
Low Prevalence of Plasmodium falciparum Histidine-Rich Protein 2 and 3 Gene Deletions—A Multiregional Study in Central and West Africa

 , , , , ,
, , , , ,
Abstract
:1. Introduction
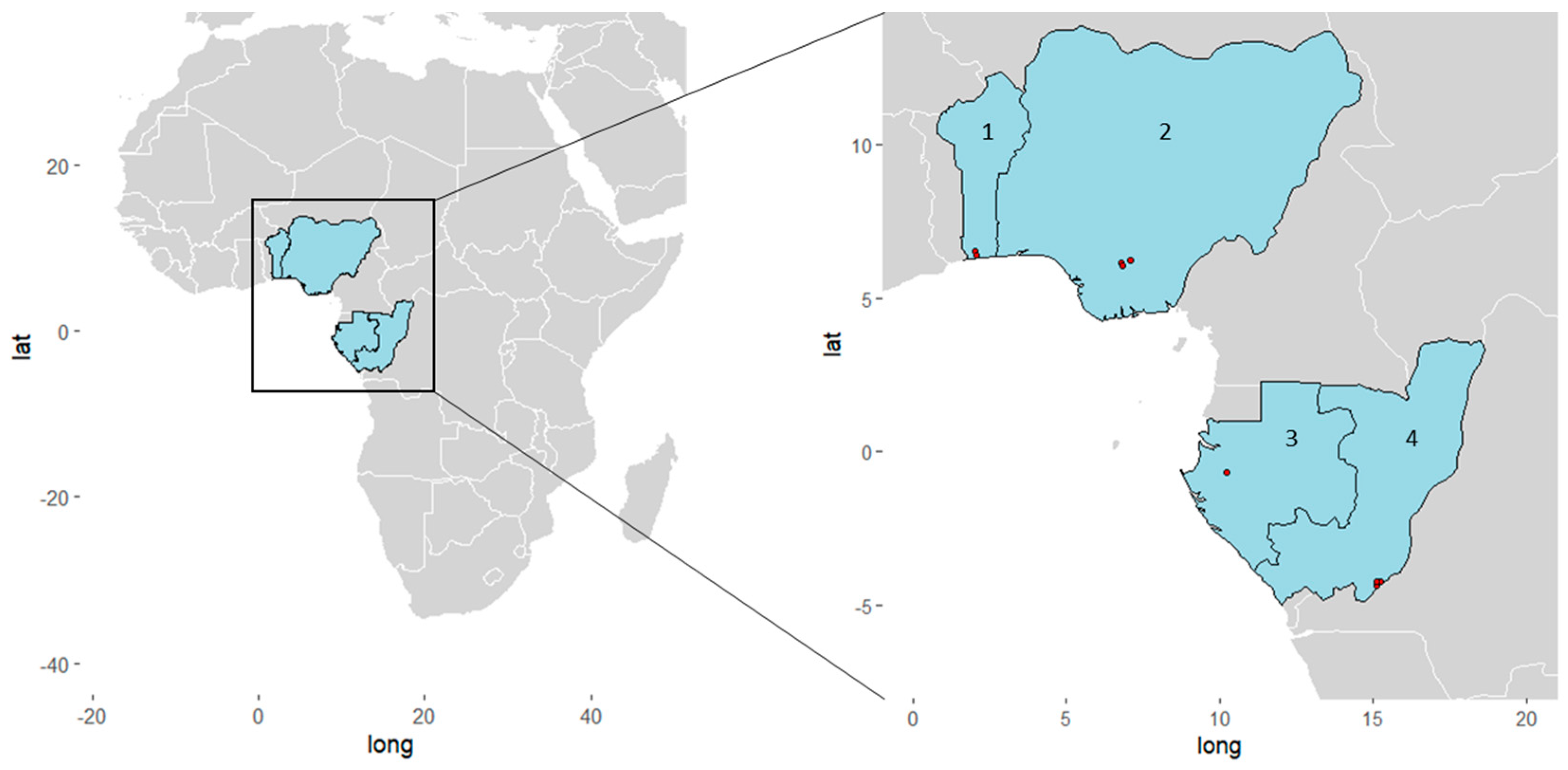
2. Materials and Methods
3. Results
3.1. Baseline Characteristics
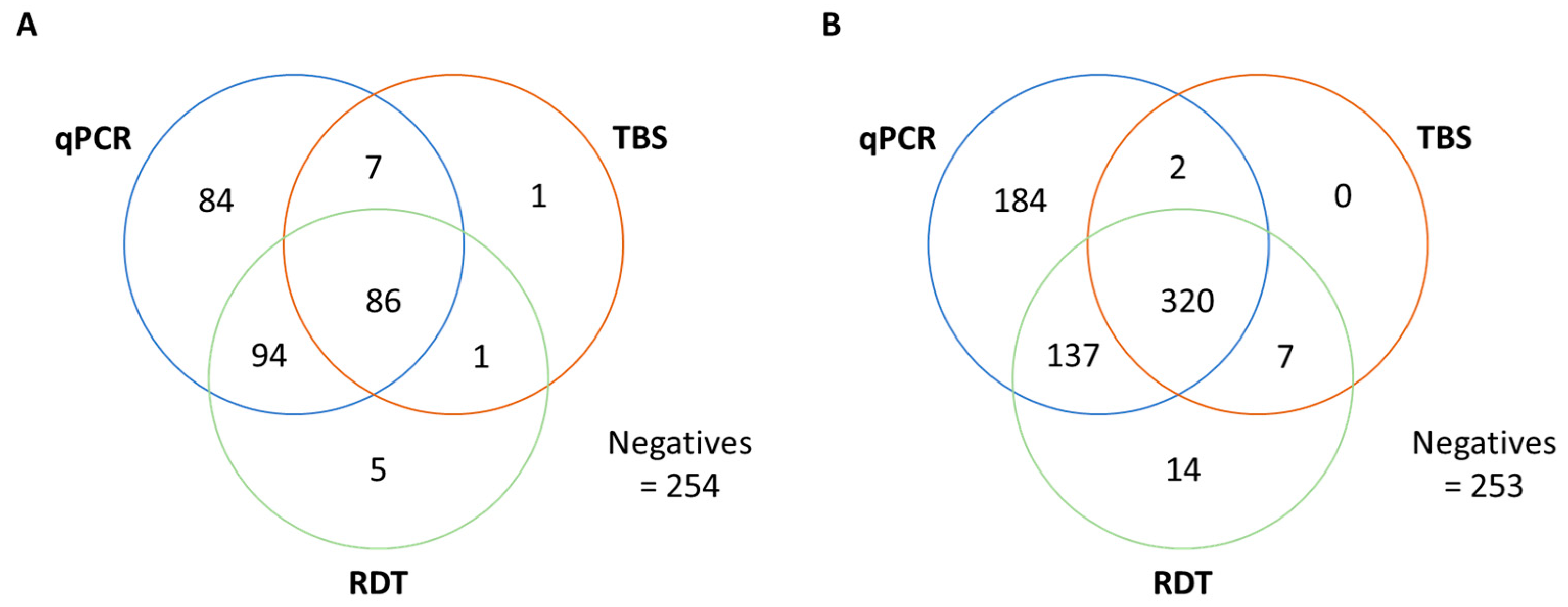
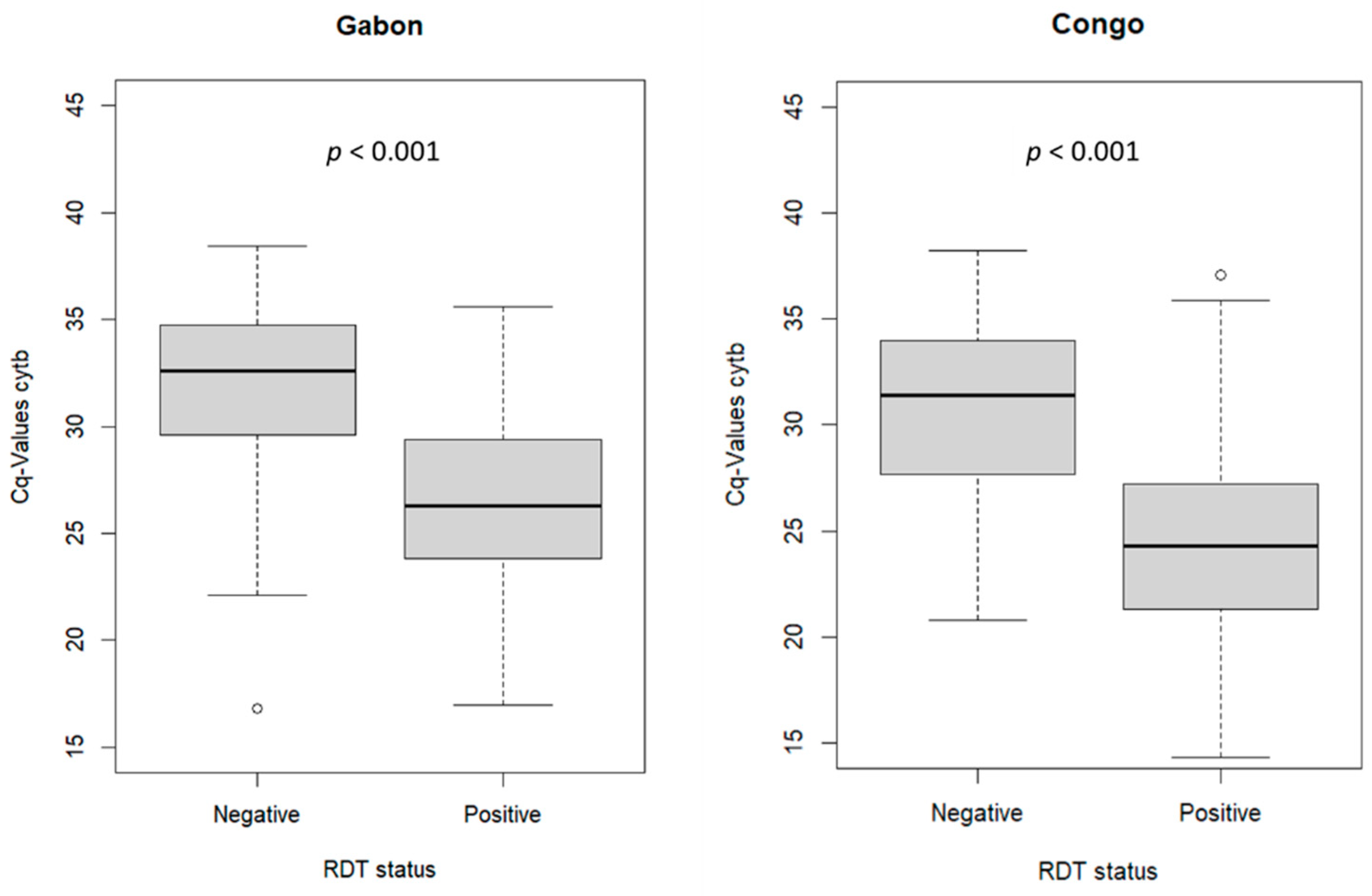
3.2. Malaria Diagnosis Outcome
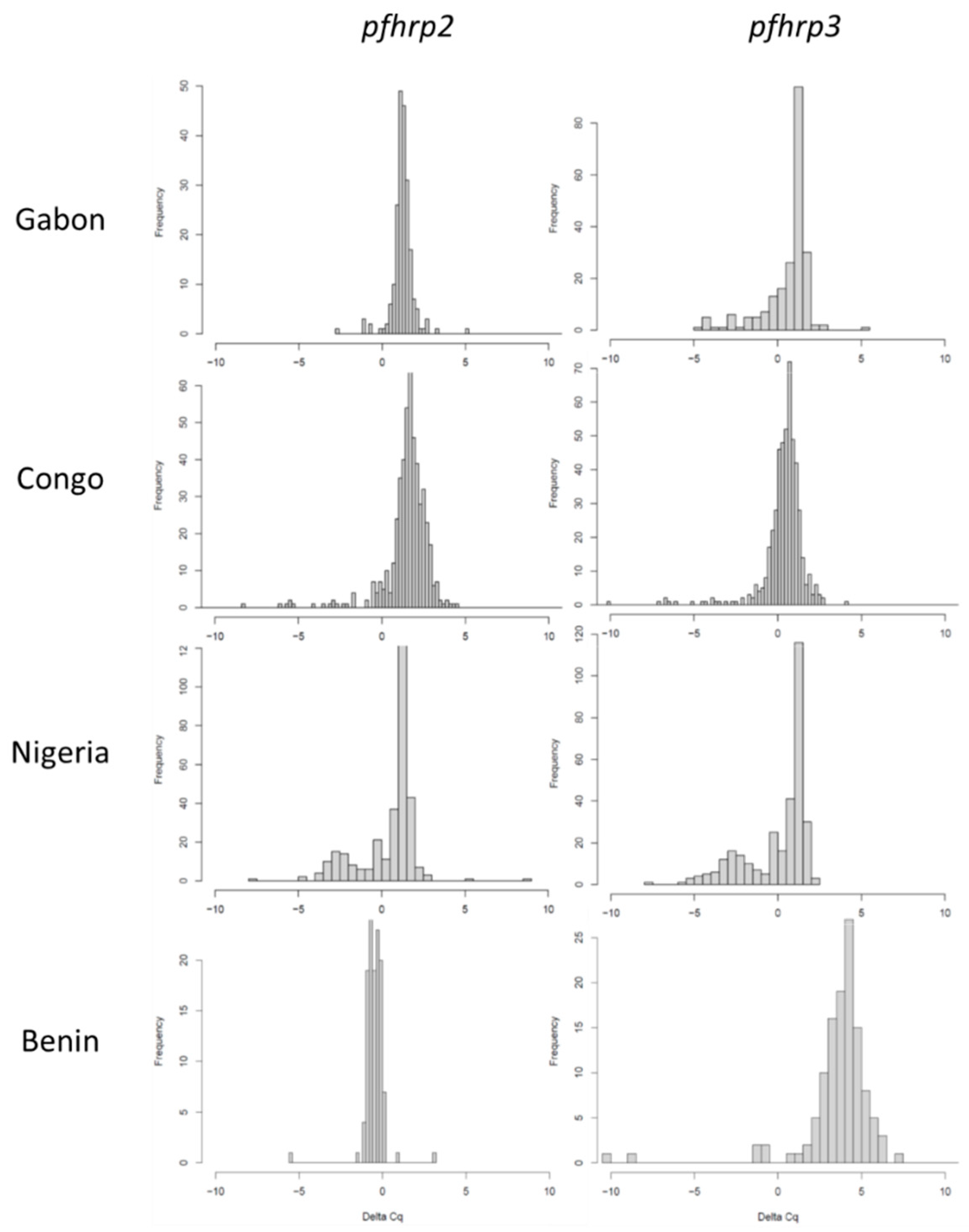
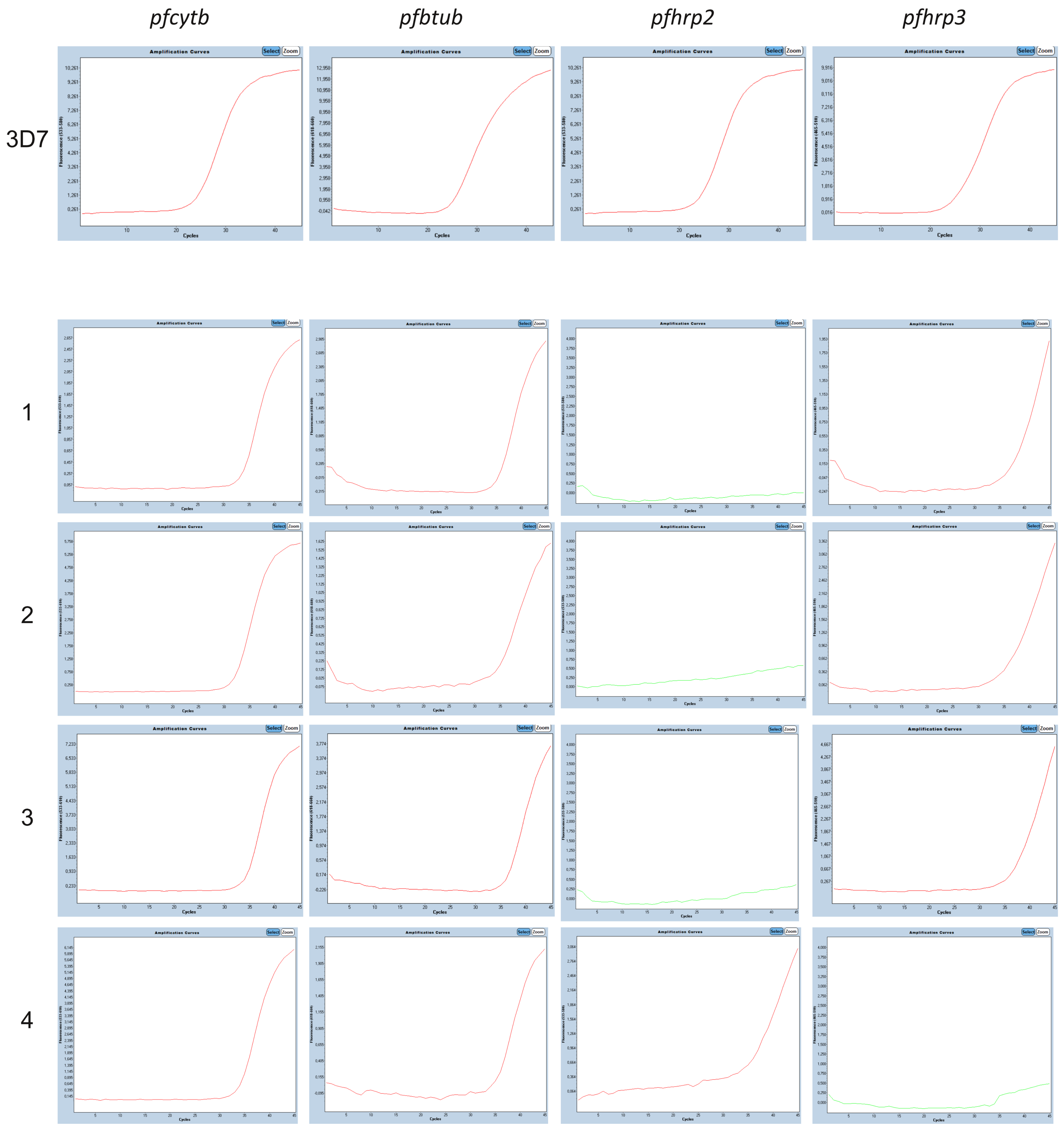
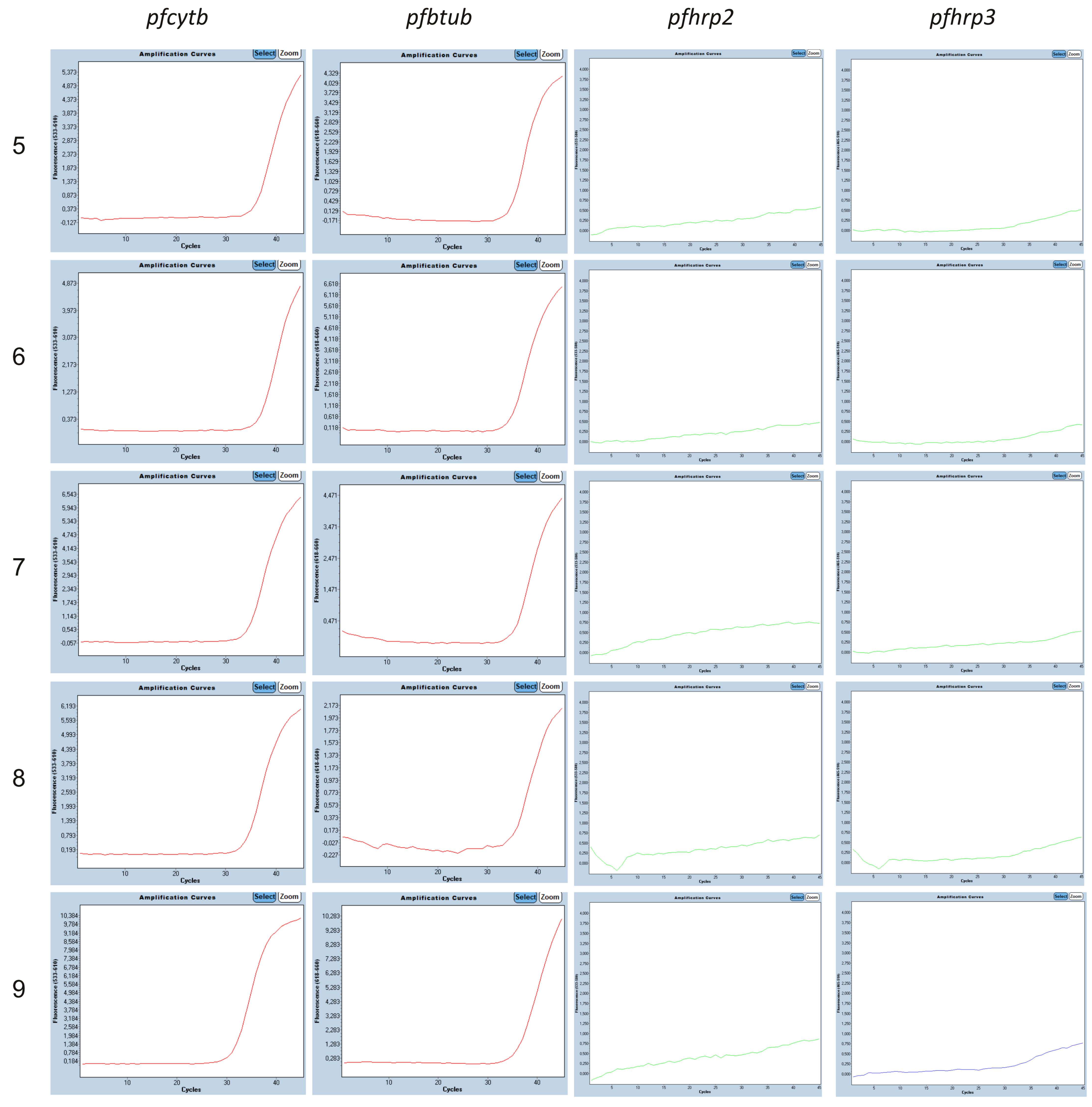
3.3. Molecular pfhrp2 and pfhrp3 Deletion Detection
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Oligonucleotide | Sequence 5′-3′ | Tm [°C] | Expected Amplicon Size [bp] |
|---|---|---|---|---|
| pfcytb | Primer forward | TAC TAA CTT GTT ATC CTC TAT TCC AGT AGC | 55.5 | 240 |
| Primer reverse | CCT TTA ACA TCA AGA CTT AAT AGA TTT GGA | 53.7 | ||
| Probe | [ROX] G+TGC+TAC+CAT+GTA+AAT+GTAA [BHQ2] | 56.0 | ||
| pfßtub | Primer forward | TGA TGT GCG CAA GTG ATC C | 55.8 | 79 |
| Primer reverse | TCC TTT GTG GAC ATT CTT CCT C | 54.6 | ||
| Probe | [Cy5] TA GCA CAT G[TAO]C CGT TAA ATA TCT TCC ATG TCT [IAbRQSp] | 59.6 | ||
| pfhrp2 | Primer forward | TTC CGC ATT TAA TAA TAA CTT GTG | 50.0 | 173 |
| Primer reverse | CGG CTA CAT GAT GAG CAT G | 53.5 | ||
| Probe | [HEX] TAC ACG AAA CTC AAG CAC A [MGBEc] | 51.7 | ||
| pfhrp3 | Primer forward | CTC CGA ATT TAA CAA TAA CTT GTT TA | 50.3 | 200 |
| Primer reverse | CAG CTA CAT GAT GTG CAT G | 51.4 | ||
| Probe | [6-FAM] GAA AGT CAA GCA CAT GCA G [MGBEc] | 52.0 |




References
- World Health Organization. World Malaria Report 2022; World Health Organization: Geneva, Switzerland, 2022. [Google Scholar]
- Lee, N.; Baker, J.; Andrews, K.T.; Gatton, M.L.; Bell, D.; Cheng, Q.; McCarthy, J. Effect of Sequence Variation in Plasmodium falciparum Histidine-Rich Protein 2 on Binding of Specific Monoclonal Antibodies: Implications for Rapid Diagnostic Tests for Malaria. J. Clin. Microbiol. 2006, 44, 2773–2778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amoah, L.E.; Abankwa, J.; Oppong, A. Plasmodium falciparum Histidine Rich Protein-2 Diversity and the Implications for pfhrp 2: Based Malaria Rapid Diagnostic Tests in Ghana. Malar. J. 2016, 15, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huy, N.T.; Serada, S.; Trang, D.T.X.; Takano, R.; Kondo, Y.; Kanaori, K.; Tajima, K.; Hara, S.; Kamei, K. Neutralization of Toxic Heme by Plasmodium Falciparum Histidine-Rich Protein 2. J. Biochem. 2003, 133, 693–698. [Google Scholar] [CrossRef]
- Bosshart, H.; Heinzelmann, M. Endotoxin-Neutralizing Effects of Histidine-Rich Peptides. FEBS Lett. 2003, 553, 135–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poti, K.E.; Sullivan, D.J.; Dondorp, A.M.; Woodrow, C.J. HRP2: Transforming Malaria Diagnosis, but with Caveats. Trends Parasitol. 2020, 36, 112–126. [Google Scholar] [CrossRef] [Green Version]
- Watson, O.J.; Slater, H.C.; Verity, R.; Parr, J.B.; Mwandagalirwa, M.K.; Tshefu, A.; Meshnick, S.R.; Ghani, A.C. Modelling the Drivers of the Spread of Plasmodium falciparum hrp2 Gene Deletions in Sub-Saharan Africa. Elife 2017, 6. [Google Scholar] [CrossRef]
- Gamboa, D.; Ho, M.-F.; Bendezu, J.; Torres, K.; Chiodini, P.; Barnwell, J.W.; Incardona, S.; Perkins, M.; Bell, D.; McCarthy, J.; et al. A Large Proportion of P. falciparum Isolates in the Amazon Region of Peru Lack pfhrp2 and pfhrp3: Implications for Malaria Rapid Diagnostic Tests. PLoS ONE 2010, 5, e8091. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization, Malaria Threats Map. Available online: https://apps.who.int/malaria/maps/threats/ (accessed on 22 February 2023).
- Nair, S.; Li, X.; Nkhoma, S.C.; Anderson, T. Fitness Costs of pfhrp2 and pfhrp3 Deletions Underlying Diagnostic Evasion in Malaria Parasites. J. Infect. Dis. 2022, 226, 1637–1645. [Google Scholar] [CrossRef]
- Koita, O.A.; Doumbo, O.K.; Ouattara, A.; Tall, L.K.; Konaré, A.; Diakité, M.; Diallo, M.; Sagara, I.; Masinde, G.L.; Doumbo, S.N.; et al. False-Negative Rapid Diagnostic Tests for Malaria and Deletion of the Histidine-Rich Repeat Region of the hrp2 Gene. Am. J. Trop. Med. Hyg. 2012, 86, 194–198. [Google Scholar] [CrossRef] [Green Version]
- Pati, P.; Dhangadamajhi, G.; Bal, M.; Ranjit, M. High Proportions of pfhrp2 Gene Deletion and Performance of hrp2-Based Rapid Diagnostic Test in Plasmodium falciparum Field Isolates of Odisha. Malar. J. 2018, 17, 1–11. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. World Malaria Report 2021; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- Li, B.; Sun, Z.; Li, X.; Li, X.; Wang, H.; Chen, W.; Chen, P.; Qiao, M.; Mao, Y. Performance of pfhrp2 Versus pldh Antigen Rapid Diagnostic Tests for the Detection of Plasmodium falciparum: A Systematic Review and Meta-Analysis. Arch. Med Sci. 2017, 13, 541–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. False-Negative RDT Results and Implications of New Reports of P. falciparum Histidine-Rich Protein 2/3 Gene Deletions; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Dechavanne, S.; Dossou, Y.; Hotchihoue, J.; Vianou, B.; Sossou, D.; Zinsou, B.E.; Fernando, A.; Akoho, R.; Massougbodji, A.; Fievet, N.; et al. Deletions in HRP2 Gene in Plasmodium falciparum Isolates in Benin. In Proceedings of the American Journal of Tropical Medicine and Hygiene—Annual Meeting, Virtual Meeting, 17–21 November 2021; Abstract Number 0625. p. 200. [Google Scholar]
- Parr, J.B.; Verity, R.; Doctor, S.M.; Janko, M.; Carey-Ewend, K.; Turman, B.J.; Keeler, C.; Slater, H.C.; Whitesell, A.N.; Mwandagalirwa, K.; et al. Pfhrp2-Deleted Plasmodium falciparum Parasites in the Democratic Republic of Congo: A National Cross-Sectional Survey. J. Infect. Dis. 2016, 216, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Funwei, R.; Nderu, D.; Nguetse, C.N.; Thomas, B.N.; Falade, C.O.; Velavan, T.P.; Ojurongbe, O. Molecular Surveillance of pfhrp2 and pfhrp3 Genes Deletion in Plasmodium falciparum Isolates and the Implications for Rapid Diagnostic Tests in Nigeria. Acta Trop. 2019, 196, 121–125. [Google Scholar] [CrossRef]
- Alemayehu, G.S.; Blackburn, K.; Lopez, K.; Dieng, C.C.; Lo, E.; Janies, D.; Golassa, L. Detection of High Prevalence of Plasmodium falciparum Histidine-Rich Protein 2/3 Gene Deletions in Assosa Zone, Ethiopia: Implication for Malaria Diagnosis. Malar. J. 2021, 20, 1–11. [Google Scholar] [CrossRef]
- Prosser, C.; Gresty, K.; Ellis, J.; Meyer, W.; Anderson, K.; Lee, R.; Cheng, Q. Plasmodium falciparum Histidine-Rich Protein 2 and 3 Gene Deletions in Strains from Nigeria, Sudan, and South Sudan. Emerg. Infect. Dis. 2021, 27, 471–479. [Google Scholar] [CrossRef]
- A Vera-Arias, C.; Holzschuh, A.; O Oduma, C.; Badu, K.; Abdul-Hakim, M.; Yukich, J.; Hetzel, M.W.; Fakih, B.S.; Ali, A.; Ferreira, M.U.; et al. High-Throughput Plasmodium falciparum hrp2 and hrp3 Gene Deletion Typing by Digital PCR to Monitor Malaria Rapid Diagnostic Test Efficacy. Elife 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Kreidenweiss, A.; Trauner, F.; Rodi, M.; Koehne, E.; Held, J.; Wyndorps, L.; Manouana, G.P.; McCall, M.; Adegnika, A.A.; Lalremruata, A.; et al. Monitoring the Threatened Utility of Malaria Rapid Diagnostic Tests by Novel High-Throughput Detection of Plasmodium falciparum hrp2 and hrp3 Deletions: A Cross-Sectional, Diagnostic Accuracy Study. Ebiomedicine 2019, 50, 14–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grignard, L.; Nolder, D.; Sepúlveda, N.; Berhane, A.; Mihreteab, S.; Kaaya, R.; Phelan, J.; Moser, K.; van Schalkwyk, D.A.; Campino, S.; et al. A Novel Multiplex qPCR Assay for Detection of Plasmodium falciparum with Histidine-Rich Protein 2 and 3 (pfhrp2 and pfhrp3) Deletions in Polyclonal Infections. Ebiomedicine 2020, 55, 102757. [Google Scholar] [CrossRef]
- Schindler, T.; Deal, A.C.; Fink, M.; Guirou, E.; Moser, K.A.; Mwakasungula, S.M.; Mihayo, M.G.; Jongo, S.A.; Chaki, P.P.; Abdulla, S.; et al. A Multiplex qPCR Approach for Detection of pfhrp2 and pfhrp3 Gene Deletions in Multiple Strain Infections of Plasmodium falciparum. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Ntabi, J.D.M.; Lissom, A.; Djontu, J.C.; Diafouka-Kietela, S.; Vouvoungui, C.; Boumpoutou, R.K.; Mayela, J.; Nguiffo-Nguete, D.; Nkemngo, F.N.; Ndo, C.; et al. Prevalence of Non-Plasmodium falciparum Species in Southern Districts of Brazzaville in The Republic of the Congo. Parasites Vectors 2022, 15, 1–13. [Google Scholar] [CrossRef]
- Agonhossou, R.; Akoton, R.; Dossou, Y.A.; Avokpaho, E.; Mbama, D.N.J.; Boussougou-Sambe, T.S.; Francis, N.N.; Ndo, C.; Ntoumi, F.; Wondji, C.S.; et al. Surveillance of Plasmodium malariae Infection Among Inhabitants of Rural Areas in Ouidah–Kpomasse–Tori Bossito Health District, Benin. Parasitol. Res. 2022, 121, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Joanny, F.; Löhr, S.J.; Engleitner, T.; Lell, B.; Mordmüller, B. Limit of Blank and Limit of Detection of Plasmodium falciparum Thick Blood Smear Microscopy in a Routine Setting in Central Africa. Malar. J. 2014, 13, 234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plowe, C.V.; Djimde, A.; Bouare, M.; Doumbo, O.; Wellems, T.E. Pyrimethamine and Proguanil Resistance-Conferring Mutations in Plasmodium falciparum Dihydrofolate Reductase: Polymerase Chain Reaction Methods for Surveillance in Africa. Am. J. Trop. Med. Hyg. 1995, 52, 565–568. [Google Scholar] [CrossRef]
- Woldearegai, T.G.; Lalremruata, A.; Nguyen, T.T.; Gmeiner, M.; Veletzky, L.; Tazemda-Kuitsouc, G.B.; Matsiegui, P.B.; Mordmüller, B.; Held, J. Characterization of Plasmodium infections Among Inhabitants of Rural Areas in Gabon. Sci. Rep. 2019, 9, 9784. [Google Scholar] [CrossRef] [Green Version]
- Luu-The, V.; Paquet, N.; Calvo, E.; Cumps, J. Improved Real-Time RT-PCR Method for High-Throughput Measurements Using Second Derivative Calculation and Double Correction. Biotechniques 2005, 38, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, R. Quantification on the LightCycler. In Rapid Cycle Real-Time PCR: Methods and Applications; Meuer, S., Wittwer, C., Nakagawara, K.-I., Eds.; Springer: Berlin/Heidelberg, Germany, 2001; pp. 21–34. [Google Scholar]
- Fleiss, J.L. Measuring Nominal Scale Agreement Among Many Raters. Psychol. Bull. 1971, 76, 378–382. [Google Scholar] [CrossRef]
- Landis, J.R.; Koch, G.G. The Measurement of Observer Agreement for Categorical Data. Biometrics 1977, 33, 159–174. [Google Scholar] [CrossRef] [Green Version]
- Mihreteab, S.; Anderson, K.; Pasay, C.; Smith, D.; Gatton, M.L.; Cunningham, J.; Berhane, A.; Cheng, Q. Epidemiology of Mutant Plasmodium falciparum Parasites Lacking Histidine-Rich Protein 2/3 Genes in Eritrea 2 Years After Switching from hrp2-Based RDTs. Sci. Rep. 2021, 11, 1–12. [Google Scholar] [CrossRef]
- Amoah, L.E.; Abuaku, B.; Bukari, A.H.; Dickson, D.; Amoako, E.O.; Asumah, G.; Asamoah, A.; Preprah, N.Y.; Malm, K.L. Contribution of P. falciparum Parasites with Pfhrp 2 Gene Deletions to False Negative pfhrp 2 Based Malaria RDT Results in Ghana: A Nationwide Study of Symptomatic Malaria Patients. PLoS ONE 2020, 15, e0238749. [Google Scholar] [CrossRef]
- Feleke, S.M.; Reichert, E.N.; Mohammed, H.; Brhane, B.G.; Mekete, K.; Mamo, H.; Petros, B.; Solomon, H.; Abate, E.; Hennelly, C.; et al. Plasmodium falciparum is Evolving to Escape Malaria Rapid Diagnostic Tests in Ethiopia. Nat. Microbiol. 2021, 6, 1289–1299. [Google Scholar] [CrossRef]
- Golassa, L.; Messele, A.; Amambua-Ngwa, A.; Swedberg, G. High Prevalence and Extended Deletions in Plasmodium falciparum hrp2/3 Genomic Loci in Ethiopia. PLoS ONE 2020, 15, e0241807. [Google Scholar] [CrossRef] [PubMed]
- Agaba, B.B.; Yeka, A.; Nsobya, S.; Arinaitwe, E.; Nankabirwa, J.; Opigo, J.; Mbaka, P.; Lim, C.S.; Kalyango, J.N.; Karamagi, C.; et al. Systematic Review of the Status of pfhrp2 and pfhrp3 Gene Deletion, Approaches and Methods Used for its Estimation and Reporting in Plasmodium falciparum Populations in Africa: Review of Published Studies 2010–2019. Malar. J. 2019, 18, 355. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Surveillance Template Protocol for pfhrp2/pfhrp3 Gene Deletions; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]


| Gabon | Congo | Nigeria | Benin | |
|---|---|---|---|---|
| Total N | 534 | 917 | 466 | 120 |
| Age in years: | ||||
| Median (IQR) | 16 (6–25) | 15 (8–38) | * | 9 (5–13) |
| Age cohorts: | ||||
| 0–6 years, n (%) | 77 (14%) | 169 (18%) | - | 45 (38%) |
| 7–18 years, n (%) | 91 (17%) | 334 (36%) | - | 49 (41%) |
| 18+ years, n (%) | 134 (25%) | 412 (45%) | - | 26 (22%) |
| Missing data | 232 (43%) | 2 (0.2%) | 466 (100%) | - |
| Sex: | ||||
| Female | 258 (48%) | 514 (56%) | 75 (63%) | |
| Male | 154 (29%) | 403 (44%) | 45 (38%) | |
| Missing data: | 122 (23%) | - | 466 (100%) | - |
| Symptomatic: | * | 73/914 (8%) | 299 (64%) | 9 (8%) |
| Sampling period: | 2019–2020 | 2021 | 2018–2019 | 2019 |
| Number of study sites: | ||||
| 1 | Lambaréné 534 (100%) | Goma Tsé-Tsé District 573 (62%) | Awka 200 (43%) | Kpomasse 68 (57%) |
| 2 | - | Brazzaville 344 (38%) | Nnewi 167 (36%) | Ouidah 52 (43%) |
| 3 | - | - | Onitsha 99 (21%) | - |
| Measurements: | ||||
| TBS done, n (%) | 534 (100%) | 917 (100%) | not done | 120 (100%) |
| RDT done, n (%) | 532 (99.6%) | 917 (100%) | not available | not done |
| RDT brand: | Paracheck Pf RDT | Malaria P.f./Pan Antigen kit, Cypress Diagnostics | - | - |
| Gabon | Congo | Nigeria | Benin | |
|---|---|---|---|---|
| Total N | 534 | 917 | 466 | 120 |
| TBS positive, n (%) | 96 (18%) | 329 (36%) | - | 120 (100%) |
| Median parasitemia [p/µL] | 475 (145–1849) | 530 (147–5514) | - | 2240 (1008–6900) |
| RDT positive, n (%) | 186/532 * (35%) | 478 (52%) | - | - |
| qPCR positive | 273 (51%) | 643 (70%) | 379 (81%) | 120 (100%) |
| False-negative RDTs vs TBS vs qPCR | 8 91 | 2 186 | - - | - - |
| Kappa | ||||
| RDT/TBS | 0.50 | 0.67 | - | - |
| RDT/qPCR | 0.64 | 0.54 | - | - |
| TBS/qPCR | 0.33 | 0.36 | - | - |
| RDT/TBS/qPCR | 0.47 | 0.50 | - | - |
| Gabon | Republic of Congo | Nigeria | Benin | |
|---|---|---|---|---|
| pfcytb and pfbtub positive, n | 218 | 512 | 316 | 120 |
| pfhrp2-deleted | 2 (1%) | 0 (0%) | 1 (0.03%) | 0 (0%) |
| pfhrp3-deleted | 0 (0%) | 0 (0%) | 1 (0.03%) | 0 (0%) |
| pfhrp2- and pfhrp3-deleted | 0 (0%) | 0 (0%) | 5 (1.6%) | 0 (0%) |
| ID | Country/ Subregion | TBS Result | HRP2-RDT Result | 4plex qPCR | CQR | |||
|---|---|---|---|---|---|---|---|---|
| pfhrp2 Cq | pfhrp3 Cq | pfcytb Cq | pfbtub Cq | Haplotype | ||||
| 1 | Gabon/* | Neg | Neg | del | 38.2 | 34.0 | 35.3 | * |
| 2 | Gabon/* | Neg | Neg | del | 37.1 | 31.7 | 34.5 | * |
| 3 | Nigeria/Onitsha | * | Pos | del | 37.4 | 34.6 | 35.0 | CVIET |
| 4 | Nigeria/Nnewi | * | * | 34.8 | del | 33.9 | 35.3 | * |
| 5 | Nigeria/Nnewi | * | * | del | del | 35.7 | 34.7 | * |
| 6 | Nigeria/Nnewi | * | * | del | del | 36.2 | 35.4 | SVMNT |
| 7 | Nigeria/Onitsha | * | * | del | del | 35.2 | 36.2 | SVMNT |
| 8 | Nigeria/Onitsha | * | * | del | del | 35.1 | 35.5 | CVIET |
| 9 | Nigeria/Awka | * | * | del | del | 31.3 | 37.0 | * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krueger, T.; Ikegbunam, M.; Lissom, A.; Sandri, T.L.; Ntabi, J.D.M.; Djontu, J.C.; Baina, M.T.; Lontchi, R.A.L.; Maloum, M.; Ella, G.Z.; et al. Low Prevalence of Plasmodium falciparum Histidine-Rich Protein 2 and 3 Gene Deletions—A Multiregional Study in Central and West Africa. Pathogens 2023, 12, 455. https://doi.org/10.3390/pathogens12030455
Krueger T, Ikegbunam M, Lissom A, Sandri TL, Ntabi JDM, Djontu JC, Baina MT, Lontchi RAL, Maloum M, Ella GZ, et al. Low Prevalence of Plasmodium falciparum Histidine-Rich Protein 2 and 3 Gene Deletions—A Multiregional Study in Central and West Africa. Pathogens. 2023; 12(3):455. https://doi.org/10.3390/pathogens12030455
Chicago/Turabian StyleKrueger, Tina, Moses Ikegbunam, Abel Lissom, Thaisa Lucas Sandri, Jacques Dollon Mbama Ntabi, Jean Claude Djontu, Marcel Tapsou Baina, Roméo Aimé Laclong Lontchi, Moustapha Maloum, Givina Zang Ella, and et al. 2023. "Low Prevalence of Plasmodium falciparum Histidine-Rich Protein 2 and 3 Gene Deletions—A Multiregional Study in Central and West Africa" Pathogens 12, no. 3: 455. https://doi.org/10.3390/pathogens12030455