
Chalcone Derivative Induces Flagellar Disruption and Autophagic Phenotype in Phytomonas serpens In Vitro

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. NaF Synthesis
2.3. Parasites Culture and the Derivative NaF
2.4. Analysis of Plasma Membrane Integrity
2.5. Analysis of Reactive Oxygen Species (ROS) Production
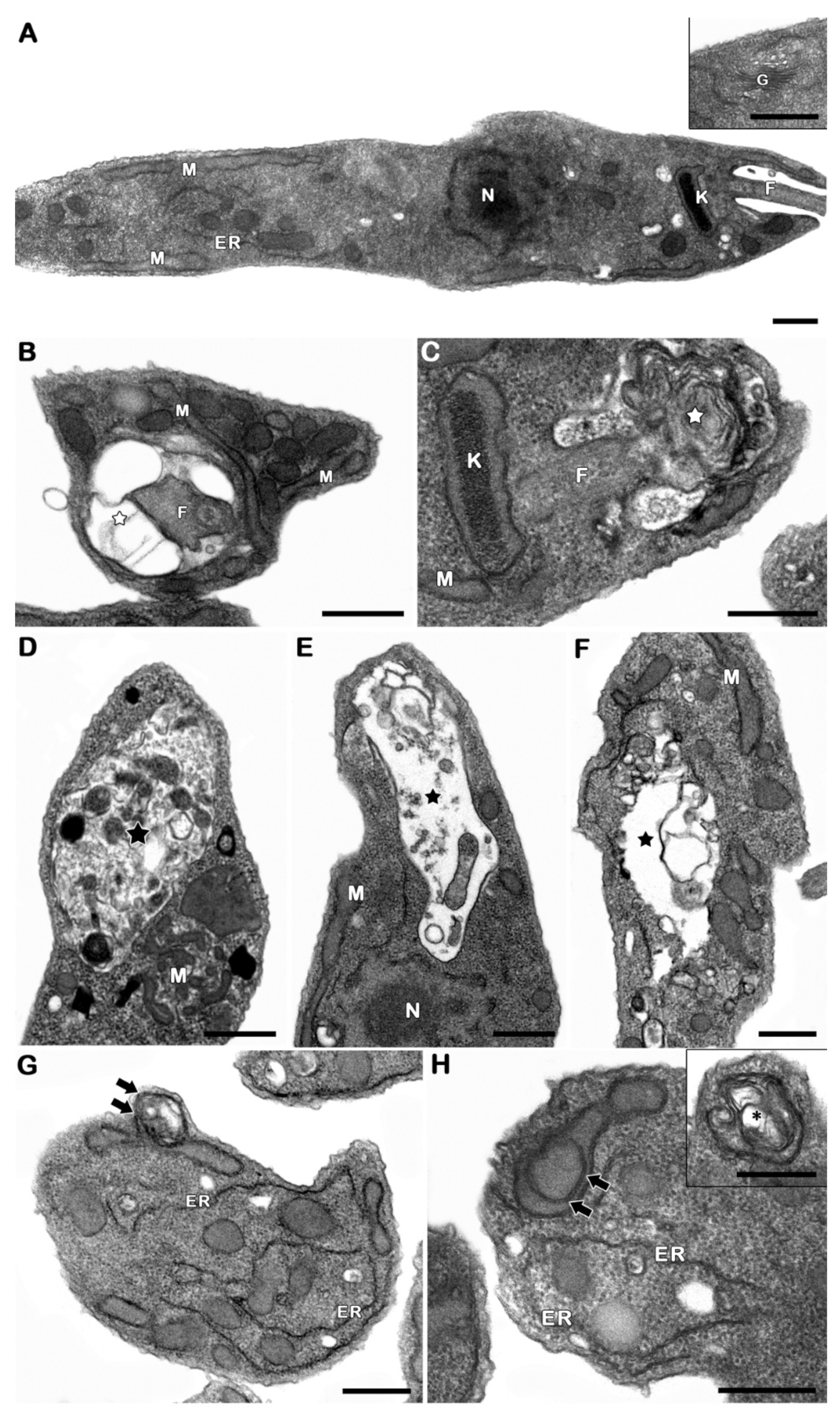
2.6. Ultrastructural Analysis
2.7. Effect of NaF on the Promastigote Flagellar Morphology
2.8. Statistical Analysis
3. Results
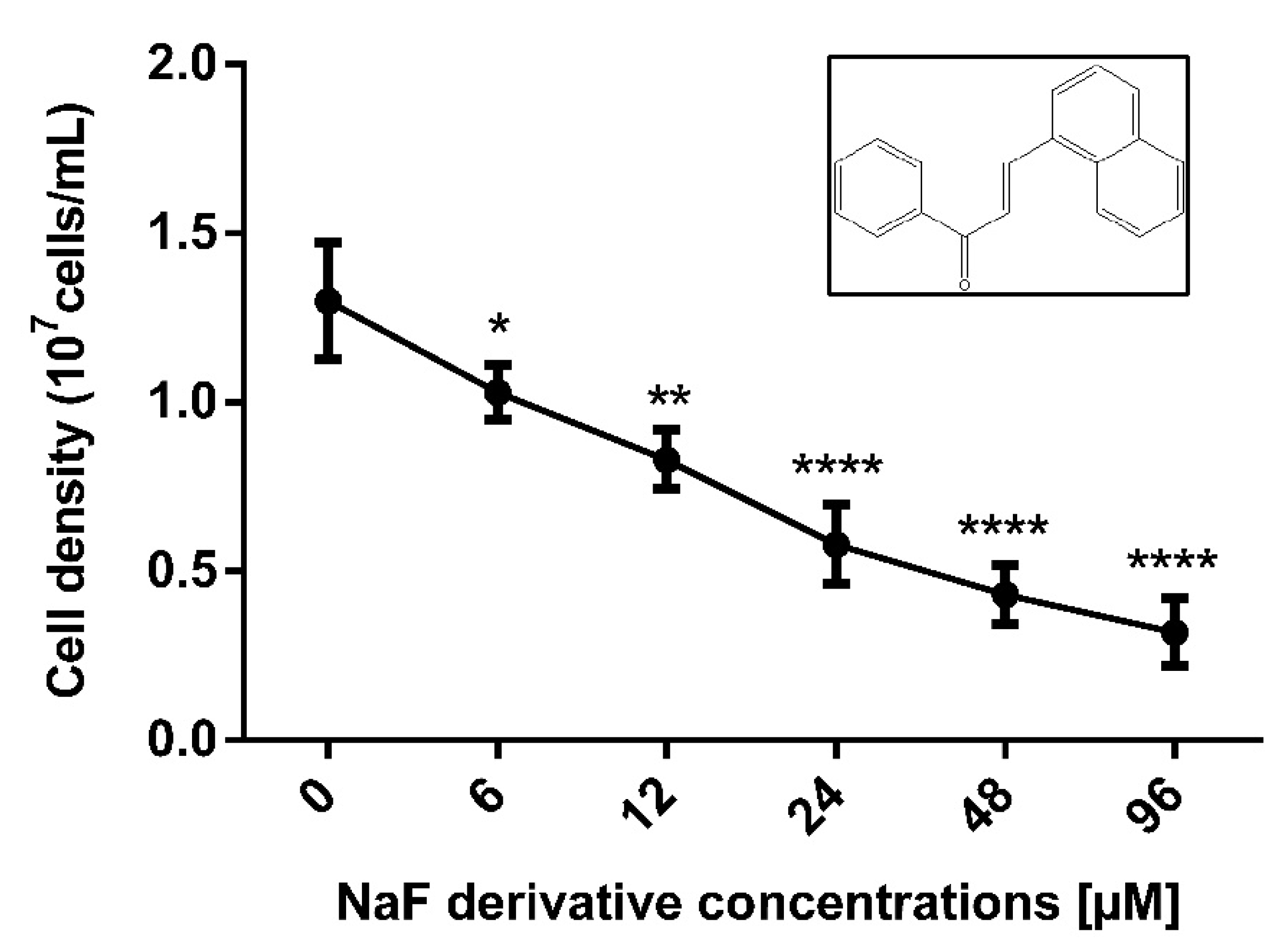
3.1. NaF Affects P. serpens Promastigotes Proliferation
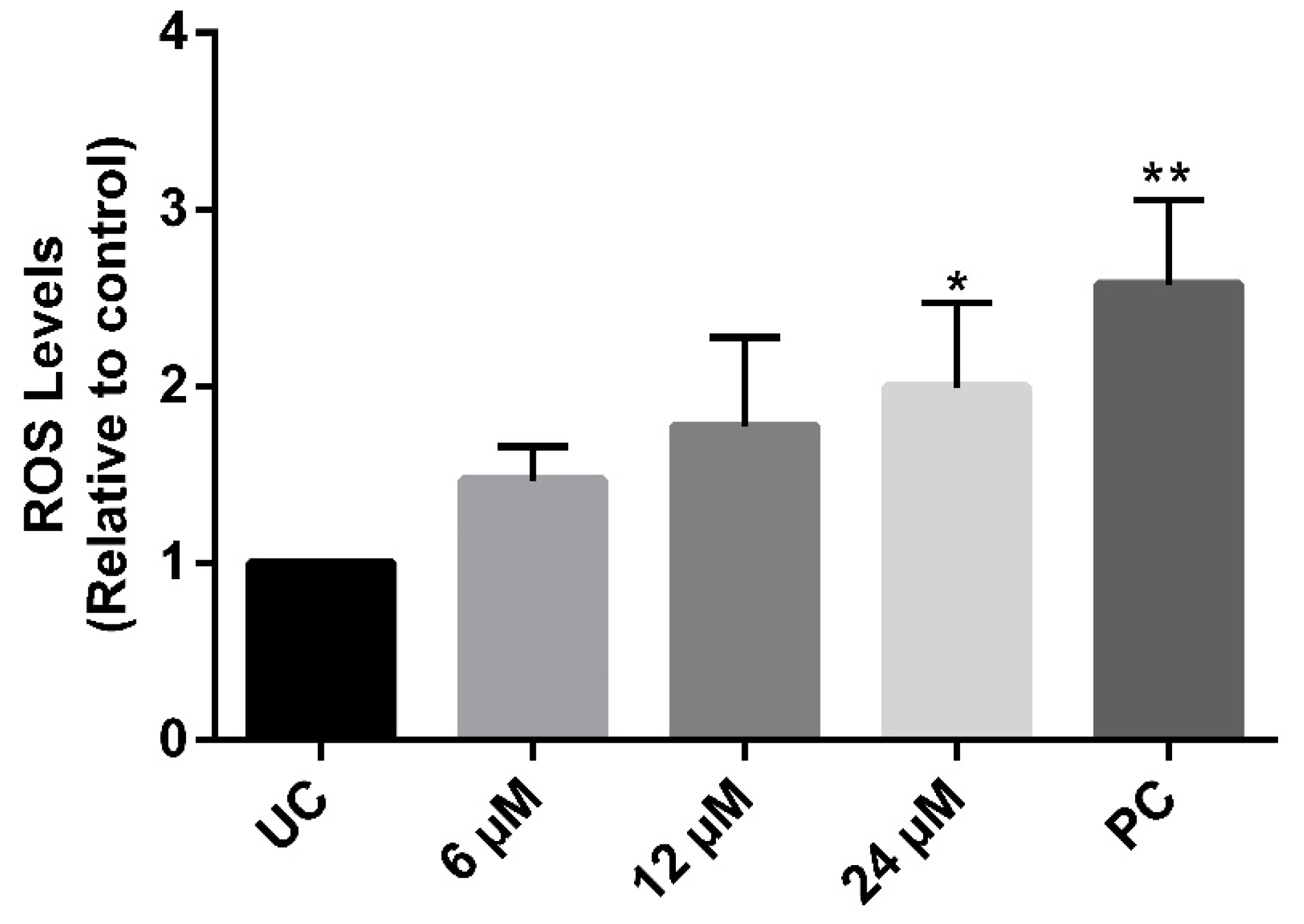
3.2. The Compound NaF Induces an Increase in ROS Production without Affecting Plasma Membrane Integrity
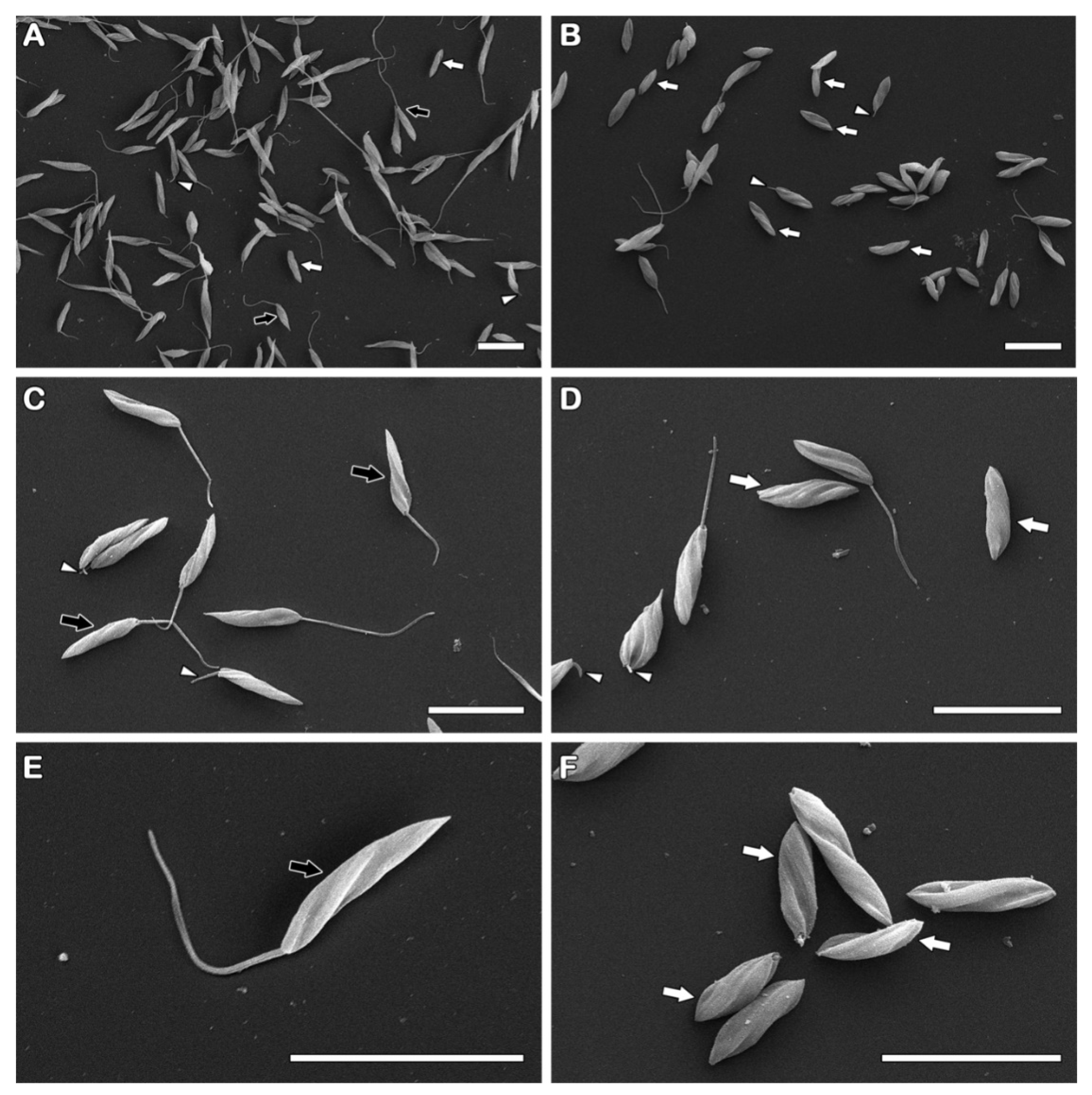
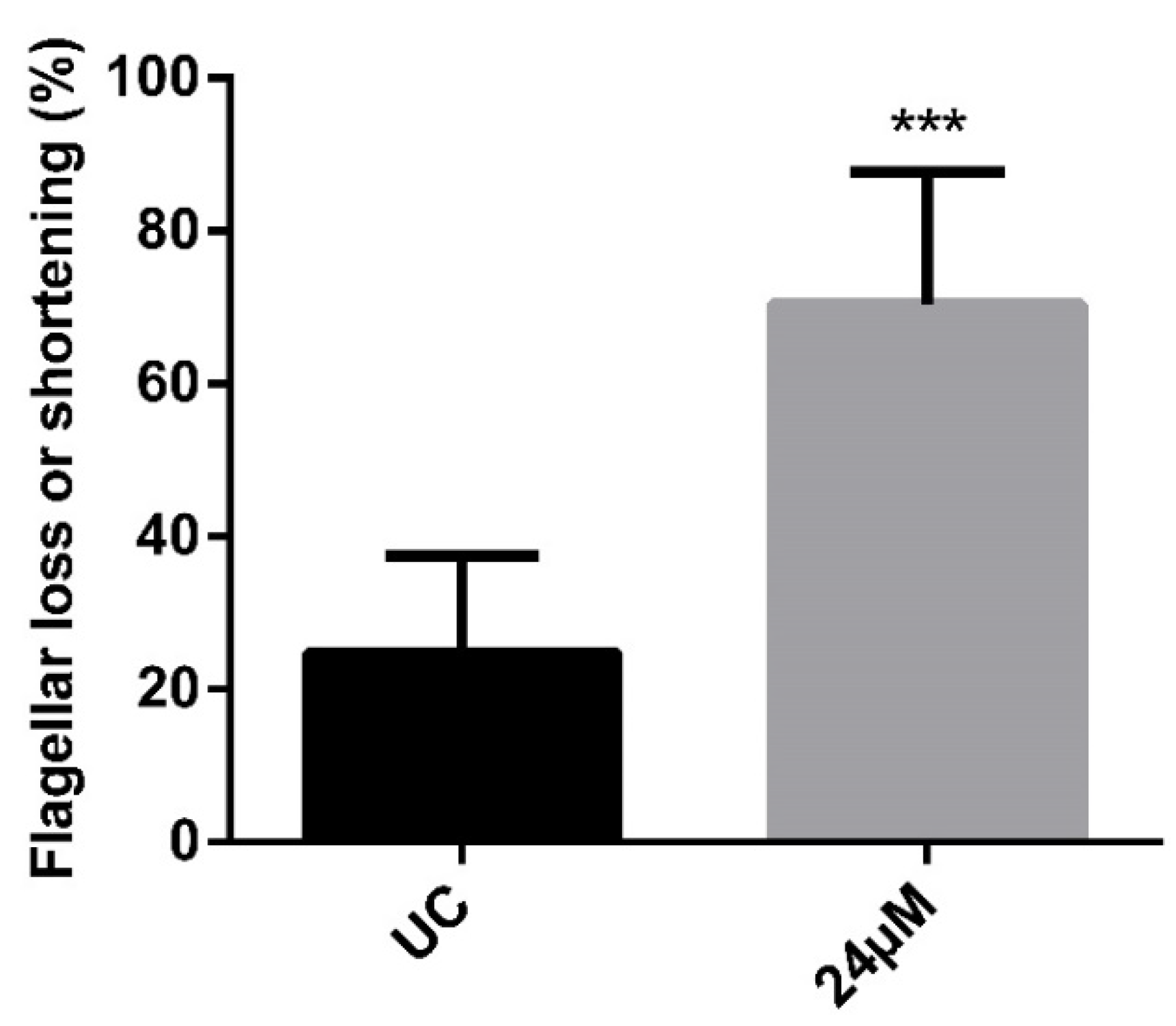
3.3. The Derivative NaF Induces Flagellarloss and/or Shortening
3.4. The Derivative NaF Promoted the Appearance of Autophagic Phenotype in P. serpens
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schwelm, A.; Badstöber, J.; Bulman, S.; Desoignies, N.; Etemadi, M.; Falloon, R.E.; Gachon, C.M.M.; Legreve, A.; Lukeš, J.; Merz, U.; et al. Not in Your Usual Top 10: Protists That Infect Plants and Algae. Mol. Plant Pathol. 2018, 19, 1029–1044. [Google Scholar] [CrossRef] [Green Version]
- Jaskowska, E.; Butler, C.; Preston, G.; Kelly, S. Phytomonas: Trypanosomatids Adapted to Plant Environments. PLoS Pathog. 2015, 11, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camargo, E.P. Phytomonas and Other Trypanosomatid Parasites of Plants and Fruit. Adv. Parasitol. 1999, 42, 29–112. [Google Scholar] [PubMed]
- Santos, A.L.S.; Branquinha, M.H.; D’Avila-Levy, C.M. The Ubiquitous Gp63-like Metalloprotease from Lower Trypanosomatids: In the Search for a Function. An. Acad. Bras. Cienc. 2006, 78, 687–714. [Google Scholar] [CrossRef] [Green Version]
- Santos, A.L.S.; d’Avila-Levy, C.M.; Elias, C.G.R.; Vermelho, A.B.; Branquinha, M.H. Phytomonas serpens: Immunological Similarities with the Human Trypanosomatid Pathogens. Microbes Infect. 2007, 9, 915–921. [Google Scholar] [CrossRef] [PubMed]
- Porcel, B.M.; Denoeud, F.; Opperdoes, F.; Noel, B.; Madoui, M.A.; Hammarton, T.C.; Field, M.C.; Da Silva, C.; Couloux, A.; Poulain, J.; et al. The Streamlined Genome of Phytomonas spp. Relative to Human Pathogenic Kinetoplastids Reveals a Parasite Tailored for Plants. PLoS Genet. 2014, 10, e1004007. [Google Scholar] [CrossRef]
- Dos Santos, A.D.C.M., Jr.; Ricart, C.A.O.; Pontes, A.H.; Fontes, W.; De Souza, A.R.; Castro, M.S.; De Sousa, M.V.; De Lima, B.D. Proteome Analysis of Phytomonas serpens, a Phytoparasite of Medical Interest. PLoS ONE 2018, 13, 1–15. [Google Scholar] [CrossRef]
- Melo, J.O.d.; Bitencourt, T.A.; Fachin, A.L.; Cruz, E.M.O.; de Jesus, H.C.R.; Alves, P.B.; Arrigoni-Blank, M.d.F.; de Castro Franca, S.; Beleboni, R.O.; Fernandes, R.P.M.; et al. Antidermatophytic and Antileishmanial Activities of Essential Oils from Lippia gracilis Schauer Genotypes. Acta Trop. 2013, 128, 110–115. [Google Scholar] [CrossRef]
- Silva, K.P.; Santos, T.A.d.C.; Moutinho, B.L.; da Silva, R.S.; dos Santos Pinto, V.; Blank, A.F.; Corrêa, C.B.; Scher, R.; Fernandes, R.P.M. Using Varronia curassavica (Cordiaceae) Essential Oil for the Biocontrol of Phytomonas serpens. Ind. Crops Prod. 2019, 139, 111523. [Google Scholar] [CrossRef]
- Pereira, K.L.G.; Vasconcelos, N.B.R.; Braz, J.V.C.; Inácio, J.D.F.; Estevam, C.S.; Correa, C.B.; Fernandes, R.P.M.; Almeida-Amaral, E.E.; Scher, R. Ethanolic Extract of Croton blanchetianus Ball Induces Mitochondrial Defects in Leishmania amazonensis Promastigotes. An. Acad. Bras. Cienc. 2020, 92, e20180968. [Google Scholar] [CrossRef]
- Pereira, K.L.G.; Nizio, D.A.d.C.; de Lima, P.C.N.; Fernandes, R.P.M.; Arrigoni-Blank, M.d.F.; de Sá Filho, J.C.F.; Nascimento, L.F.d.A.; de Souza, V.T.; Silva, K.P.; Blank, A.F. Seasonal Variance in the Chemical Composition of Essential Oils from Lantana camara Accessions and Their Trypanocidal Activity on Phytomonas serpens. Bol. Latinoam. Caribe Plantas Med. Aromat. 2022, 21, 737–756. [Google Scholar] [CrossRef]
- Farias-Junior, A.P.; Rios, M.C.; Moura, T.A.; Almeida, R.P.; Alves, P.B.; Blank, A.F.; Fernandes, R.P.M.; Scher, R. Leishmanicidal Activity of Carvacrol-Rich Essential Oil from Lippia sidoides. Cham. Biol. Res. 2012, 45, 399–402. [Google Scholar] [CrossRef] [Green Version]
- Medina, J.M.; Rodrigues, J.C.F.; Moreira, O.C.; Atella, G.; De Souza, W.; Barrabin, H. Mechanisms of Growth Inhibition of Phytomonas serpens by the Alkaloids Tomatine and Tomatidine. Mem. Inst. Oswaldo Cruz. 2015, 110, 48–55. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.R.S.T.; Scher, R.; Santos, F.V.; Ferreira, S.R.; Cavalcanti, S.C.H.; Correa, C.B.; Bueno, L.L.; Alves, R.J.; Souza, D.P.; Fujiwara, R.T.; et al. Leishmanicidal Activity and Structure-Activity Relationships of Essential Oil Constituents. Molecules. 2017, 22, 815. [Google Scholar] [CrossRef] [Green Version]
- Fournet, A.; Munoz, V. Natural Products as Trypanocidal, Antileishmanial and Antimalarial. Drugs. Curr. Top. Med. Chem. 2002, 2, 1215–1237. [Google Scholar] [CrossRef]
- Ferreira, M.K.A.; Fontenelle, R.O.S.; Magalhães, F.E.A.; Bandeira, P.N.; De Menezes, J.S.E.A.; Dos Santos, H.S. Chalcones Pharmacological Potential: A Brief Review. Rev. Virtual Quim. 2018, 10, 1455–1473. [Google Scholar] [CrossRef]
- Rozmer, Z.; Perjési, P. Naturally Occurring Chalcones and Their Biological Activities. Phytochem. Rev. 2016, 15, 87–120. [Google Scholar] [CrossRef]
- Yazdan, S.K.; Sagar, D.V.; Shaik, A.B. Organic & Medicinal Chemistry Chemical and Biological Potentials of Chalcones: A Review. Org. Med. Chem. Int. J. 2015, 1, 555553. [Google Scholar]
- Pola, S.; Banoth, K.K.; Sankaranarayanan, M.; Ummani, R.; Garlapati, A. Design, Synthesis, in Silico Studies, and Evaluation of Novel Chalcones and Their Pyrazoline Derivatives for Antibacterial and Antitubercular Activities. Med. Chem. Res. 2020, 29, 1819–1835. [Google Scholar] [CrossRef]
- Ortalli, M.; Ilari, A.; Colotti, G.; De Ionna, I.; Battista, T.; Bisi, A.; Gobbi, S.; Rampa, A.; Di Martino, R.M.C.; Gentilomi, G.A.; et al. Identification of Chalcone-Based Antileishmanial Agents Targeting Trypanothione Reductase. Eur. J. Med. Chem. 2018, 152, 527–541. [Google Scholar] [CrossRef]
- De Mello, T.F.P.; Bitencourt, H.R.; Pedroso, R.B.; Aristides, S.M.A.; Lonardoni, M.V.C.; Silveira, T.G. V Leishmanicidal Activity of Synthetic Chalcones in Leishmania (Viannia) braziliensis. Exp. Parasitol. 2014, 136, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Santiago-Silva, K.M.d.; Bortoleti, B.T.d.S.; Oliveira, L.d.N.; Maia, F.L.d.A.; Castro, J.C.; Costa, I.C.; Lazarin, D.B.; Wardell, J.L.; Wardell, S.M.S.V.; Albuquerque, M.G.; et al. Antileishmanial Activity of 4,8-Dimethoxynaphthalenyl Chalcones on Leishmania amazonensis. Antibiotics 2022, 11, 1402. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, E.P.; Gomes, N.D.B.; Freitas, T.A.d.; Silva, B.P.; Ribeiro, L.R.; Ameida-Neto, F.W.Q.; Marinho, M.M.; Lima-Neto, P.d.; Marinho, E.S.; Santos, H.S.d.; et al. Chloride Substitution on 2-Hydroxy-3,4,6-Trimethoxyphenyl chalcones Improves in vitro Selectivity on Trypanosoma cruzi Strain Y. Chem. Biol. Interact. 2022, 361, 109920. [Google Scholar] [CrossRef] [PubMed]
- Boniface, P.K.; Ferreira, E.I.; Fabrice, F.B. The Role of Flavanones as Scaffolds for the Development of New Treatments against Malaria and African and American Trypanosomiases. Mini-Rev. Med. Chem. 2022, in press. [Google Scholar] [CrossRef]
- Zhai, L.; Chen, M.; Blom, J.; Theander, T.G.; Christensen, S.B.; Kharazmi, A. The Antileishmanial Activity of Novel Oxygenated Chalcones and Their Mechanism of Action. J. Antimicrob. Chemother. 1999, 43, 793–803. [Google Scholar] [CrossRef] [Green Version]
- Mello, T.F.P.d.; Cardoso, B.M.; Bitencourt, H.R.; Donatti, L.; Aristides, S.M.A.; Lonardoni, M.V.C.; Silveira, T.G.V. Ultrastructural and Morphological Changes in Leishmania (Viannia) braziliensis Treated with Synthetic Chalcones. Exp. Parasitol. 2016, 160, 23–30. [Google Scholar] [CrossRef]
- Chen, M.; Zhai, L.; Christensen, S.B.; Theander, T.G.; Kharazmi, A. Inhibition of Fumarate Reductase in Leishmania major and L. donovani by Chalcones. Antimicrob. Agents Chemother. 2001, 45, 2023–2029. [Google Scholar] [CrossRef] [Green Version]
- Torres-Santos, E.C.; Sampaio-Santos, M.I.; Buckner, F.S.; Yokoyama, K.; Gelb, M.; Urbina, J.A.; Rossi-Bergmann, B. Altered Sterol Profile Induced in Leishmania amazonensis by a Natural Dihydroxymethoxylated Chalcone. J. Antimicrob. Chemother. 2009, 63, 469–472. [Google Scholar] [CrossRef] [Green Version]
- Souza, G.B.; Santos, T.A.C.; Silva, A.P.S.; Barreiros, A.L.B.S.; Nardelli, V.B.; Siqueira, I.B.; Dolabella, S.S.; Costa, E.V.; Alves, P.B.; Scher, R.; et al. Synthesis of Chalcone Derivatives by Claisen-Schmidt Condensation and in Vitro Analyses of Their Antiprotozoal Activities. Nat. Prod. Res. 2022, 1–4. [Google Scholar] [CrossRef]
- Araújo, J.C.A.d.; Pereira, J.C.R.; Gasparotto, L. Murcha-de-Phytomonas Do Coqueiro No Amazonas. Circ. Técnica 2003, 17, 1–6. [Google Scholar]
- Díaz-Tielas, C.; Graña, E.; Reigosa, M.J.; Sánchez-Moreiras, A.M. Biological Activities and Novel Applications of Chalcones. Planta Daninha. 2016, 34, 607–616. [Google Scholar] [CrossRef] [Green Version]
- Espinoza-Hicks, J.C.; Chacón-Vargas, K.F.; Hernández-Rivera, J.L.; Nogueda-Torres, B.; Tamariz, J.; Sánchez-Torres, L.E.; Camacho-Dávila, A. Novel Prenyloxy Chalcones as Potential Leishmanicidal and Trypanocidal Agents: Design, Synthesis and Evaluation. Eur. J. Med. Chem. 2019, 167, 402–413. [Google Scholar] [CrossRef]
- Leeza Zaidi, S.; Mittal, S.; Rajala, M.S.; Avecilla, F.; Husain, M.; Azam, A. Synthesis, Characterization and Antiamoebic Activity of Chalcones Bearing N -Substituted Ethanamine Tail. Eur. J. Med. Chem. 2015, 98, 179–189. [Google Scholar] [CrossRef]
- Chiaradia, L.D.; Mascarello, A.; Purificação, M.; Vernal, J.; Cordeiro, M.N.S.; Zenteno, M.E.; Villarino, A.; Nunes, R.J.; Yunes, R.A.; Terenzi, H. Synthetic Chalcones as Efficient Inhibitors of Mycobacterium tuberculosis Protein Tyrosine Phosphatase PtpA. Bioorgan. Med. Chem. Lett. 2008, 18, 6227–6230. [Google Scholar] [CrossRef]
- Cabrera, M.; Cerecetto, H.; González, M. New Hybrid Bromopyridine-Chalcones as in vivo Phase II Enzyme Inducers: Potential Chemopreventive Agents. Med. Chem. Comm. 2016, 7, 2395–2409. [Google Scholar] [CrossRef]
- Attar, S.; O’Brien, Z.; Alhaddad, H.; Golden, M.L.; Calderón-Urrea, A. Ferrocenyl Chalcones versus Organic Chalcones: A Comparative Study of Their Nematocidal Activity. Bioorgan. Med. Chem. 2011, 19, 2055–2073. [Google Scholar] [CrossRef]
- Singh, J.V.; Sharma, S.; Rahar, S. Synthesis and Spermicidal Activity of Substituted (E)-3-(Aryl/Heteroaryl)-1-Phenylprop-2-En-1-Ones. Der. Pharm. Chem. 2015, 7, 93–103. [Google Scholar]
- Upegui, Y.; Gil, J.F.; Quiñones, W.; Torres, F.; Escobar, G.; Robledo, S.M.; Echeverri, F. Preparation of Rotenone Derivatives and in Vitro Analysis of Their Antimalarial, Antileishmanial and Selective Cytotoxic Activities. Molecules 2014, 19, 18911–18922. [Google Scholar] [CrossRef] [Green Version]
- Pedra-Rezende, Y.; Macedo, I.S.; Midlej, V.; Mariante, R.M.; Menna-Barreto, R.F.S. Different Drugs, Same End: Ultrastructural Hallmarks of Autophagy in Pathogenic Protozoa. Front. Microbiol. 2022, 13, 856686. [Google Scholar] [CrossRef]
- Escrivani, D.O.; Charlton, R.L.; Caruso, M.B.; Burle-Caldas, G.A.; Borsodi, M.P.G.; Zingali, R.B.; Arruda-Costa, N.; Palmeira-Mello, M.V.; De Jesus, J.B.; Souza, A.M.T.; et al. Chalcones Identify cTXNPx as a Potential Antileishmanial Drug Target. PLoS Negl. Trop. Dis. 2021, 15, 1–23. [Google Scholar] [CrossRef]
- Bombaça, A.C.S.; Von Dossow, D.; Barbosa, J.M.C.; Paz, C.; Burgos, V.; Menna-Barreto, R.F.S. Trypanocidal Activity of Natural Sesquiterpenoids Involves Mitochondrial Dysfunction, ROS Production and Autophagic Phenotype in Trypanosoma cruzi. Molecules 2018, 23, 2800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonseca-Silva, F.; Canto-Cavalheiro, M.M.; Menna-Barreto, R.F.S.; Almeida-Amaral, E.E. Effect of Apigenin on Leishmania amazonensis Is Associated with Reactive Oxygen Species Production Followed by Mitochondrial Dysfunction. J. Nat. Prod. 2015, 78, 880–884. [Google Scholar] [CrossRef] [PubMed]
- Bombaça, A.C.S.; Viana, P.G.; Santos, A.C.C.; Silva, T.L.; Rodrigues, A.B.M.; Guimarães, A.C.R.; Goulart, M.O.F.; da Silva, E.N., Jr.; Menna-Barreto, R.F.S. Mitochondrial Disfunction and ROS Production Are Essential for Anti-Trypanosoma cruzi Activity of β-Lapachone-Derived Naphthoimidazoles. Free Radic. Biol. Med. 2019, 130, 408–418. [Google Scholar] [CrossRef] [PubMed]
- Bombaça, A.C.S.; Silva, L.A.; Chaves, O.A.; da Silva, L.S.; Barbosa, J.M.C.; da Silva, A.M.; Ferreira, A.B.B.; Menna-Barreto, R.F.S. Novel N,N-Di-Alkylnaphthoimidazolium Derivative of β-Lapachone Impaired Trypanosoma cruzi Mitochondrial Electron Transport System. Biomed. Pharmacother. 2021, 135, 111186. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, M.C.; Da Silva, E.N.; Pinto, A.V.; De Castro, S.L.; Menna-Barreto, R.F.S. A Novel Triazolic Naphthofuranquinone Induces Autophagy in Reservosomes and Impairment of Mitosis in Trypanosoma cruzi. Parasitology 2012, 139, 26–36. [Google Scholar] [CrossRef]
- Nawathean, P.; Maslov, D.A. The Absence of Genes for Cytochrome c Oxidase and Reductase Subunits in Maxicircle Kinetoplast DNA of the Respiration-Deficient Plant Trypanosomatid Phytomonas serpens. Curr. Genet. 2000, 38, 95–103. [Google Scholar] [CrossRef]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants Maintain Cellular Redox Homeostasis by Elimination of Reactive Oxygen Species. Cell. Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef]
- Beltrame-Botelho, I.T.; Talavera-López, C.; Andersson, B.; Grisard, E.C.; Stoco, P.H. A Comparative In Silico Study of the Antioxidant Defense Gene Repertoire of Distinct Lifestyle Trypanosomatid Species. Evol. Bioinform. 2016, 12, 263–275. [Google Scholar] [CrossRef] [Green Version]
- Irigoín, F.; Cibils, L.; Comini, M.A.; Wilkinson, S.R.; Flohé, L.; Radi, R. Insights into the Redox Biology of Trypanosoma cruzi: Trypanothione Metabolism and Oxidant Detoxification. Free Radic. Biol. Med. 2008, 45, 733–742. [Google Scholar] [CrossRef]
- Mohamed, M.F.A.; Abuo-Rahma, G.E.D.A. Molecular Targets and Anticancer Activity of Quinoline-Chalcone Hybrids: Literature Review. RSC Adv. 2020, 10, 31139–31155. [Google Scholar] [CrossRef]
- Shin, S.Y.; Lee, J.M.; Lee, M.S.; Koh, D.; Jung, H.; Lim, Y.; Lee, Y.H. Targeting Cancer Cells via the Reactive Oxygen Species-Mediated Unfolded Protein Response with a Novel Synthetic Polyphenol Conjugate. Clin. Cancer Res. 2014, 20, 4302–4313. [Google Scholar] [CrossRef] [Green Version]
- D’Oliveira, G.D.C.; Custodio, J.M.F.; Moura, A.F.; Napolitano, H.B.; Pérez, C.N.; Moraes, M.O.; Prókai, L.; Perjési, P. Different Reactivity to Glutathione but Similar Tumor Cell Toxicity of Chalcones and Their Quinolinone Analogues. Med. Chem. Res. 2019, 28, 1448–1460. [Google Scholar] [CrossRef] [Green Version]
- Barrias, E.; Reignault, L.C.; Calogeropoulou, T.; de Souza, W. In Vitro Activities of Adamantylidene-Substituted Alkylphosphocholine TCAN26 against Trypanosoma cruzi: Antiproliferative and Ultrastructural Effects. Exp. Parasitol. 2019, 206, 107730. [Google Scholar] [CrossRef]
- Menna-Barreto, R.F.S.; Henriques-Pons, A.; Pinto, A.V.; Morgado-Diaz, J.A.; Soares, M.J.; De Castro, S.L. Effect of a β-Lapachone-Derived Naphthoimidazole on Trypanosoma cruzi: Identification of Target Organelles. J. Antimicrob. Chemother. 2005, 56, 1034–1041. [Google Scholar] [CrossRef]
- Da Silva, A.M.; Araújo-Silva, L.; Bombaça, A.C.S.; Menna-Barreto, R.F.S.; Rodrigues-Santos, C.E.; Buarque Ferreira, A.B.; De Castro, S.L. Synthesis and Biological Evaluation of N-Alkyl Naphthoimidazoles Derived from β-Lapachone against Trypanosoma cruzi Bloodstream Trypomastigotes. Med. Chem. Comm. 2017, 8, 952–959. [Google Scholar] [CrossRef]
- Reggiori, F.; Klionsky, D.J. Autophagy in the Eukaryotic Cell. Eukaryot Cell. 2002, 1, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Menna-Barreto, R.F.S. Cell Death Pathways in Pathogenic Trypanosomatids: Lessons of (over)Kill. Cell Death Dis. 2019, 10, 93. [Google Scholar] [CrossRef] [Green Version]
- Salomão, K.; De Santana, N.A.; Molina, M.T.; De Castro, S.L.; Menna-Barreto, R.F.S. Trypanosoma cruzi Mitochondrial Swelling and Membrane Potential Collapse as Primary Evidence of the Mode of Action of Naphthoquinone Analogues. BMC Microbiol. 2013, 13, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, J.H.d.S.; Miranda, N.; Volpato, H.; Ueda-Nakamura, T.; Nakamura, C.V. The Antidepressant Clomipramine Induces Programmed Cell Death in Leishmania amazonensis through a Mitochondrial Pathway. Parasitol. Res. 2019, 118, 977–989. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, T.A.C.; Silva, K.P.; Souza, G.B.; Alves, P.B.; Menna-Barreto, R.F.S.; Scher, R.; Fernandes, R.P.M. Chalcone Derivative Induces Flagellar Disruption and Autophagic Phenotype in Phytomonas serpens In Vitro. Pathogens 2023, 12, 423. https://doi.org/10.3390/pathogens12030423
Santos TAC, Silva KP, Souza GB, Alves PB, Menna-Barreto RFS, Scher R, Fernandes RPM. Chalcone Derivative Induces Flagellar Disruption and Autophagic Phenotype in Phytomonas serpens In Vitro. Pathogens. 2023; 12(3):423. https://doi.org/10.3390/pathogens12030423
Chicago/Turabian StyleSantos, Tamiris A. C., Kleiton P. Silva, Gabriella B. Souza, Péricles B. Alves, Rubem F. S. Menna-Barreto, Ricardo Scher, and Roberta P. M. Fernandes. 2023. "Chalcone Derivative Induces Flagellar Disruption and Autophagic Phenotype in Phytomonas serpens In Vitro" Pathogens 12, no. 3: 423. https://doi.org/10.3390/pathogens12030423