defence peptides from invertebrates: Emerging antimicrobial strategies. Immunobiology 2006, 211, 315–322. [Google Scholar] [CrossRef]
- Harris, F.; Dennison, S.; Phoenix, D.A. Anionic Antimicrobial Peptides from Eukaryotic Organisms. Curr. Protein Pept. Sci. 2009, 10, 585–606. [Google Scholar] [CrossRef]
- Torensello, A.L.; Borrelli, A.; Buonaguro, L.; Buonaguro, F.M.; Tornsello, M.L. Antimicrobial Peptides as Anticancer Agents: Functional Properties and Biological Activities. Molecules 2020, 25, 2850. [Google Scholar] [CrossRef]
- Zasloff, M. Magainins, a class of antimicrobial peptides from Xenopus. skin: Isolation, characterization of two active forms, and partial cDNA sequence of a precursor. Proc. Natl. Acad. Sci. USA 1987, 84, 5449–5453. [Google Scholar] [CrossRef] [Green Version]
- Jin, G.; Weinberg, A. Human antimicrobial peptides and cancer. Semin. Cell Dev. Biol. 2019, 88, 156–162. [Google Scholar] [CrossRef]
- Błaszczyk, U.; Moczarny, J. Bakteriocyny bakterii Gram-ujemnych–struktura, mechanizm działania i zastosowanie. Postępy Mikrobiologii. 2016, 2, 157–171. [Google Scholar]
- Zhang, C.; Yang, M. Antimicrobial Peptides: From Design to Clinical Application. Antibiotics 2022, 11, 349. [Google Scholar] [CrossRef] [PubMed]
- Berrocal-Lobo, M.; Molina, A.; Rodriguez-Palenzuela, P.; Garcia-Olmedo, F.; Rivas, L. Leishmania donovani: Thionins, plant antimicrobial peptides with leishmanicidal activity. Exp. Parasitol. 2009, 122, 247–249. [Google Scholar] [CrossRef] [Green Version]
- Dass, M.; White, M.R.; Tecle, T.; Hartsharn, K.L. Human defensines and LL-37 in mucosal immunity. J. Leukoc. Biol. 2010, 87, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Cytryńska, M. O odporności bez przeciwciał. Postępy Biol. Komórki. 2009, 36, 309–324. [Google Scholar]
- Lee, Y.S.; Yun, E.K.; Jang, W.S.; Kim, I.; Lee, J.H.; Park, S.Y.; Ryu, K.S.; Seo, S.J.; Kim, C.H.; Lee, I.H. Purification, cDNA cloning and expression of an insect defensin from the great wax moth, Galleria mellonella. Insect Mol. Biol. 2004, 13, 65–72. [Google Scholar] [CrossRef]
- Lehane, M.J.; Wu, D.; Lehane, S.M. Midgut-specific immune molecules are produced by the blood-sucking insect Stomoxys calcitrans. Proc. Natl. Acad. Sci. USA 1997, 94, 11502–11507. [Google Scholar] [CrossRef] [Green Version]
- Zdybicka-Barabas, A.; Bulak, P.; Polakowski, C.; Bieganowski, A.; Waśko, A.; Cytryńska, M. Immune response in the larvae of the black soldier fly Hermetia illucens. ISJ-Invertebr. Surviv. J. 2017, 14, 9–17. [Google Scholar]
- Boman, H.G.; Steiner, H. Humoral immunity in Cecropia pupae. Curr. Top. Microbiol. Immunol. 1981, 94–95, 75–91. [Google Scholar]
- Vallet-Gely, I.; Lemaitre, B.; Boccard, F. Bacterial strategies to overcome insect defences. Nat. Rev. Microbiol. 2008, 6, 302–313. [Google Scholar] [CrossRef] [Green Version]
- Martens, E.; Demain, A.L. The antibiotic resistance crisis, with a focus on the United States. J. Antibiot. 2017, 70, 520–526. [Google Scholar] [CrossRef] [Green Version]
- Kanost, M.R.; Jiang, H.; Yu, X.Q. Innate immune responses of a lepidopteran insect, Manduca sexta. Immunol. Rev. 2004, 198, 97–105. [Google Scholar] [CrossRef]
- Hultmark, D.; Steiner, H.; Rasmuson, T.; Boman, H.G. Insect immunity. Purification and properties of three inducible bactericidal proteins from hemolymph of immunized pupae of Hyalophora cecropia. Eur. J. Biochem. 1980, 106, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Steiner, H.; Hultmark, D.; Engström, Å.; Bennich, H.; Boman, H.G. Sequence and specificity of two antibacterial proteins involved in insect immunity. Nature 1981, 292, 246–248. [Google Scholar] [CrossRef]
- Sultana, A.; Luo, H.; Ramakrishna, S. Harvesting of Antibacterial Peptides from Insect (Hermetia illucens) and Its Applications in the Food Packaging. Appl. Sci. 2021, 11, 6991. [Google Scholar] [CrossRef]
- Koehbach, J. Structure-Activity Relationships of Insect Defensins. Front. Chem. 2017, 5, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimarcq, J.-L.; Bulet, P.; Hetru, C.; Hoffman, J. Cysteine-Rich Antimicrobial Peptides in Invertebrates. Pept. Sci. 1998, 47, 465–477. [Google Scholar] [CrossRef]
- Otvos, L., Jr.; Rogers, I.O.M.; Consolvo, P.J.; Condie, B.A.; Lovas, S.; Blaszczyk-Thurin, M. Interaction between heat shock proteins and antimicrobial peptides. Biochemistry 2000, 21, 14150–14159. [Google Scholar] [CrossRef]
- Torres, A.M.; Kuchel, P.W. The beta-defensin-fold family of polypeptides. Toxicon 2004, 44, 581–588. [Google Scholar] [CrossRef]
- Gao, B.; Zhu, S. An insect defensin-derived B-hairpin peptide with enhanced antibacterial activity. ACS Chem Biol. 2014, 9, 405–413. [Google Scholar] [CrossRef]
- Yi, H.Y.; Chowdhury, M.; Huang, Y.D.; Yu, X.Q. Insect antimicrobial peptides and their application. Appl. Microbiol. Biotechnol. 2014, 5, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Ratcliffe, N.A.; Gotz, P. Functional studies on insect haemocytes, including non-self recognition. Res. ImmunoI. 1990, 141, 919–923. [Google Scholar] [CrossRef] [PubMed]
- Bulet, P.; Charlet, M.; Hetru, C. Antimicrobial Peptides in Insect Immunity. In Innate Immunity; Alan, R., Ezekowitz, B., Hoffamn, J.A., Eds.; Humana Press: Totowa, NJ, USA, 2003. [Google Scholar]
- Van Hofsten, P.; Faye, I.; Kockum, K.; Lee, J.Y.; Xanthopoulos, K.G.; Boman, I.A. Molecular cloning, cDNA sequencing, and chemical synthesis of cecropin B from Hyalophora cecropia. Proc. Nat. Acad. Sci. USA 1985, 82, 2240–2243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, A.J.; Beazley, W.D.; Bibby, M.C.; Devine, D.A. Antimicrobial activity of cecropins. J. Antimicrobiol. Chemother. 1996, 37, 1077–1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bechinger, B.; Lohner, K. Detergent-like actions of linear amphipathic cationic antimicrobial peptides. Biochim. Biophys. Acta 2006, 1758, 1529–1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouyang, L.; Xu, X.; Freed, S.; Gao, Y.; Yu, J.; Wang, S.; Ju, W.; Zhang, Y.; Jin, F. Cecropins from Plutella xylostella and Their Interaction with Metarhizium anisopliae. PLoS ONE 2015, 10, e0142451. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.; Lee, D.G. Cecropin A-induced apoptosis is regulated by ion balance and glutathione antioxidant system in Candida albicans. IUBMB Life 2016, 68, 652–662. [Google Scholar] [CrossRef] [Green Version]
- Lu, D.; Geng, T.; Hou, C.; Huang, Y.; Qin, G.; Guo, X. Bombyx mori Cecropin A has a high antifungal activity to entomopathogenic fungus Beauveria bassiana. Gene 2016, 583, 29–35. [Google Scholar] [CrossRef]
- Giacometti, A.; Cirioni, O.; Ghiselli, R.; Viticchi, C.; Mocchegiani, F.; Riva, A.; Saba, V.; Scalise, G. Effect of mono-dose intraperitoneal cecropins in experimental septic shock. Crit. Care Med. 2001, 29, 1666–1669. [Google Scholar] [CrossRef]
- Arrowood, M.J.; Jaynes, J.M.; Healey, M.C. In vitro activities of lytic peptides against the sporozoites of Cryptosporidium parvum. Antimicrob. Agents Chemother. 1991, 35, 224–227. [Google Scholar] [CrossRef] [Green Version]
- Barr, S.C.; Rose, D.; Jaynes, J.M. Activity of lytic peptides against intracellular Trypanosoma cruzi amastigotes in vitro and parasitemias in mice. J. Parasitol. 1995, 81, 974–978. [Google Scholar] [CrossRef]
- Boulanger, N.; Brun, R.; Ehret-Sabatier, L.; Kunz, C.; Bulet, P. Immunopeptides in the defense reactions of Glossina morsitans to bacterial and Trypanosoma brucei brucei infections. Insect Biochem. Mol. Biol. 2002, 32, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Wachinger, M.; Kleinschmidt, A.; Winder, D.; von Pechmann, N.; Ludvigsen, A.; Neumann, M.; Holle, R.; Salmons, B.; Erfle, V.; Brack-Werner, R. Antimicrobial peptides melittin and cecropin inhibit replication of human immunodeficiency virus 1 by suppressing viral gene expression. J. Gen. Virol. 1998, 79, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Orivel, J.; Redeker, V.; Le Caer, J.P. Ponericins, new antibacterial and insecticidal peptides from the venom of the ant Pachycondyla goeldii. J. BioI. Chern. 2001, 276, 17823–17829. [Google Scholar] [CrossRef] [Green Version]
- Hara, S.; Yamakawa, M.M. A novel type of antibacterial peptide isolated from the silkworm, Bombyx mori. J. Bloch. Chem. 1995, 270, 29923–29927. [Google Scholar] [CrossRef] [Green Version]
- Dai, H.; Rayaprolu, S.; Gong, Y.; Huang, R.; Prakash, O.; Jiang, H. Solution structure, antibacterial activity, and expression profile of Manduca sexta moricin. J. Pept. Sci. 2008, 14, 855–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara, S.; Yamakawa, M. A novel antibacterial peptide family isolated from the silkworm, Bombyx mori. Blochem. J. 1995, 310, 651–656. [Google Scholar] [CrossRef] [Green Version]
- Scocchi, M.; Tossi, A.; Gennaro, R. Proline-rich antimicrobial peptides: Converging to a non-lytic mechanism of action. Cell. Mol. Life Sci. 2011, 68, 2317–2330. [Google Scholar] [CrossRef]
- Veldkamp, T.; Dong, L.; Paul, A.; Govers, C. Bioactive properties of insect products for monogastric animals—A review. J. Insects Food Feed 2022, 9, 1027–1040. [Google Scholar] [CrossRef]
- Kragol, G.; Lovas, S.; Varadi, G.; Condie, B.A.; Hoffmann, R.; Otvos, L., Jr. The antibacterial peptide pyrrhocoricin inhibits the ATPase actions of DnaK and prevents chaperone-assisted protein folding. Biochemistry 2001, 40, 3016–3026. [Google Scholar] [CrossRef]
- Wu, Q.; Patocka, J.; Kuca, K. Insect Antimicrobial Peptides, a Mini Review. Toxins 2018, 10, 461. [Google Scholar] [CrossRef]
- Ongey, E.L.; Pflugmacher, S.; Neubauer, P. Bioinspired designs, molecular premise and tools for evaluating the ecological importance of antimicrobial peptides. Pharmaceuticals 2018, 11, 68. [Google Scholar] [CrossRef] [Green Version]
- Silvestro, L.; Axelsen, P.H. Membrane-induced folding of cecropin A. Biophys. J. 2000, 79, 1465–1477. [Google Scholar] [CrossRef] [Green Version]
- Pálffy, R.; Gardlík, R.; Behuliak, M.; Kadasi, L.; Turna, J.; Celec, P. On the physiology and pathophysiology of antimicrobial peptides. Mol. Med. 2009, 15, 51–59. [Google Scholar] [CrossRef]
- Brodgen, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar]
- Ragheb, M.N.; Thomason, M.K.; Hsu, C.; Nugent, P.; Gage, J.; Samadpour, A.N.; Karissa, A.; Merrikh, C.N.; Miller, S.I.; Sherman, D.R.; et al. Inhibiting the Evaluation of Antibiotic Resistance. Mol. Cell. 2019, 73, 157–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munoz Madero, C.; Lopez Navas, A.; Padila Leon, E.; Alvarez, A.S. Use of antibiotic with ONE HEALTH perspective: Spanish Strategic Action Plan to reduce the risk of selection and dissemination of antibiotic resistance. Small Rumin. Res. 2016, 142, 44–47. [Google Scholar] [CrossRef]
- Kang, K.M.; Park, J.H.; Kim, S.H.; Yang, S.J. Potetntial role of host defense antimicrobial peptide resistance in increased virulence of health care-associated MRSA strains of sequence type (ST) versus livestock-associated and community-associated MRSA strains of ST72. Comp. Immunol. Microbiol. Infect. Dis. 2019, 62, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Nuri, R.; Shprung, T.; Shai, Y. Defensive remodeling: How bacteria Surface properties and biofilm formation promote resistance to antimicrobial peptides. Biochem. Biophys. Acta-Biomemb. 2015, 1848, 3089–3100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peschel, A.; Sahl, H.-G. The co-evolution of host cationic antimicrobial peptides and microbial resistance. Nat. Rev. Microbiol. 2006, 4, 529–536. [Google Scholar] [CrossRef]
- Fedtke, I.; Gotz, F.; Peschel, A. Bacterial evasion of innate host defense-the Staphylococcus aureus lesson. Int. J. Med. Microbiol. 2004, 294, 189–194. [Google Scholar] [CrossRef]
- Moranta Mesquida, D.; Regueiro, V.; March, C.; Llobet, E.; Margareto, J.; Larrate, E.; Garmendia, J.; Bengoechea, J.A. Klebsiella pneumoniae capsule polisaccharide impedes the expression of beta-defensins by airway epithelial cells. Infect. Immun. 2010, 78, 1135–1146. [Google Scholar] [CrossRef] [Green Version]
- Schmidtchen, A.; Frick, I.M.; Andersson, E.; Tapper, H.; Bjorck, L. Proteinases of common pathogenic bacteria degrade and inactivate the antibacterial peptide LL-37. Mol. Microbiol. 2002, 46, 157–168. [Google Scholar] [CrossRef] [Green Version]
- Mason, K.M.; Munson RSJr Bakaletz, L.O. A mutation in the sap operon attenuates survival of nontypeable Haemophilus infuezae in a Chinchilla model of otitis media. Infect. Immun. 2005, 73, 599–608. [Google Scholar] [CrossRef] [Green Version]
- Gasco, L.; Józefiak, A.; Henry, M. Beyond the protein concept: Health aspects of using edible insects on animals. J. Insects Food Feed 2020, 7, 715–741. [Google Scholar] [CrossRef]
- Xiao, H.; Tan, B.E.; Wu, M.M. Effects of composite antimicrobial peptides in weanling piglets challenged withdeoxynivalenol: II. Intestinal morphology and function. J. Anim. Sci. 2013, 91, 4750–4756. [Google Scholar] [CrossRef] [PubMed]
- Józefiak, A.; Kierończyk, B.; Rawski, M.; Mazurkiewicz, J.; Benzertiha, A.; Gobbi, P.; Nogales-Merida, S.; Świątkiewicz, A.; Józefiak, D. Full-fat insect meals as feeds additive–the effect on broiler chicken growth performance and gastroinestinal tract microbiota. J. Anim. Feed Sci. 2018, 27, 131–139. [Google Scholar] [CrossRef]
- Islam, M.M.; Yang, C.-J. Efficacy of mealworm and super mealworm larvae probiotics as an alternative to antibiotics challenged orally with Salmonella and E. coli infection in broiler chicks. Poult. Sci. 2017, 96, 27–34. [Google Scholar] [CrossRef]
- Ji, Y.J.; Liu, H.N.; Kong, X.F.; Blachier, F.; Geng, M.M.; Liu, Y.Y. Use of insect powder as a source of dietary protein in early-weaned piglets. J. Anim. Sci. 2016, 94, 111–116. [Google Scholar] [CrossRef] [Green Version]
- Lai, Y.; Gallo, R.L. AMPed up immunity: How antimicrobial peptides have multiple roles in immune defense. Trends Immunol. 2009, 30, 131–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Zeng, X.; Yang, Q.; Qiao, S. Antimicrobial Peptides as Potential Alternatives to Antibiotics in Food Animal Industry. Int. J. Mol. Sci. 2016, 17, 603. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.F.; Wu, G.Y.; Liao, Y.P.; Hou, Z.P.; Liu, H.J.; Yin, F.G. Effects of Chinese herbal ultra-fine powder as a dietary additive on growth performance, serum metabolites and intestinal health in early-weaned piglets. Livest. Sci. 2007, 108, 272–275. [Google Scholar] [CrossRef]
- Chaturvedi, P.; Bhat, R.A.H.; Pande, A. Antimicrobial peptides of fish: Innocuous alternatives to antibiotics. Rev. Aquacult. 2020, 12, 85–106. [Google Scholar] [CrossRef]
- Horner, K.; Drummond, E.; Brennan, L. Bioavailability of milk protein-derived bioactive peptides: A glycaemic management perspective. Nutr. Res. Rev. 2016, 29, 91–101. [Google Scholar] [CrossRef] [Green Version]
- Aoki, W.; Ueda, M. Characterization of Antimicrobial Peptides toward the Development of Novel Antibiotics. Pharmaceuticals 2013, 6, 1055–1081. [Google Scholar] [CrossRef] [Green Version]
- López-García, G.; Dublan-García, O.; Arizmendi-Cotero, D.; Gómez Oliván, L.M. Antioxidant and Antimicrobial Peptides Derived from Food Proteins. Molecules 2022, 27, 1343. [Google Scholar] [CrossRef]
- Commission Regulation (EU) 2017/893 of 24 May 2017 amending Annexes I and IV to Commission Regulation (EC) No 999/2001 of the European Parliament and of the Council and Annexes X, XIV and XV to Commission Regulation (EU) No 142/2011 as Regards the Provisions on Processed Animal Protein. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32017R0893 (accessed on 3 January 2023).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patyra, E.; Kwiatek, K. Insect Meals and Insect Antimicrobial Peptides as an Alternative for Antibiotics and Growth Promoters in Livestock Production. Pathogens 2023, 12, 854. https://doi.org/10.3390/pathogens12060854
Patyra E, Kwiatek K. Insect Meals and Insect Antimicrobial Peptides as an Alternative for Antibiotics and Growth Promoters in Livestock Production. Pathogens. 2023; 12(6):854. https://doi.org/10.3390/pathogens12060854
Chicago/Turabian StylePatyra, Ewelina, and Krzysztof Kwiatek. 2023. "Insect Meals and Insect Antimicrobial Peptides as an Alternative for Antibiotics and Growth Promoters in Livestock Production" Pathogens 12, no. 6: 854. https://doi.org/10.3390/pathogens12060854