
Bis-Benzylisoquinoline Alkaloids Inhibit Porcine Epidemic Diarrhea Virus by Disrupting Virus Entry

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Antibody and Reagents
2.3. PEDV Propagation
2.4. Cytotoxicity Assay
2.5. Determination of the Half Maximal Inhibitory Concentration (IC50) of Compounds
2.6. The Effect of the Compounds on PEDV Binding and Entry
2.7. RNA Extraction and RT-PCR
2.8. Western Blot Analysis
2.9. Immunofluorescence Analysis
2.10. Plaque Formation Assay (PFU)
2.11. Lysosomal pH Measurement
2.12. Determining the Activity of CTSL and CTSB
2.13. Statistical Analysis
3. Results
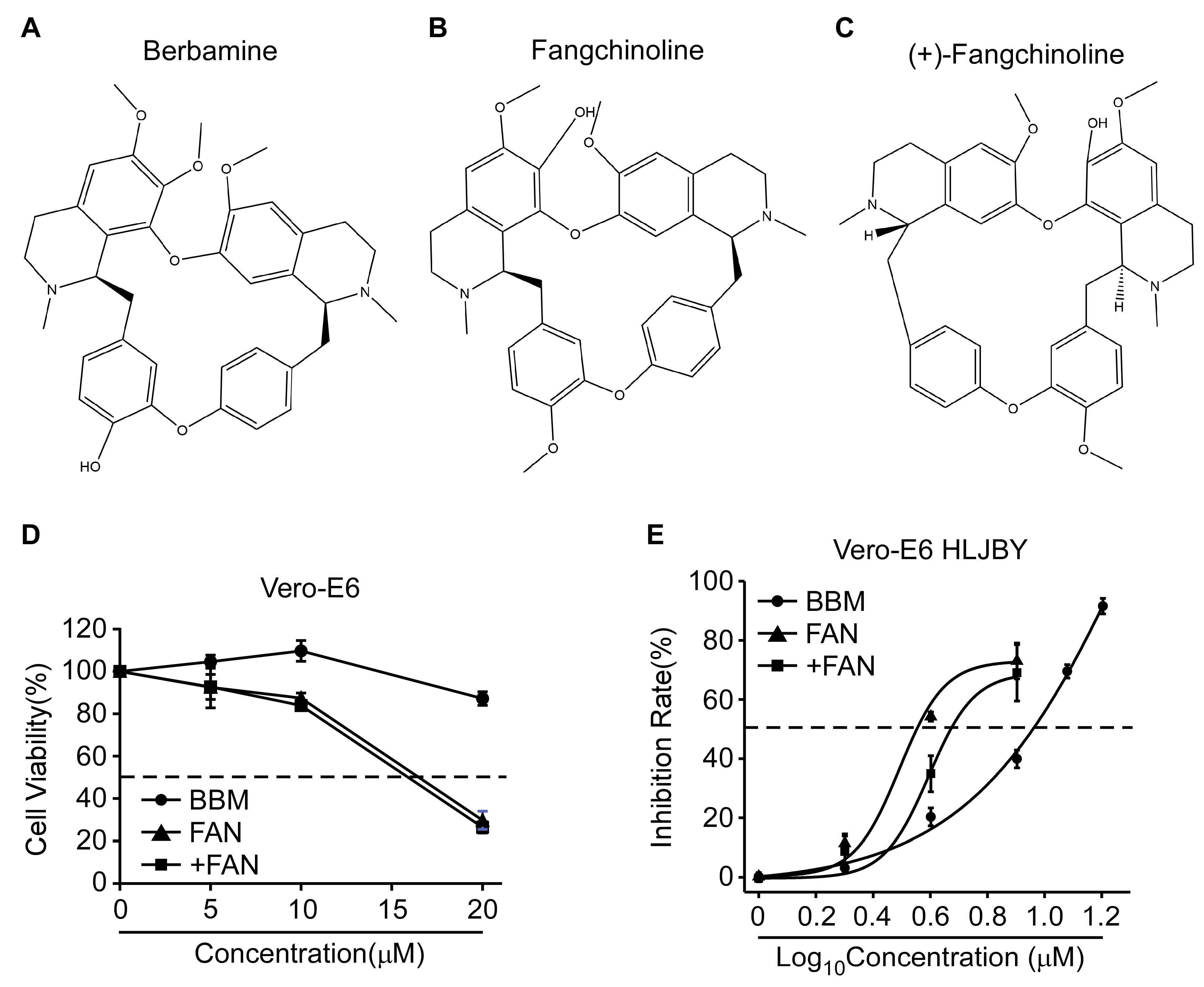
3.1. The 50% Cytotoxicity Concentration (CC50) and IC50 Values of BBM, FAN, and +FAN in Vero-E6 Cells
3.2. PEDV Infection Was Inhibited by BBM, FAN, and +FAN In Vitro
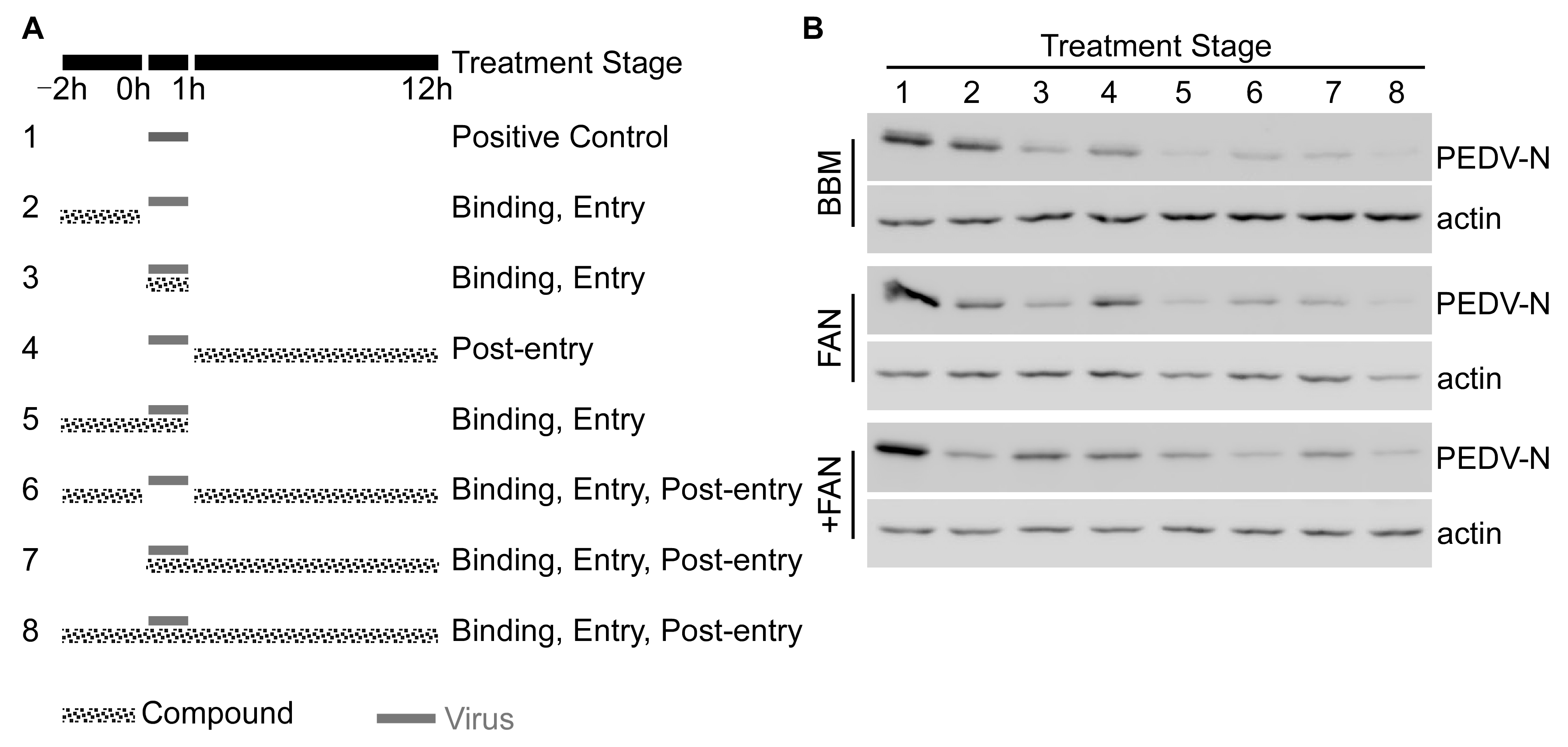
3.3. BBM, FAN, and +FAN Inhibited PEDV Infection Mainly at the Early Stage
3.4. BBM, FAN, and +FAN Did Not Target PEDV Particles Directly
3.5. BBM, FAN, and +FAN Inhibited PEDV Entry by Suppressing Lysosome Acidification
3.6. BBM, FAN, and +FAN Decreased the Activity of CTSL and CTSB
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, X.; Wang, Y.; Lu, P.; Shen, Y.; Zhao, X.; Zhu, Y.; Jiang, Z.; Yang, H.; Pan, H.; Zhao, L.; et al. Pebp1 Suppresses HIV Transcription and Induces Latency by Inactivating Mapk/Nf-Kappab Signaling. EMBO Rep. 2020, 21, e49305. [Google Scholar] [CrossRef]
- Lin, F.; Zhang, H.; Li, L.; Yang, Y.; Zou, X.; Chen, J.; Tang, X. PEDV: Insights and Advances into Types, Function, Structure, and Receptor Recognition. Viruses 2022, 14, 1744. [Google Scholar] [CrossRef]
- Shi, P.; Su, Y.; Li, R.; Liang, Z.; Dong, S.; Huang, J. PEDV Nsp16 Negatively Regulates Innate Immunity to Promote Viral Proliferation. Virus Res. 2019, 265, 57–66. [Google Scholar] [CrossRef]
- Wei, X.; She, G.; Wu, T.; Xue, C.; Cao, Y. PEDV Enters Cells through Clathrin-, Caveolae-, and Lipid Raft-Mediated Endocytosis and Traffics via the Endo-/Lysosome Pathway. Vet. Res. 2020, 51, 10. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Ma, Y.; Yang, Y.; Zheng, Y.; Shang, J.; Zhou, Y.; Jiang, S.; Du, L.; Li, J.; Li, F. Cell Entry of Porcine Epidemic Diarrhea Coronavirus Is Activated by Lysosomal Proteases. J. Biol. Chem. 2016, 291, 24779–24786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, C.; Kim, Y.; Chang, K.O. Proteases Facilitate the Endosomal Escape of Porcine Epidemic Diarrhea Virus during Entry into Host Cells. Virus Res. 2019, 272, 197730. [Google Scholar] [CrossRef]
- Shivanna, V.; Kim, Y.; Chang, K.O. Endosomal Acidification and Cathepsin L Activity Is Required for Calicivirus Replication. Virology 2014, 464–465, 287–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, M.C.; Solania, A.; Jiang, Z.; Christy, M.P.; Podvin, S.; Mosier, C.; Lietz, C.B.; Ito, G.; Gerwick, W.H.; Wolan, D.W.; et al. Selective Neutral Ph Inhibitor of Cathepsin B Designed Based on Cleavage Preferences at Cytosolic and Lysosomal Ph Conditions. ACS Chem. Biol. 2021, 16, 1628–1643. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.; Saif, L.J.; Wang, Q. Porcine Epidemic Diarrhea Virus (PEDV): An Update on Etiology, Transmission, Pathogenesis, and Prevention and Control. Virus Res. 2020, 286, 198045. [Google Scholar] [CrossRef]
- Su, M.; Li, C.; Qi, S.; Yang, D.; Jiang, N.; Yin, B.; Guo, D.; Kong, F.; Yuan, D.; Feng, L.; et al. A Molecular Epidemiological Investigation of PEDV in China: Characterization of Co-Infection and Genetic Diversity of S1-Based Genes. Transbound. Emerg. Dis. 2020, 67, 1129–1140. [Google Scholar] [CrossRef] [Green Version]
- Lee, C. Porcine Epidemic Diarrhea Virus: An Emerging and Re-Emerging Epizootic Swine Virus. Virol. J. 2015, 12, 193. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Li, S.; Song, K.; Ye, J.; Li, W.; Zhong, Y.; Feng, Z.; Liang, S.; Cai, Z.; Xu, K. A Broad Antiviral Strategy: Inhibitors of Human DHODH Pave the Way for Host-Targeting Antivirals against Emerging and Re-Emerging Viruses. Viruses 2022, 14, 928. [Google Scholar] [CrossRef]
- Xu, W.; Chen, S.; Wang, X.; Wu, H.; Yamada, H.; Hirano, T. Bisbenzylisoquinoline Alkaloids and P-Glycoprotein Function: A Structure Activity Relationship Study. Bioorg. Med. Chem. 2020, 28, 115553. [Google Scholar] [CrossRef]
- Farooqi, A.A.; Wen, R.; Attar, R.; Taverna, S.; Butt, G.; Xu, B. Regulation of Cell-Signaling Pathways by Berbamine in Different Cancers. Int. J. Mol. Sci. 2022, 23, 2758. [Google Scholar] [CrossRef]
- Jia, X.J.; Li, X.; Wang, F.; Liu, H.Q.; Zhang, D.J.; Chen, Y. Berbamine Exerts Anti-Inflammatory Effects Via Inhibition of Nf-Kappab and Mapk Signaling Pathways. Cell. Physiol. Biochem. 2017, 41, 2307–2318. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; Song, Y.; Zhan, Y.; Zhou, S.; Ke, J.; Ao, W.; Zhang, Y.; Liang, Q.; He, M.; Li, S.; et al. Fangchinoline Inhibits Non-Small Cell Lung Cancer Metastasis by Reversing Epithelial-Mesenchymal Transition and Suppressing the Cytosolic ROS-Related Akt-mTOR Signaling Pathway. Cancer Lett. 2022, 543, 215783. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Yuen, T.T.; Ye, Z.; Liu, S.; Zhang, G.; Chu, H.; Yue, J. Berbamine Inhibits Sars-Cov-2 Infection by Compromising TRPMLs-Mediated Endolysosomal Trafficking of ACE2. Signal Transduct. Target. Ther. 2021, 6, 168. [Google Scholar] [CrossRef] [PubMed]
- Gunaratne, G.S.; Yang, Y.; Li, F.; Walseth, T.F.; Marchant, J.S. Naadp-Dependent Ca2+ Signaling Regulates Middle East Respiratory Syndrome-Coronavirus Pseudovirus Translocation through the Endolysosomal System. Cell Calcium 2018, 75, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Li, H.; Ye, Z.; Xu, Q.; Fu, Q.; Sun, W.; Qi, W.; Yue, J. Berbamine Inhibits Japanese Encephalitis Virus (JEV) Infection by Compromising TPRMLs-Mediated Endolysosomal Trafficking of Low-Density Lipoprotein Receptor (LDLR). Emerg. Microbes Infect. 2021, 10, 1257–1271. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Huang, L.; Gao, F.; Jian, W.; Chen, H.; Liao, M.; Qi, W. Berbamine Hydrochloride Inhibits African Swine Fever Virus Infection In Vitro. Molecules 2022, 28, 170. [Google Scholar] [CrossRef]
- Yi, D.; Li, Q.; Wang, H.; Lv, K.; Ma, L.; Wang, Y.; Wang, J.; Zhang, Y.; Liu, M.; Li, X.; et al. Repurposing of Berbamine Hydrochloride to Inhibit Ebola Virus by Targeting Viral Glycoprotein. Acta Pharm. Sin. B 2022, 12, 4378–4389. [Google Scholar] [CrossRef]
- Wan, Z.; Lu, Y.; Liao, Q.; Wu, Y.; Chen, X. Fangchinoline Inhibits Human Immunodeficiency Virus Type 1 Replication by Interfering with Gp160 Proteolytic Processing. PLoS ONE 2012, 7, e39225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.E.; Min, J.S.; Jang, M.S.; Lee, J.Y.; Shin, Y.S.; Song, J.H.; Kim, H.R.; Kim, S.; Jin, Y.H.; Kwon, S. Natural Bis-Benzylisoquinoline Alkaloids-Tetrandrine, Fangchinoline, and Cepharanthine, Inhibit Human Coronavirus Oc43 Infection of Mrc-5 Human Lung Cells. Biomolecules 2019, 9, 696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Chen, H.; Ming, X.; Bo, Z.; Shin, H.J.; Jung, Y.S.; Qian, Y. Porcine Epidemic Diarrhea Virus Infection Induces Caspase-8-Mediated G3bp1 Cleavage and Subverts Stress Granules to Promote Viral Replication. J. Virol. 2021, 95, e02344-20. [Google Scholar] [CrossRef] [PubMed]
- Zhai, X.; Wang, N.; Jiao, H.; Zhang, J.; Li, C.; Ren, W.; Reiter, R.J.; Su, S. Melatonin and Other Indoles Show Antiviral Activities against Swine Coronaviruses In Vitro at Pharmacological Concentrations. J. Pineal Res. 2021, 71, e12754. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Yu, R.; Wang, X.; Chen, B.; Si, F.; Zhou, J.; Xie, C.; Li, Z.; Zhang, D. Bis-Benzylisoquinoline Alkaloids Inhibit Porcine Epidemic Diarrhea Virus In Vitro and In Vivo. Viruses 2022, 14, 1231. [Google Scholar] [CrossRef]
- Kausar, S.; Khan, F.S.; Rehman, M.I.M.U.; Akram, M.; Riaz, M.; Rasool, G.; Khan, A.H.; Saleem, I.; Shamim, S.; Malik, A. A Review: Mechanism of Action of Antiviral Drugs. Int. J. Immunopathol. Pharmacol. 2021, 35, 20587384211002621. [Google Scholar] [CrossRef]
- Koyama, A.H.; Uchida, T. The Effect of Ammonium Chloride on the Multiplication of Herpes Simplex Virus Type 1 In Vero Cells. Virus Res. 1989, 13, 271–281. [Google Scholar] [CrossRef]
- Antas, M.; Woźniakowski, G. Current Status of Porcine Epidemic Diarrhoea (Ped) in European Pigs. J. Vet. Res. 2019, 63, 465–470. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.; Ma, J.; Wang, Y.; Wang, M.; Song, W.; Zhang, W.; Lu, C.; Yao, H. Genomic and Epidemiological Characteristics Provide New Insights into the Phylogeographical and Spatiotemporal Spread of Porcine Epidemic Diarrhea Virus in Asia. J. Clin. Microbiol. 2015, 53, 1484–1492. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Liang, D.; Zheng, Q.; Zhao, M.; Lv, R.; Tang, J.; Chen, N. Berbamine Dihydrochloride Suppresses the Progression of Colorectal Cancer via RTKs/Akt Axis. J. Ethnopharmacol. 2023, 303, 116025. [Google Scholar] [CrossRef] [PubMed]
- He, C.L.; Huang, L.Y.; Wang, K.; Gu, C.J.; Hu, J.; Zhang, G.J.; Xu, W.; Xie, Y.H.; Tang, N.; Huang, A.L. Identification of Bis-Benzylisoquinoline Alkaloids as SARS-CoV-2 Entry Inhibitors from a Library of Natural Products. Signal Transduct. Target. Ther. 2021, 6, 131. [Google Scholar] [CrossRef] [PubMed]
- Saurav, S.; Tanwar, J.; Ahuja, K.; Motiani, R.K. Dysregulation of Host Cell Calcium Signaling during Viral Infections: Emerging Paradigm with High Clinical Relevance. Mol. Aspects Med. 2021, 81, 101004. [Google Scholar] [CrossRef] [PubMed]
- Simmons, G.; Gosalia, D.N.; Rennekamp, A.J.; Reeves, J.D.; Diamond, S.L.; Bates, P. Inhibitors of Cathepsin L Prevent Severe Acute Respiratory Syndrome Coronavirus Entry. Proc. Natl. Acad. Sci. USA 2005, 102, 11876–11881. [Google Scholar] [CrossRef] [Green Version]
- Huang, I.C.; Bosch, B.J.; Li, F.; Li, W.; Lee, K.H.; Ghiran, S.; Vasilieva, N.; Dermody, T.S.; Harrison, S.C.; Dormitzer, P.R.; et al. Sars Coronavirus, but Not Human Coronavirus Nl63, Utilizes Cathepsin L to Infect Ace2-Expressing Cells. J. Biol. Chem. 2006, 281, 3198–3203. [Google Scholar] [CrossRef] [Green Version]
- Kleine-Weber, H.; Elzayat, M.T.; Hoffmann, M.; Pohlmann, S. Functional Analysis of Potential Cleavage Sites in the MERS-Coronavirus Spike Protein. Sci. Rep. 2018, 8, 16597. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.M.; Yang, W.L.; Yang, F.Y.; Zhang, L.; Huang, W.J.; Hou, W.; Fan, C.F.; Jin, R.H.; Feng, Y.M.; Wang, Y.C.; et al. Cathepsin L Plays a Key Role in SARS-CoV-2 Infection in Humans and Humanized Mice and Is a Promising Target for New Drug Development. Signal Transduct. Target. Ther. 2021, 6, 134. [Google Scholar] [CrossRef]
- Ou, X.; Liu, Y.; Lei, X.; Li, P.; Mi, D.; Ren, L.; Guo, L.; Guo, R.; Chen, T.; Hu, J.; et al. Characterization of Spike Glycoprotein of SARS-CoV-2 on Virus Entry and Its Immune Cross-Reactivity with SARS-CoV. Nat. Commun. 2020, 11, 1620. [Google Scholar] [CrossRef] [Green Version]
- Hood, C.L.; Abraham, J.; Boyington, J.C.; Leung, K.; Kwong, P.D.; Nabel, G.J. Biochemical and Structural Characterization of Cathepsin L-Processed Ebola Virus Glycoprotein: Implications for Viral Entry and Immunogenicity. J. Virol. 2010, 84, 2972–2982. [Google Scholar] [CrossRef] [Green Version]
- Coleman, M.D.; Ha, S.D.; Haeryfar, S.M.M.; Barr, S.D.; Kim, S.O. Cathepsin B Plays a Key Role in Optimal Production of the Influenza a Virus. J. Virol. Antivir. Res. 2018, 7, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Zhou, T.; Tsybovsky, Y.; Gorman, J.; Rapp, M.; Cerutti, G.; Chuang, G.Y.; Katsamba, P.S.; Sampson, J.M.; Schon, A.; Bimela, J.; et al. Cryo-Em Structures of SARS-CoV-2 Spike without and with Ace2 Reveal a Ph-Dependent Switch to Mediate Endosomal Positioning of Receptor-Binding Domains. Cell Host Microbe 2020, 28, 867–879.e5. [Google Scholar] [CrossRef]
- Liu, Q.; Somiya, M.; Kuroda, S. Elucidation of the Early Infection Machinery of Hepatitis B Virus by Using Bio-Nanocapsule. World J. Gastroenterol. 2016, 22, 8489–8496. [Google Scholar] [CrossRef] [PubMed]
- Valdeira, M.L.; Bernardes, C.; Cruz, B.; Geraldes, A. Entry of African Swine Fever Virus into Vero Cells and Uncoating. Vet. Microbiol. 1998, 60, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Nicola, A.V. Herpesvirus Entry into Host Cells Mediated by Endosomal Low Ph. Traffic 2016, 17, 965–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, J.L.; Weed, D.J.; Lee, B.H.; Pritchard, S.M.; Nicola, A.V. Low-Ph Endocytic Entry of the Porcine Alphaherpesvirus Pseudorabies Virus. J. Virol. 2019, 93, e01849-18. [Google Scholar] [CrossRef] [Green Version]
- Komala Sari, T.; Gianopulos, K.A.; Weed, D.J.; Schneider, S.M.; Pritchard, S.M.; Nicola, A.V. Herpes Simplex Virus Glycoprotein C Regulates Low-Ph Entry. mSphere 2020, 5, e00826-19. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Ojha, C.R.; Russell, C.J. Relationship between Hemagglutinin Stability and Influenza Virus Persistence after Exposure to Low Ph or Supraphysiological Heating. PLoS Pathog. 2021, 17, e1009910. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PEDV | Compound | CC50 (μM) | IC50 (μM) | SI (CC50/IC50) |
|---|---|---|---|---|
| HLJBY | BBM | >20 | 9 | >2.22 |
| FAN | 17 | 3.54 | 4.8 | |
| +FAN | 16 | 4.68 | 3.41 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Chen, H.; Sun, L.; Zhao, P.; Qi, C.; Yang, Y.; Si, A.; Qian, Y.; Jung, Y.-S. Bis-Benzylisoquinoline Alkaloids Inhibit Porcine Epidemic Diarrhea Virus by Disrupting Virus Entry. Pathogens 2023, 12, 845. https://doi.org/10.3390/pathogens12060845
Zhang C, Chen H, Sun L, Zhao P, Qi C, Yang Y, Si A, Qian Y, Jung Y-S. Bis-Benzylisoquinoline Alkaloids Inhibit Porcine Epidemic Diarrhea Virus by Disrupting Virus Entry. Pathogens. 2023; 12(6):845. https://doi.org/10.3390/pathogens12060845
Chicago/Turabian StyleZhang, Caisheng, Huan Chen, Liumei Sun, Pu Zhao, Chuanxiang Qi, Ying Yang, Anqi Si, Yingjuan Qian, and Yong-Sam Jung. 2023. "Bis-Benzylisoquinoline Alkaloids Inhibit Porcine Epidemic Diarrhea Virus by Disrupting Virus Entry" Pathogens 12, no. 6: 845. https://doi.org/10.3390/pathogens12060845