

Curcumin and Its Supramolecular Complex with Disodium Glycyrrhizinate as Potential Drugs for the Liver Fluke Infection Caused by Opisthorchis felineus
 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Drugs
2.3. Design of In Vitro Experiments
2.4. Design of In Vivo Experiments
2.5. The Open Field Test
2.6. Biochemical Assays
2.7. RNA Isolation for Real-Time PCR
2.8. Histological and Immunohistochemical Examination of Liver Sections
2.9. Statistical Analysis
3. Results
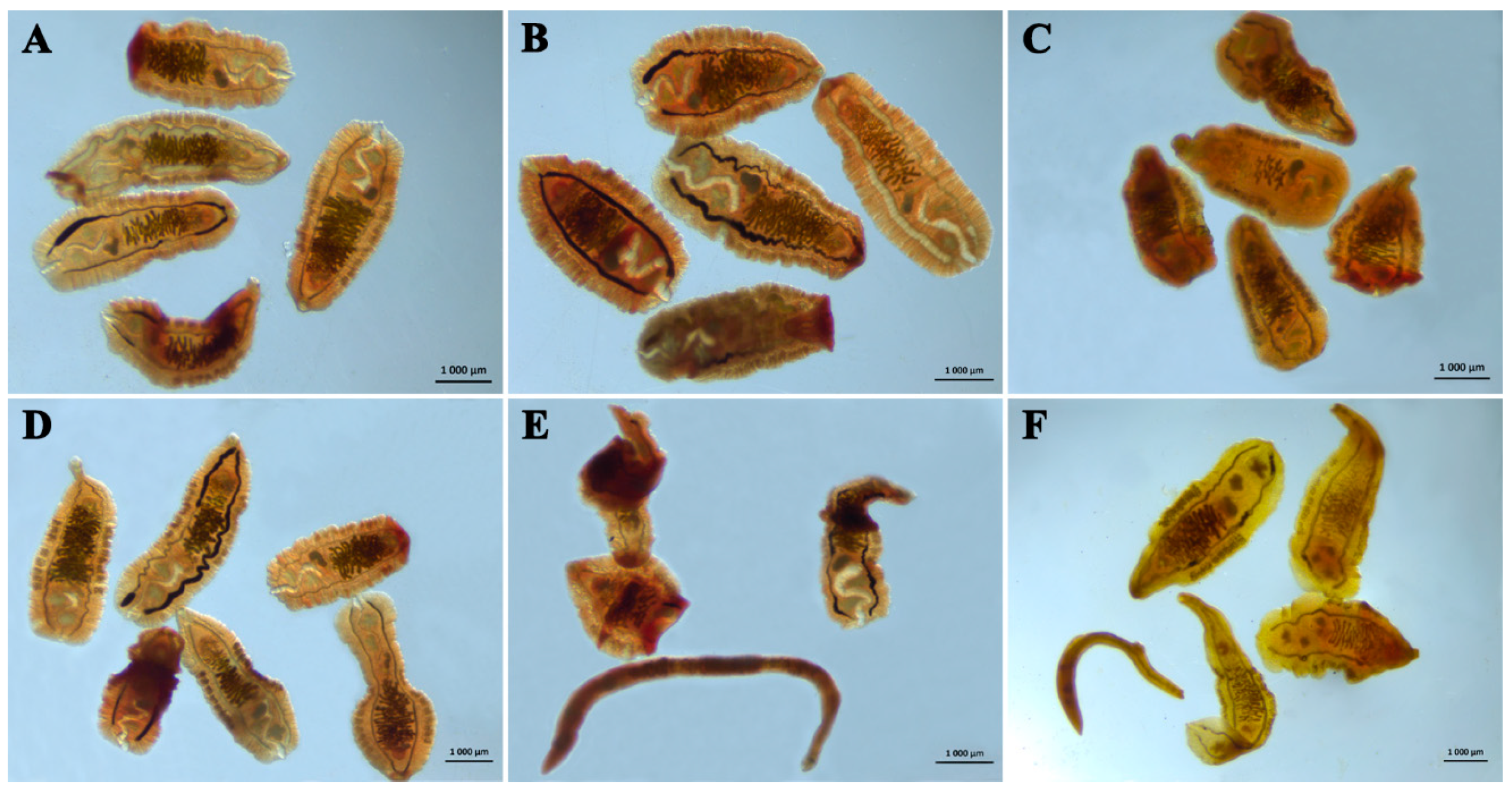
3.1. In Vitro Data
3.2. In Vivo Data
3.2.1. Animals’ Behavior in the Open Field Test
3.2.2. Inspection of the Pancreatic Duct
3.2.3. The Livers of Golden Hamsters
3.2.4. Anthelmintic Effects of the Compounds
3.2.5. Body Weights and Relative Liver and Spleen Weights
3.2.6. Biochemical Results
3.2.7. Gene Expression Analysis in Liver Samples
3.2.8. Histological Data
3.2.9. Immunohistochemical Data
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kassai, T.; Cordero del Campillo, M.; Euzeby, J.; Gaafar, S.; Hiepe, T.; Himonas, C.A. Standardized nomenclature of animal parasitic diseases (SNOAPAD). Vet. Parasitol. 1988, 29, 299–326. [Google Scholar] [CrossRef]
- Mordvinov, V.A.; Yurlova, N.I.; Ogorodova, L.M.; Katokhin, A.V. Opisthorchis felineus and Metorchis bilis are the main agents of liver fluke infection of humans in Russia. Parasitol. Int. 2012, 61, 25–31. [Google Scholar] [CrossRef]
- Sripa, B.; Tangkawattana, S.; Brindley, P.J. Update on pathogenesis of opisthorchiasis and cholangiocarcinoma. Adv Parasitol. 2018, 102, 97–113. [Google Scholar] [CrossRef]
- Sripa, B.; Suwannatrai, A.T.; Sayasone, S.; Do, D.T.; Khieu, V.; Yang, Y. Current status of human liver fluke infections in the Greater Mekong Subregion. Acta Trop. 2021, 224, 106133. [Google Scholar] [CrossRef]
- Traverso, A.; Repetto, E.; Magnani, S.; Meloni, T.; Natrella, M.; Marchisio, P.; Giacomazzi, C.; Bernardi, P.; Gatti, S.; Gomez Morales, M.A.; et al. A large outbreak of Opisthorchis felineus in Italy suggests that opisthorchiasis develops as a febrile eosinophilic syndrome with cholestasis rather than a hepatitis-like syndrome. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 1089–1093. [Google Scholar] [CrossRef]
- Yurlova, N.I.; Yadrenkina, E.N.; Rastyazhenko, N.M.; Serbina, E.A.; Glupov, V.V. Opisthorchiasis in Western Siberia: Epidemiology and distribution in human, fish, snail, and animal populations. Parasitol. Int. 2017, 66, 355–364. [Google Scholar] [CrossRef]
- Schuster, R.; Bonin, J.; Staubach, C.; Heidrich, R. Liver fluke (Opisthorchiidae) findings in red foxes (Vulpes vulpes) in the eastern part of the Federal State Brandenburg, Germany: A contribution to the epidemiology of opisthorchiidosis. Parasitol. Res. 1999, 85, 142–146. [Google Scholar] [CrossRef]
- WHO World Health Organization. Neglected Tropical Diseases: Opisthorchiasis. 28 July 2020. Available online: https://www.who.int/news-room/questions-and-answers/item/neglected-tropical-diseases-opisthorchiasis (accessed on 6 June 2023).
- Sayasone, S.; Keiser, J.; Meister, I.; Vonghachack, Y.; Xayavong, S.; Senggnam, K.; Phongluxa, K.; Hattendorf, J.; Odermatt, P. Efficacy and safety of tribendimidine versus praziquantel against Opisthorchis viverrini in Laos: An open-label, randomised, non-inferiority, phase 2 trial. Lancet Infect. Dis. 2018, 18, 155–161. [Google Scholar] [CrossRef] [Green Version]
- Sereerak, P.; Upontain, S.; Tangkawattana, P.; Mallory, F.F.; Sripa, B.; Tangkawattana, S. Efficacious and safe dose of praziquantel for the successful treatment of feline reservoir hosts with opisthorchiasis. Parasitol. Int. 2017, 66, 448–452. [Google Scholar] [CrossRef]
- Pakharukova, M.Y.; Shilov, A.G.; Pirozhkova, D.S.; Katokhin, A.V.; Mordvinov, V.A. The first comprehensive study of praziquantel effects in vivo and in vitro on European liver fluke Opisthorchis felineus (Trematoda). Int. J. Antimicrob. Agents. 2015, 46, 94–100. [Google Scholar] [CrossRef]
- Avgustinovich, D.; Tsyganov, M.; Vishnivetskaya, G.; Kovner, A.; Sorokina, I.; Orlovskaya, I.; Toporkova, L.; Goiman, E.; Tolstikova, T.; Dushkin, A.; et al. Effects of supramolecular complexation of praziquantel with disodium glycyrrhizinate on the liver fluke Opisthorchis felineus: An in vitro and in vivo study. Acta Tropica. 2019, 194, 1–12. [Google Scholar] [CrossRef]
- Kamsa-Ard, S.; Luvira, V.; Pugkhem, A.; Luvira, V.; Thinkhamrop, B.; Suwanrungruang, K.; Bhudhisawasdi, V. Association between praziquantel treatment and cholangiocarcinoma: A hospital-based matched case-control study. BMC Cancer. 2015, 15, 776. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, B.B.; Sung, B. Pharmacological basis for the role of curcumin in chronic diseases: An age-old spice with modern targets. Trends Pharmacol. Sci. 2009, 30, 85–94. [Google Scholar] [CrossRef]
- Yallapu, M.M.; Nagesh, P.K.; Jaggi, M.; Chauhan, S.C. Therapeutic applications of curcumin nanoformulations. AAPS J. 2015, 17, 1341–1356. [Google Scholar] [CrossRef] [Green Version]
- Cole, G.M.; Teter, B.; Frautschy, S.A. Neuroprotective effects of curcumin. Adv. Exp. Med. Biol. 2007, 595, 197–212. [Google Scholar] [CrossRef] [Green Version]
- Pinlaor, S.; Yongvanit, P.; Prakobwong, S.; Kaewsamut, B.; Khoontawad, J.; Pinlaor, P.; Hiraku, Y. Curcumin reduces oxidative and nitrative DNA damage through balancing of oxidant-antioxidant status in hamsters infected with Opisthorchis viverrini. Mol. Nutr. Food Res. 2009, 53, 1316–1328. [Google Scholar] [CrossRef]
- Charoensuk, L.; Pinlaor, P.; Prakobwong, S.; Hiraku, Y.; Laothong, U.; Ruangjirachuporn, W.; Yongvanit, P.; Pinlaor, S. Curcumin induces a nuclear factor-erythroid 2-related factor 2-driven response against oxidative and nitrative stress after praziquantel treatment in liver fluke-infected hamsters. Int. J. Parasitol. 2011, 41, 615–626. [Google Scholar] [CrossRef]
- Pinlaor, S.; Prakobwong, S.; Hiraku, Y.; Pinlaor, P.; Laothong, U.; Yongvanit, P. Reduction of periductal fibrosis in liver fluke-infected hamsters after long-term curcumin treatment. Eur. J. Pharmacol. 2010, 638, 134–141. [Google Scholar] [CrossRef]
- El-Bahy, N.M.; Bazh, E.K. Anthelmintic activity of ginger, curcumin, and praziquentel against Raillietina cesticillus (in vitro and in vivo). Parasitol. Res. 2015, 114, 2427–2434. [Google Scholar] [CrossRef]
- Nasai, N.B.; Abba, Y.; Abdullah, F.F.; Marimuthu, M.; Tijjani, A.; Sadiq, M.A.; Mohammed, K.; Chung, E.L.; Omar, M.A. In vitro larvicidal effects of ethanolic extract of Curcuma longa Linn. on Haemonchus larval stage. Vet. World. 2016, 9, 417–420. [Google Scholar] [CrossRef] [Green Version]
- Martínez-González, J.J.; Ríos-Morales, S.L.; Guevara-Flores, A.; Ramos-Godinez, M.D.P.; López-Saavedra, A.; Rendón, J.L.; Del Arenal Mena, I.P. Evaluating the effect of curcumin on the metacestode of Taenia crassiceps. Exp. Parasitol. 2022, 239, 108319. [Google Scholar] [CrossRef]
- Anand, P.; Kunnumakkara, A.B.; Newman, R.A.; Aggarwal, B.B. Bioavailability of Curcumin: Problems and Promises. Mol. Pharm. 2007, 4, 807–818. [Google Scholar] [CrossRef]
- Yavarpour-Bali, H.; Ghasemi-Kasman, M.; Pirzadeh, M. Curcumin-loaded nanoparticles: A novel therapeutic strategy in treatment of central nervous system disorders. Int. J. Nanomed. 2019, 14, 4449–4460. [Google Scholar] [CrossRef] [Green Version]
- Jantawong, C.; Priprem, A.; Intuyod, K.; Pairojkul, C.; Pinlaor, P.; Waraasawapati, S.; Mongkon, I.; Chamgramol, Y.; Pinlaor, S. Curcumin-loaded nanocomplexes: Acute and chronic toxicity studies in mice and hamsters. Toxicol. Rep. 2021, 8, 1346–1357. [Google Scholar] [CrossRef]
- Charoensuk, L.; Pinlaor, P.; Wanichwecharungruang, S.; Intuyod, K.; Vaeteewoottacharn, K.; Chaidee, A.; Yongvanit, P.; Pairojkul, C.; Suwannateep, N.; Pinlaor, S. Nanoencapsulated curcumin and praziquantel treatment reduces periductal fibrosis and attenuates bile canalicular abnormalities in Opisthorchis viverrini-infected hamsters. Nanomedicine 2016, 12, 21–32. [Google Scholar] [CrossRef]
- Kong, R.; Zhu, X.; Meteleva, E.S.; Chistyachenko, Y.S.; Suntsova, L.P.; Polyakov, N.E.; Khvostov, M.V.; Baev, D.S.; Tolstikova, T.G.; Yu, J.; et al. Enhanced solubility and bioavailability of simvastatin by mechanochemically obtained complexes. Int. J. Pharm. 2017, 534, 108–118. [Google Scholar] [CrossRef]
- Meteleva, E.S.; Chistyachenko, Y.S.; Suntsova, L.P.; Tsyganov, M.A.; Vishnivetskaya, G.B.; Avgustinovich, D.F.; Khvostov, M.V.; Polyakov, N.E.; Tolstikova, T.G.; Mordvinov, V.A.; et al. Physicochemical properties and anti-opisthorchosis effect of mechanochemically synthesized solid compositions of praziquantel with glycyrrhizic acid disodium salt. Dokl. Biochem. Biophys. 2018, 481, 228–231. [Google Scholar] [CrossRef]
- Meteleva, E.S.; Chistyachenko, Y.S.; Suntsova, L.P.; Khvostov, M.V.; Polyakov, N.E.; Selyutina, O.Y.; Tolstikova, T.G.; Frolova, T.S.; Mordvinov, V.A.; Dushkin, A.V.; et al. Disodium salt of glycyrrhizic acid—A novel supramolecular delivery system for anthelmintic drug praziquantel. J. Drug Deliv. Sci. Technol. 2019, 50, 66–77. [Google Scholar] [CrossRef]
- Selyutina, O.Y.; Polyakov, N.E. Glycyrrhizic acid as a multifunctional drug carrier—From physicochemical properties to biomedical applications: A modern insight on the ancient drug. Int. J. Pharm. 2019, 559, 271–279. [Google Scholar] [CrossRef]
- Zhang, Q.; Polyakov, N.E.; Chistyachenko, Y.S.; Khvostov, M.V.; Frolova, T.S.; Tolstikova, T.G.; Dushkin, A.V.; Su, W. Preparation of curcumin self-micelle solid dispersion with enhanced bioavailability and cytotoxic activity by mechanochemistry. Drug Deliv. 2018, 25, 198–209. [Google Scholar] [CrossRef]
- Keiser, J.; Manneck, T.; Vargas, M. Interactions of mefloquine with praziquantel in the Schistosomam mansoni mouse model and in vitro. J. Antimicrob. Chemother. 2011, 66, 1791–1797. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.C.; Martin, N. Compu Syn for Drug Combinations: PC Software and User’s Guide: A Computer Program for Quantitation of Synergism and Antagonismin Drug Combinations, and the Determination of IC50 and ED50 and LD50 Values; ComboSyn Inc: Paramus, NJ, USA, 2005. [Google Scholar]
- Avgustinovich, D.F.; Orlovskaya, I.A.; Toporkova, L.B.; Vishnivetskaya, G.B.; Katokhin, A.V.; Lvova, M.N.; Kashina, E.V.; Bondar, N.P.; Feofanova, N.A.; Mordvinov, V.A. Experimental opisthorchiasis: Study of blood cell composition, hematopoiesis, and startle reflex in laboratory animals. Russ. J. Genet. Appl. Res. 2017, 7, 82–92. [Google Scholar] [CrossRef]
- Wonkchalee, N.; Boonmars, T.; Laummaunwai, P.; Aromdee, C.; Hahnvajanawong, C.; Wu, Z.; Sriraj, P.; Aukkanimart, R.; Chamgramol, Y.; Pairojkul, C.; et al. A combination of praziquantel and the traditional medicinal plant Thunbergia laurifolia on Opisthorchis viverrini infection and cholangiocarcinoma in a hamster model. Parasitol. Res. 2013, 112, 4211–4219. [Google Scholar] [CrossRef] [PubMed]
- Prommajun, P.; Phetcharaburanin, J.; Namwat, N.; Klanrit, P.; Sa-Ngiamwibool, P.; Thanee, M.; Dokduang, H.; Kittirat, Y.; Li, J.V.; Loilome, W. Metabolic Profiling of Praziquantel-mediated Prevention of Opisthorchis viverrini-induced Cholangiocyte Transformation in the Hamster Model of Cholangiocarcinoma. Cancer Genom. Proteom. 2021, 18, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Sotelo, J.; Jung, H. Pharmacokinetic optimisation of the treatment of neurocysticercosis. Clin. Pharm. 1998, 34, 503–515. [Google Scholar] [CrossRef]
- Maiti, P.; Dunbar, G.L. Use of curcumin, a natural polyphenol for targeting molecular pathways in treating age-related neurodegenerative diseases. Int. J. Mol. Sci. 2018, 19, 1637. [Google Scholar] [CrossRef] [Green Version]
- Mandour, M.E.M.; Turabi, H.E.; Homeida, M.M.A.; el Sadig, T.; Ali, H.M.; Bennett, J.L.; Leahey, W.J.; Harron, D.W. Pharmacokinetics of praziquantel in healthy volunteers and patients with schistosomiasis. Trans. R. Soc. Trop. Med. Hyg. 1990, 84, 389–393. [Google Scholar] [CrossRef]
- Shoba, G.; Joy, D.; Joseph, T.; Majeed, M.; Rajendran, R.; Srinivas, P.S. Influence of piperine on the pharmacokinetics of curcumin in animals and human volunteers. Planta Med. 1998, 64, 353–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, R.A.; Steward, W.P.; Gescher, A.J. Pharmacokinetics and pharmacodynamics of curcumin. In The Molecular Targets and Therapeutic Uses of Curcumin in Health and Disease; Aggarwal, B.B., Surh, Y.-J., Shishodia, S., Eds.; Springer Science+Business Media, LLC: New York, NY, USA, 2007; pp. 453–470. Available online: https://link.springer.com/chapter/10.1007/978-0-387-46401-5_20 (accessed on 6 June 2023).
- Friard, O.; Gamba, M. BORIS: A free, versatile open-source event-logging software for video/audio coding and live observations. Meth. Ecol. Evol. 2016. 7, 1325–1330. [CrossRef]
- Pakharukova, M.Y.; Zaparina, O.; Baginskaya, N.V.; Mordvinov, V.A. Global changes in gene expression related to Opisthorchis felineus liver fluke infection reveal temporal heterogeneity of a mammalian host response. Food Waterborne Parasitol. 2022, 27, e00159. [Google Scholar] [CrossRef]
- Pakharukova, M.Y.; Zaparina, O.G.; Kovner, A.V.; Mordvinov, V.A. Inhibition of Opisthorchis felineus glutathione-dependent prostaglandin synthase by resveratrol correlates with attenuation of cholangiocyte neoplasia in a hamster model of opisthorchiasis. Int. J. Parasitol. 2019, 49, 963–973. [Google Scholar] [CrossRef]
- Mistlberger, R.E.; Antle, M.C.; Webb, I.C.; Jones, M.; Weinberg, J.; Pollock, M.S. Circadian clock resetting by arousal in Syrian hamsters: The role of stress and activity. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003, 285, R917–R925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Wang, H.; Jiao, R.; Peng, C.; Wong, Y.M.; Yeung, V.S.Y.; Huang, Y.; Chen, Z.-Y. Choosing hamsters but not rats as a model for studying plasma cholesterol-lowering activity of functional foods. Mol. Nutr. Food. Res. 2009, 53, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Stofan, M.; Guo, G.L. Bile acids and FXR: Novel targets for liver diseases. Front. Med. 2020, 7, 544. [Google Scholar] [CrossRef] [PubMed]
- Repa, J.J.; Turley, S.D.; Lobaccaro, J.A.; Medina, J.; Li, L.; Lustig, K.; Shan, B.; Heyman, R.A.; Dietschy, J.M.; Mangelsdorf, D.J. Regulation of absorption and ABC1-mediated efflux of cholesterol by RXR heterodimers. Science 2000, 289, 1524–15299. [Google Scholar] [CrossRef] [PubMed]
- Mordvinov, V.A.; Furman, D.P. The Digenea parasite Opisthorchis felineus: A target for the discovery and development of novel drugs. Infect. Disord. Drug. Targets 2010, 10, 385–401. [Google Scholar] [CrossRef]
- Brazhnikova, N.A.; Tskhai, V.F. Clinical features, diagnisis and management of the complicated opistorkhosis. Ann. Surg. Pathol. 2004, 9, 40–44. Available online: https://hepatoassociation.ru/ASH/Volumes/Pdf92/Brag92.pdf (accessed on 1 September 2022). (In Russian).
- Feng, D.; Zou, J.; Zhang, S.; Li, X.; Lu, M. Hypocholesterolemic activity of curcumin is mediated by down-regulating the expression of niemann-pick c1-like 1 in hamsters. J. Agric. Food Chem. 2017, 65, 276–280. [Google Scholar] [CrossRef]
- Hong, T.; Zou, J.; Jiang, X.; Yang, J.; Cao, Z.; He, Y.; Feng, D. Curcumin supplementation ameliorates bile cholesterol supersaturation in hamsters by modulating gut microbiota and cholesterol absorption. Nutrients 2022, 14, 1828. [Google Scholar] [CrossRef]
- Lee, F.Y.; Lee, H.; Hubbert, M.L.; Edwards, P.A.; Zhang, Y. FXR, a multipurpose nuclear receptor. Trends Biochem. Sci. 2006, 31, 572–580. [Google Scholar] [CrossRef]
- Kast, H.R.; Nguyen, C.M.; Sinal, C.J.; Jones, S.A.; Laffitte, B.A.; Reue, K.; Gonzalez, F.J.; Willson, T.M.; Edwards, P.A. Farnesoid X-activated receptor induces apolipoprotein C-II transcription: A molecular mechanism linking plasma triglyceride levels to bile acids. Mol. Endocrinol. 2001, 15, 1720–1728. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.Y.; He, H.; Nguyen, T.; Mennone, A.; Boyer, J.L. Retinoic acid represses CYP7A1 expression in human hepatocytes and HepG2 cells by FXR/RXR-dependent and independent mechanisms. J. Lipid Res. 2010, 51, 2265–2274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, J.Y.L. Bile acid metabolism and signaling. Compr. Physiol. 2013, 3, 1191–1212. [Google Scholar] [CrossRef] [Green Version]
- Charoensuk, L.; Pinlaor, P.; Laothong, U.; Yongvanit, P.; Pairojkul, C.; Nawa, Y.; Pinlaor, S. Bile canalicular changes and defective bile secretion in Opisthorchis viverrini-infected hamsters. Folia Parasitol. 2014, 61, 512–522. [Google Scholar] [CrossRef] [Green Version]
- Toydemir, T.; Kanter, M.; Erboga, M.; Oguz, S.; Erenoglu, C. Antioxidative, antiapoptotic, and proliferative effect of curcumin on liver regeneration after partial hepatectomy in rats. Toxicol. Ind. Health 2015, 31, 162–172. [Google Scholar] [CrossRef] [PubMed]
- Chuang, S.E.; Cheng, A.L.; Lin, J.K.; Kuo, M.L. Inhibition by curcumin of diethylnitrosamine-induced hepatic hyperplasia, inflammation, cellular gene products and cell-cycle-related proteins in rats. Food Chem. Toxicol. 2000, 38, 991–995. [Google Scholar] [CrossRef]
- Liu, J.J.; Raynal, S.; Bailbé, D.; Gausseres, B.; Carbonne, C.; Autier, V.; Movassat, J.; Kergoat, M.; Portha, B. Expression of the kynurenine pathway enzymes in the pancreatic islet cells. Activation by cytokines and glucolipotoxicity. Biochim. Biophys. Acta. 2015, 1852, 980–991. [Google Scholar] [CrossRef] [Green Version]
- Garrison, A.M.; Parrott, J.M.; Tuñon, A.; Delgado, J.; Redus, L.; O’Connor, J.C. Kynurenine pathway metabolic balance influences microglia activity: Targeting kynurenine monooxygenase to dampen neuroinflammation. Psychoneuroendocrinology 2018, 94, 1–10. [Google Scholar] [CrossRef]
- Huang, Y.S.; Ogbechi, J.; Clanchy, F.I.; Williams, R.O.; Stone, T.W. IDO and kynurenine metabolites in peripheral and CNS disorders. Front. Immunol. 2020, 11, 388. [Google Scholar] [CrossRef] [Green Version]
- Jin, H.; Zhang, Y.; You, H.; Tao, X.; Wang, C.; Jin, G.; Wang, N.; Ruan, H.; Gu, D.; Huo, X.; et al. Prognostic significance of kynurenine 3-monooxygenase and effects on proliferation, migration, and invasion of human hepatocellular carcinoma. Sci. Rep. 2015, 5, 10466. [Google Scholar] [CrossRef] [Green Version]
- Karbysheva, N.; Nikonorova, M.; Matros, O.; Kiushkina, I.; Nemilostiva, E.; Choroshilova, I.; Gorobchenko, A.; Umbetova, K.; Volchkova, E. Clinical polymorphism in patients with Opisthorchis felineus infection in the Western Siberia. IDCases 2021, 24, e01064. [Google Scholar] [CrossRef] [PubMed]
- Pungpak, S.; Riganti, M.; Bunnag, D.; Harinasuta, T. Clinical features in severe opisthorchiasis viverrini. Southeast Asian J. Trop. Med. Public Health 1985, 16, 405–409. [Google Scholar] [PubMed]
- Gupta, S.; Stravitz, R.T.; Dent, P.; Hylemon, P.B. Down-regulation of cholesterol 7alpha-hydroxylase (CYP7A1) gene expression by bile acids in primary rat hepatocytes is mediated by the c-Jun N-terminal kinase pathway. J. Biol. Chem. 2001, 276, 15816–15822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feingold, K.R.; Spady, D.K.; Pollock, A.S.; Moser, A.H.; Grunfeld, C. Endotoxin, TNF, and IL-1 decrease cholesterol 7 alpha-hydroxylase mRNA levels and activity. J. Lipid Res. 1996, 37, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Pakharukova, M.Y.; Zaparina, O.G.; Kapushchak, Y.K.; Baginskaya, N.V.; Mordvinov, V.A. Opisthorchis felineus infection provokes time-dependent accumulation of oxidative hepatobiliary lesions in the injured hamster liver. PLoS ONE 2019, 14, e0216757. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NEMs * | Adult Flukes | |
|---|---|---|
| Na2GA | 30,400 μg/mL | 1236.470 μg/mL |
| PZQ | 0.587 μg/mL | 0.208 μg/mL |
| Cur | 0.611 μg/mL | 0.500 μg/mL |
| Cur:Na2GA (1:1) | 31.720 μg/mL | 3.007 μg/mL |
| Trials | Behavioral Parameters | Experimental Groups | ||||
|---|---|---|---|---|---|---|
| CON | OF | OF + PZQ | OF + Cur | OF + Cur:Na2GA | ||
| 1 | Orienting reaction time, s | 7.2 ± 3.32 | 7.1 ± 1.81 | 9.2 ± 1.52 | 11.6 ± 4.15 | 10.6 ± 4.34 |
| 2 | Orienting reaction time, s | 12.4 ± 2.62 | 11.1 ± 1.81 | 13.7 ± 3.24 | 12.2 ± 4.21 | 9.9 ± 3.95 |
| 1 | Number of crossed squares | 45.2 ± 10.13 | 69.7 ± 10.18 | 45.0 ± 8.08 | 64.0 ± 12.34 | 78.3 ± 11.18 (*) + |
| 2 | Number of crossed squares | 47.7 ± 6.70 | 45.8 ± 7.84 (a) | 24.2 ± 5.07 * (#) (a) | 43.1 ± 10.82 | 40.8 ± 5.80 + a |
| 1 | Number of rearings | 3.4 ± 1.08 | 8.8 ± 1.91 * | 8.4 ± 1.70 (*) | 12.1 ± 2.70 * | 11.2 ± 2.46 * |
| 2 | Number of rearings | 5.3 ± 2.29 | 7.7 ± 2.45 | 1.7 ± 0.42 # aa | 7.9 ± 1.91 + | 5.1 ± 2.58 (a) |
| 1 | Rearing duration, s | 9.6 ± 2.69 | 16.2 ± 4.11 | 18.8 ± 4.76 | 26.1 ± 4.76 * | 21.7 ± 4.42 * |
| 2 | Rearing duration, s | 14.9 ± 6.38 | 13.6 ± 4.07 | 4.6 ± 1.65 a | 19.8 ± 3.7 + | 13.3 ± 7.12 |
| 1 | Number of corner diggings | 0.7 ± 0.49 | 2.3 ± 1.02 | 3.6 ± 1.03 (*) | 3.4 ± 1.18 (*) | 0.9 ± 0.42 (+) |
| 2 | Number of corner diggings | 1.1 ± 0.66 | 3.2 ± 1.44 | 5.5 ± 1.79 * | 4.4 ± 1.42 (*) | 2.3 ± 0.84 |
| 1 | Corner digging duration, s | 1.2 ± 1.01 | 12.0 ± 9.10 | 23.7 ± 9.39 (*) | 17.8 ± 7.42 (*) | 4.4 ± 2.71 |
| 2 | Corner digging duration, s | 2.6 ± 1.77 | 16.2 ± 8.50 | 48.9 ± 18.39 * (#) | 17.8 ± 5.29 * | 19.1 ± 8.77 |
| 1 | Time near central empty Falcon® tube, s | 2.3 ± 0.59 | 1.7 ± 0.36 | 2.3 ± 1.10 | 0.7 ± 0.39 (*) # (+) | 1.7 ± 0.48 |
| 2 | Time near central empty Falcon® tube, s | 2.9 ± 0.75 | 2.5 ± 0.43 | 2.9 ± 0.66 | 1.9 ± 0.89 | 4.6 ± 1.57 |
| 1 | Time near left empty Falcon® tube, s | 3.5 ± 1.02 | 3.0 ± 0.95 | 3.8 ± 1.19 | 1.9 ± 0.50 | 3.2 ± 0.46 (c) |
| 2 | Time near Falcon® tube containing food pellets, s | 14.9 ± 2.01 aa | 22.6 ± 5.06 aa | 16.9 ± 4.84 a | 20.8 ± 6.16 (a) | 28.2 ± 4.32 * aaa |
| 1 | Time near right empty Falcon® tube, s | 4.7 ± 1.92 | 6.0 ± 1.92 | 6.5 ± 2.90 | 2.7 ± 0.89 (#) | 4.8 ± 1.20 |
| 2 | Time near Falcon® tube with valerian, s | 3.4 ± 1.05 | 4.1 ± 2.24 (a) | 8.0 ± 3.32 | 6.1 ± 2.03 | 10.7 ± 3.26 |
| Parameters | Normal Range Ref. [25] | CON | OF | OF + PZQ | OF + Cur | OF + Cur:Na2GA | ANOVA |
|---|---|---|---|---|---|---|---|
| ALT, U/L | 22–128 | 57.3 ± 5.53 | 242.0 ± 39.58 *** | 179.3 ± 50.4 * | 263.7 ± 41.96 *** | 199.3 ± 26.7 ** | F(4,29) = 4.62 p = 0.005 |
| AST, U/L | 20–150 | 107.0 ± 23.35 | 109.4 ± 10.40 | 117.0 ± 13.52 | 136.3 ± 10.24 | 150.0 ± 12.93 * # | F(4,29) = 1.79 p = 0.158 |
| GLU, mmol/L | 3.3–8.3 | 4.2 ± 0.18 | 4.8 ± 0.65 | 5.3 ± 0.22 (*) | 5.0 ± 0.41 | 4.5 ± 0.37 | F(4,29) = 1.03 p = 0.407 |
| TP, g/L | 45–47 | 88.3 ± 3.45 | 80.7 ± 3.19 (*) | 79.9 ± 3.21 (*) | 74.4 ± 2.01 ** | 81.4 ± 3.26 | F(4,29) = 2.46 p = 0.067 |
| CHOL, mmol/L | 1.4–4.7 | 4.0 ± 0.23 | 5.2 ± 0.31 * | 3.7 ± 0.25 ## | 3.9 ± 0.52 ## | 4.8 ± 0.27 + (c) | F(4,29) = 3.54 p = 0.018 |
| TG, mmol/L | 0.81–2.56 | 1.9 ± 0.17 | 2.6 ± 0.18 * | 2.0 ± 0.36 (#) | 1.8 ± 0.14 # | 2.3 ± 0.14 | F(4,29) = 2.18 p = 0.096 |
| Parameters | CON | OF | OF + PZQ | OF + Cur | OF + Cur:Na2GA | ANOVA |
|---|---|---|---|---|---|---|
| Proliferation small bile duct, % | 0.93 ± 0.31 | 17.17 ± 4.59 * | 15.57 ± 5.12 (*) | 22.44 ± 8.44 ** | 17.65 ± 2.28 * | F(4,21) = 2.60 p = 0.065 |
| Cellular infiltrations, % | 0.08 ± 0.06 | 6.11 ± 1.05 (*) | 8.40 ± 4.28 * | 4.00 ± 0.99 | 5.95 ± 1.92 (*) | F(4,20) = 1.73 p = 0.183 |
| Cholangiofibrosis, % | 0.00 ± 0.00 | 16.80 ± 7.61 (*) | 22.77 ± 9.39 * | 21.47 ± 6.78 * | 21.27 ± 7.35 * | F(4,21) = 1.71 p = 0.185 |
| Periductal fibrosis, % | 0.00 ± 0.00 | 48.41 ± 2.28 *** | 24.47 ± 3.27 *** ### | 34.59 ± 7.21 *** # | 23.68 ± 6.35 ** ### | F(4,21) = 16.06 p < 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lvova, M.N.; Ponomarev, D.V.; Tarasenko, A.A.; Kovner, A.V.; Minkova, G.A.; Tsyganov, M.A.; Li, M.; Lou, Y.; Evseenko, V.I.; Dushkin, A.V.; et al. Curcumin and Its Supramolecular Complex with Disodium Glycyrrhizinate as Potential Drugs for the Liver Fluke Infection Caused by Opisthorchis felineus. Pathogens 2023, 12, 819. https://doi.org/10.3390/pathogens12060819
Lvova MN, Ponomarev DV, Tarasenko AA, Kovner AV, Minkova GA, Tsyganov MA, Li M, Lou Y, Evseenko VI, Dushkin AV, et al. Curcumin and Its Supramolecular Complex with Disodium Glycyrrhizinate as Potential Drugs for the Liver Fluke Infection Caused by Opisthorchis felineus. Pathogens. 2023; 12(6):819. https://doi.org/10.3390/pathogens12060819
Chicago/Turabian StyleLvova, Maria N., Denis V. Ponomarev, Alena A. Tarasenko, Anna V. Kovner, Galina A. Minkova, Michail A. Tsyganov, Meijun Li, Yan Lou, Veronica I. Evseenko, Alexander V. Dushkin, and et al. 2023. "Curcumin and Its Supramolecular Complex with Disodium Glycyrrhizinate as Potential Drugs for the Liver Fluke Infection Caused by Opisthorchis felineus" Pathogens 12, no. 6: 819. https://doi.org/10.3390/pathogens12060819