
Trapping Behaviour of Duddingtonia flagrans against Gastrointestinal Nematodes of Cattle under Year-Round Grazing Conditions
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
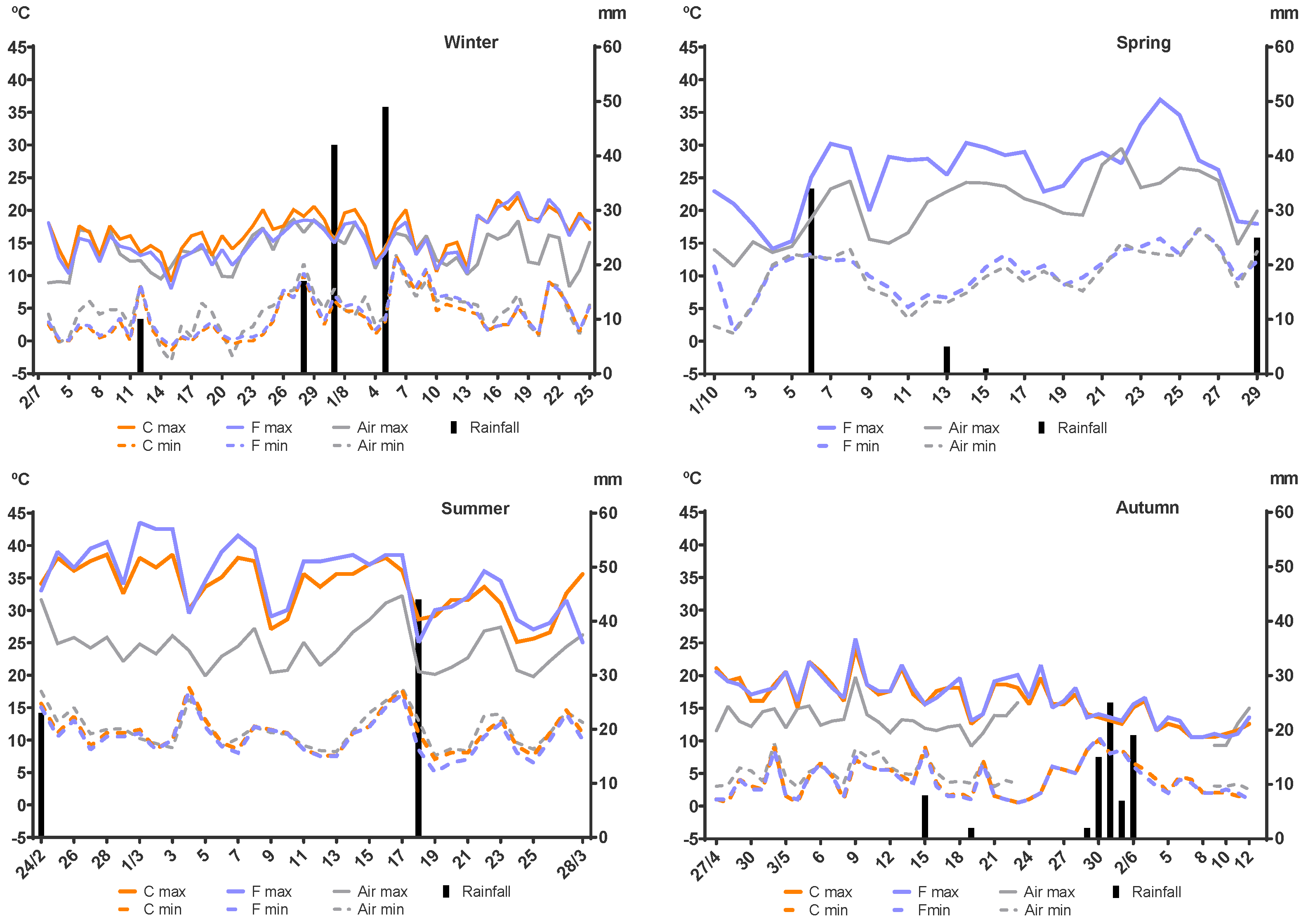
2.1. Experimental Site
2.2. Experimental Design
2.3. Faecal Cultures
2.4. Herbage Sampling
2.5. Faecal Pats Recovering
2.6. Experimental Setup Differences Due to Seasonality
2.7. Fungal Material
2.8. Parasitological Procedures
2.9. Statistical Analysis
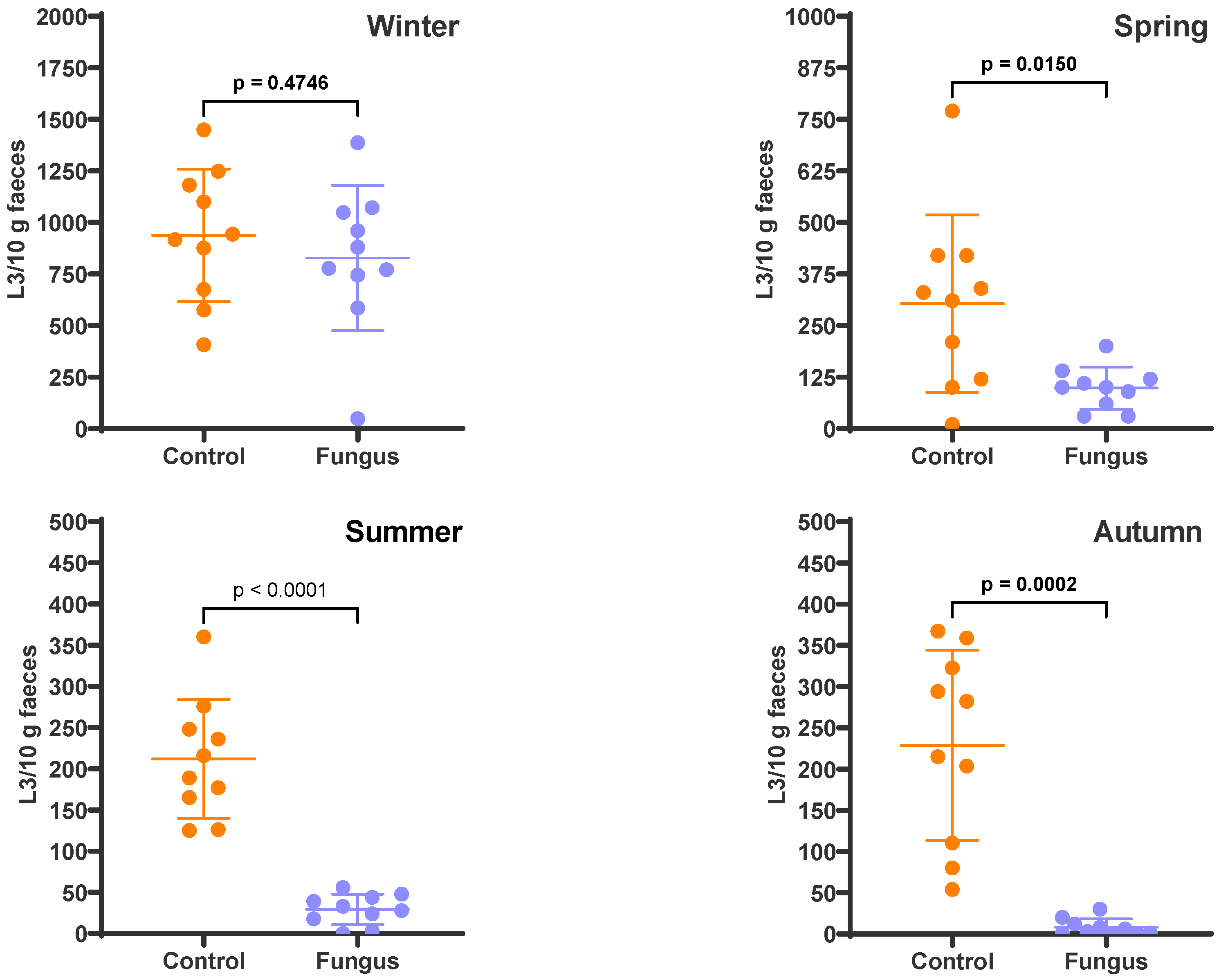
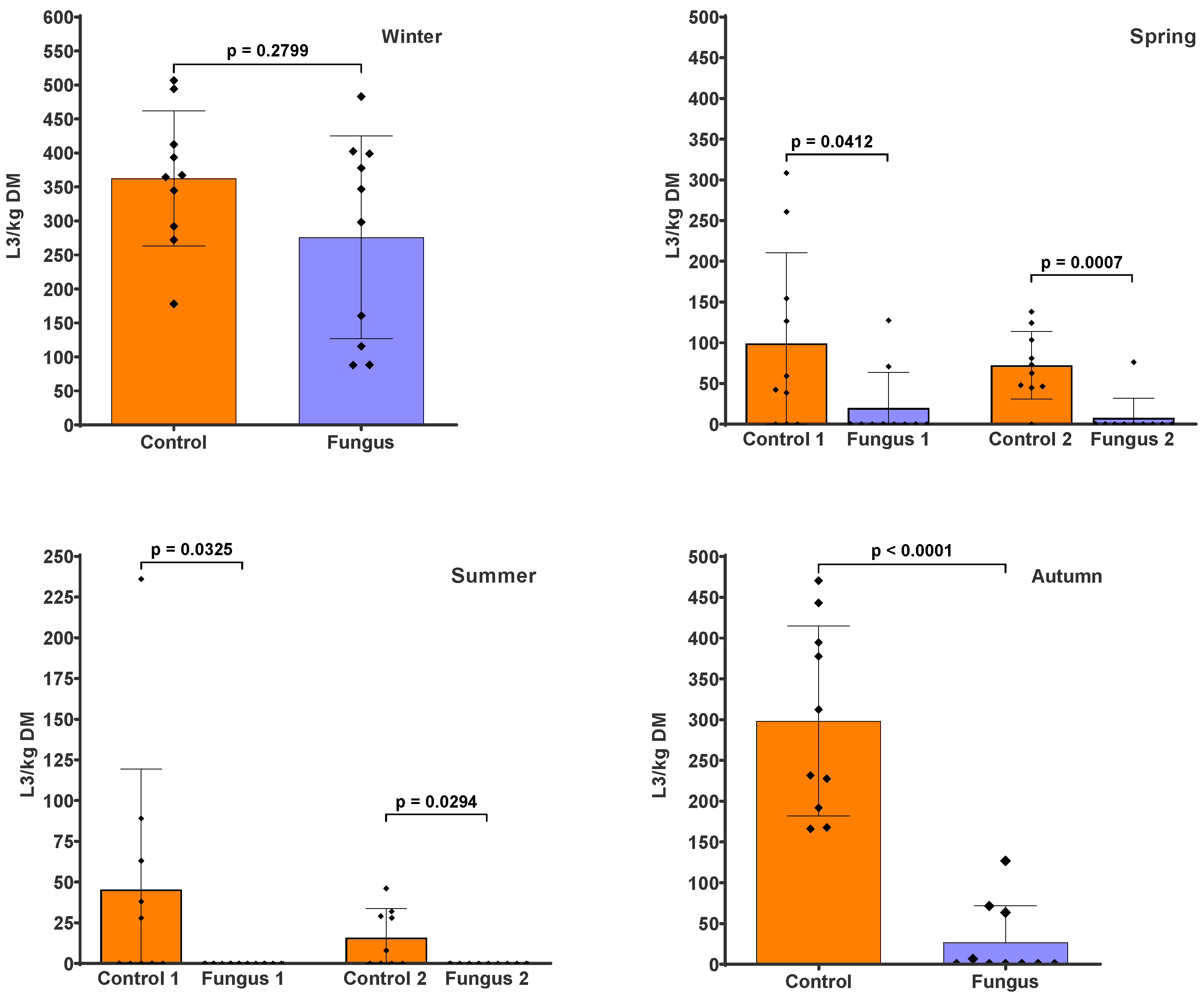
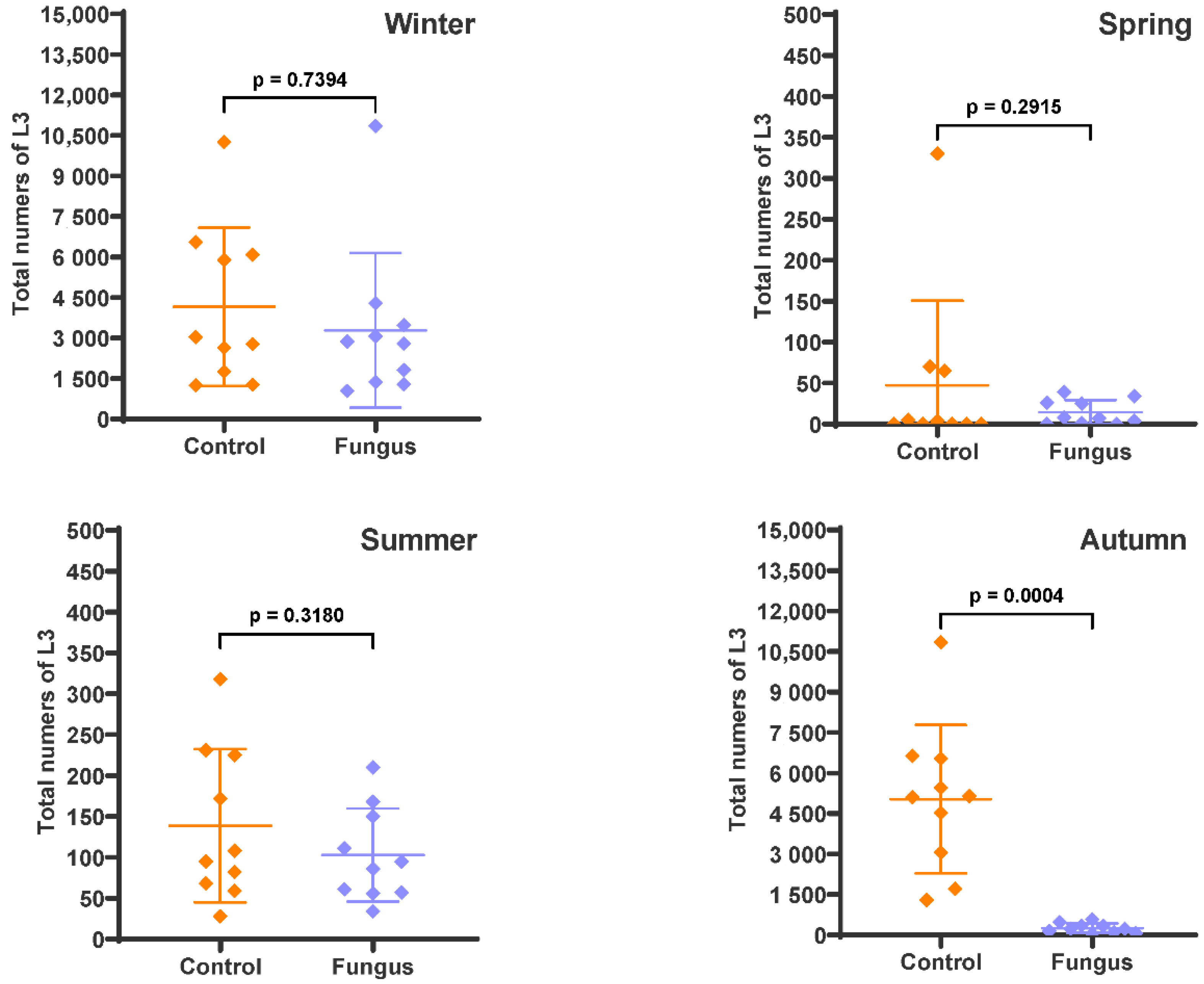
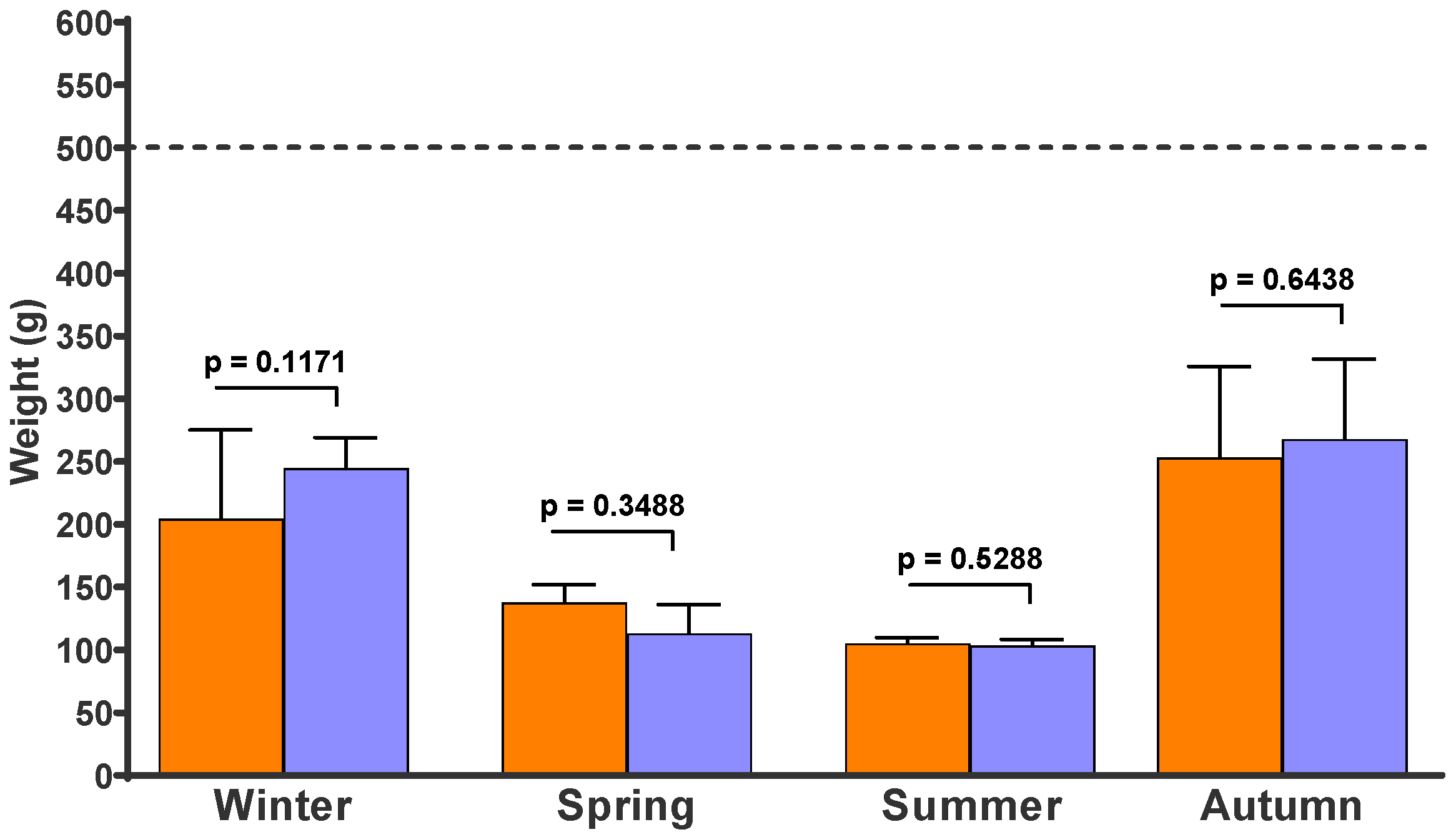
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cotter, J.L.; Van Burgel, A.; Besier, R.B. Anthelmintic resistance in nematodes of beef cattle in south-west Western Australia. Vet. Parasitol. 2015, 207, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Ramos, F.; Portella, L.P.; Rodrigues, F.S.; Reginato, C.Z.; Pötter, L.; Cezar, A.S.; Sangioni, L.A.; Vogel, F.S.F. Anthelmintic resistance in gastrointestinal nematodes of beef cattle in the state of Rio Grande do Sul, Brazil. Int. J. Parasitol. Drugs Drug Resist. 2016, 6, 93–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cristel, S.; Fiel, C.; Anziani, O.; Descarga, C.; Cetrá, B.; Romero, J.; Fernández, S.; Entrocasso, C.; Lloberas, M.; Medus, D.; et al. Anthelmintic resistance in grazing beef cattle in central and northeastern areas of Argentina—An update. Vet. Parasitol. Reg. Stud. Rep. 2017, 9, 25–28. [Google Scholar] [CrossRef] [PubMed]
- Vineer, R.H.; Morgan, E.R.; Hertzberg, H.; Bartley, D.J.; Bosco, A.; Charlier, J.; Chartier, C.; Claerebout, E.; de Waal, T.; Hendrickx, G.; et al. Increasing importance of anthelmintic resistance in European livestock: Creation and meta-analysis of an open database. Parasite 2020, 27, 69. [Google Scholar] [CrossRef] [PubMed]
- World Organisation for Animal Health (OIE). Responsible and Prudent Use of Anthelmintic Chemicals to Help Control Anthelmintic Resistance in Grazing Livestock Species; OIE: Paris, France, 2021; p. 38. [Google Scholar]
- Iglesias, L.; Saumell, C.A.; Fernández, A.S.; Fusé, C.A.; Lifschitz, A.L.; Rodríguez, E.M.; Steffan, P.E.; Fiel, C.A. Environmental impact of ivermectin excreted by cattle treated in autumn on dung fauna and degradation of feces on pasture. Parasitol. Res. 2006, 100, 93–102. [Google Scholar] [CrossRef]
- Iglesias, L.E.; Sallovitz, J.; Saumell, C.A.; Fernández, A.S.; Junco, M.; Sagües, M.F.; Zegbi, S.; Lifschitz, A. Uptake of ivermectin from growing substrate to plant species. In Proceedings of the 27th International Conference of the World Association for the Advancement of Veterinary Parasitology, Madison, WI, USA, 7–12 July 2019. [Google Scholar]
- Lumaret, J.P.; Errouissi, F.; Floate, K.; Römbke, J.; Wardhaugh, K. A review on the toxicity and non-target effects of macrocyclic lactones in terrestrial and aquatic environments. Curr. Pharm. Biotechnol. 2012, 13, 1004–1060. [Google Scholar] [CrossRef] [Green Version]
- Wagil, M.; Białk-Bielińska, A.; Puckowski, A.; Wychodnik, K.; Maszkowska, J.; Mulkiewicz, E.; Kumirska, J.; Stepnowski, P.; Stolte, S. Toxicity of anthelmintic drugs (fenbendazole and flubendazole) to aquatic organisms. Environ. Sci. Pollut. Res. Int. 2015, 22, 2566–2573. [Google Scholar] [CrossRef] [Green Version]
- De Souza, R.B.; Guimarães, J.R. Effects of avermectins on the environment based on its toxicity to plants and soil invertebrates—A review. Water Air Soil Pollut. 2022, 233, 259. [Google Scholar] [CrossRef]
- Hoste, H.; Torres-Acosta, J.F. Non chemical control of helminths in ruminants: Adapting solutions for changing worms in a changing world. Vet. Parasitol. 2011, 180, 144–154. [Google Scholar] [CrossRef]
- Šimpraga, M.; Ljubičić, I.; Hlede, J.P.; Vugrovečki, A.S.; Marinculić, A.; Tkalčić, S. Alternative approaches for the control of gastrointestinal nematodes in sheep farming: A review. Berl. Munch. Tierarztl. Wochenschr. 2015, 128, 257–270. [Google Scholar]
- Grønvold, J.; Henriksen, S.A.; Larsen, M.; Nansen, P.; Wolstrup, J. Biological control. Aspects of biological control, with special reference to arthropods, protozoans and helminths of domesticated animals. Vet. Parasitol. 1996, 64, 47–64. [Google Scholar] [CrossRef]
- Fernández, A.S.; Larsen, M.; Nansen, P.; Grønvold, J.; Henriksen, S.A.; Bjørn, H.; Wolstrup, J. The efficacy of two isolates of the nematode-trapping fungus Duddingtonia flagrans against Dictyocaulus viviparus larvae in faeces. Vet. Parasitol. 1999, 85, 289–304. [Google Scholar] [CrossRef]
- Faedo, M.; Larsen, M.; Dimander, S.O.; Yeates, G.W.; Höglund, J.; Waller, P.J. Growth of the fungus Duddingtonia flagrans in soil surrounding feces deposited by cattle or sheep fed the fungus to control nematode parasites. Biol. Contr. 2002, 23, 64–70. [Google Scholar] [CrossRef]
- Knox, M.R.; Josh, P.F.; Anderson, L.J. Deployment of Duddingtonia flagrans in an improved pasture system: Dispersal, persistence, and effects on free-living soil nematodes and microarthropods. Biol. Contr. 2002, 24, 176–182. [Google Scholar] [CrossRef]
- Yeates, G.W.; Skipp, R.A.; Chen, L.Y.; Waghorn, T.S.; Potter, J.F. Temporal and spatial aspects of the influence on soil nematodes of depositing artificial pats of sheep faeces containing a range of parasite management agents. Appl. Soil Ecol. 2007, 37, 106–117. [Google Scholar] [CrossRef]
- Paraud, C.; Lorrain, R.; Pors, I.; Chartier, C. Administration of the nematophagous fungus Duddingtonia flagrans to goats: An evaluation of the impact of this fungus on the degradation of faeces and on free-living nematodes. J. Helminthol. 2012, 86, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Saumell, C.A.; Fernández, A.S.; Echevarria, F.; Gonçalves, I.; Iglesias, M.E.; Sagües, M.F. Lack of negative effects of the biological control agents Duddingtonia flagrans on soil nematodes and other nematophagous fungi Rodríguez, M.E. J. Helminthol. 2016, 90, 706–711. [Google Scholar] [CrossRef] [PubMed]
- Fernández, A.S.; Larsen, M.; Henningsen, E.; Nansen, P.; Grønvold, J.; Bjørn, H.; Wolstrup, J. Effect of Duddingtonia flagrans against Ostertagia ostertagi in cattle grazing at different stocking rates. Parasitology 1999, 119, 105–111. [Google Scholar] [CrossRef]
- Dimander, S.-O.; Höglund, J.; Uggla, A.; Spörndly, E.; Waller, P.J. Evaluation of gastro-intestinal nematode parasite control strategies for first-season grazing cattle in Sweden. Vet. Parasitol. 2003, 111, 193–209. [Google Scholar] [CrossRef] [PubMed]
- Waller, P.J.; Schwan, O.; Ljungström, B.-L.; Rydzik, A.; Yeates, G.W. Evaluation of biological control of sheep parasites using Duddingtonia flagrans under commercial farming conditions on the island of Gotland, Sweden. Vet. Parasitol. 2004, 126, 299–315. [Google Scholar] [CrossRef]
- Chandrawathani, P.; Jamnah, O.; Adnan, M.; Waller, P.J.; Larsen, M.; Gillespie, A.T. Field studies on the biological control of nematode parasites of sheep in the tropics, using the microfungus Duddingtonia flagrans. Vet. Parasitol. 2004, 120, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Ortiz Pérez, D.O.; Sánchez Muñoz, B.; Nahed Toral, J.; Orantes Zebadúa, M.Á.; Cruz López, J.L.; Reyes García, M.E.; Mendoza de Gives, P. Using Duddingtonia flagrans in calves under an organic milk farm production system in the Mexican tropics. Exp. Parasitol. 2017, 175, 74–78. [Google Scholar] [CrossRef] [PubMed]
- Luns, F.D.; Assis, R.C.L.; Silva, L.P.C.; Ferraz, C.M.; Braga, F.R.; de Araújo, J.V. Coadministration of nematophagous fungi for biological control over nematodes in bovine in the South-Eastern Brazil. BioMed Res. 2018, 2018, 2934674. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, L.d.S.S.C.B.; Dias, F.G.S.; Melo, A.L.T.; de Carvalho, L.M.; Silva, E.N.; Araújo, J.V. Bioverm® in the control of nematodes in beef cattle raised in the Central-West region of Brazil. Pathogens 2021, 10, 548. [Google Scholar] [CrossRef] [PubMed]
- Buss Baiak, B.H.; de Sousa, K.T.; Deniz, M.; Gasparina, J.M.; Ianke, L.; Pereira, L.M.; Araújo, J.V.; Rocha, R.A.; Dittrich, J.R. Predation of the fungus Duddingtonia flagrans on infective larvae of gastrointestinal nematodes from heifers in a silvopastoral system under shaded and sunny conditions. J. Helminthol. 2022, 96, e20. [Google Scholar] [CrossRef]
- Bilotto, F.; Fusé, L.A.; Sagües, M.F.; Iglesias, L.E.; Fernández, A.S.; Zegbi, S.; Guerrero, I.; Saumell, C.A. Predatory effect of Duddingtonia flagrans on infective larvae of gastro-intestinal parasites under sunny and shaded conditions. Exp. Parasitol. 2018, 193, 27–32. [Google Scholar] [CrossRef]
- Healey, K.; Lawlor, C.; Knox, M.R.; Chambers, M.; Lamb, J.; Groves, P. Field evaluation of Duddingtonia flagrans IAH 1297 for the reduction of worm burden in grazing animals: Pasture larval studies in horses, cattle and goats. Vet. Parasitol 2018, 258, 124–132. [Google Scholar] [CrossRef]
- Beck, H.E.; Zimmermann, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and future Köppen-Geiger climate classification maps at 1-km resolution. Sci. Data 2018, 5, 180214. [Google Scholar] [CrossRef] [Green Version]
- Pereyra, F.X. Ecoregiones de la Argentina; Servicio Geológico Minero Argentino: Buenos Aires, Argentina, 2003; pp. 1–208. [Google Scholar]
- Servicio Meteorológico Nacional, Centro de Información Meteorológica. Available online: https://www.smn.gob.ar/estadisticas (accessed on 10 February 2023).
- Fernández, A.S.; Larsen, M.; Nansen, P.; Grønvold, J.; Henriksen, S.A.; Wolstrup, J. Effect of the nematode-trapping fungus Duddingtonia flagrans on the free-living stages of horse parasitic nematodes: A plot study. Vet. Parasitol. 1997, 73, 257–266. [Google Scholar] [CrossRef]
- Fiel, C.A.; Fernández, A.S.; Rodríguez, E.; Fusé, C.A.; Steffan, P.E. Observations on the free-living stages of cattle gastrointestinal nematodes. Vet. Parasitol. 2012, 187, 217–226. [Google Scholar] [CrossRef]
- Saumell, C.A.; Fernández, A.S.; Fusé, L.A.; Rodríguez, M.; Sagües, M.F.; Iglesias, M.E. Nematophagous fungi from decomposing cattle faeces in Argentina. Rev. Iberoam. Micol. 2015, 32, 252–256. [Google Scholar] [CrossRef]
- Sagües, M.F.; Fusè, L.A.; Iglesias, L.E.; Moreno, F.C.; Saumell, C.A. Optimization of production of chlamydospores of the nematode-trapping fungus Duddingtonia flagrans in solid culture media. Parasitol. Res. 2012, 112, 1047–1051. [Google Scholar] [CrossRef]
- Fiel, C.A.; Steffan, P.E.; Ferreyra, D.A. Diagnóstico de las Parasitosis Más Frecuentes de los Rumiantes: Técnicas de Diagnóstico e Interpretación de los Resultados; Abad Benjamín: Tandil, Argentina, 2011; pp. 1–131. [Google Scholar]
- Henriksen, S.A.; Korsholm, H.A. Method for culture and recovery of gastrointestinal strongyle larvae. Nord. Vet. Med. 1983, 35, 429–430. [Google Scholar]
- Ministry of Agriculture, Fisheries and Food (MAFF). Manual of Veterinary Parasitological Laboratory Techniques; Reference Book 418; HMSO: London, UK, 1986; pp. 1–160. [Google Scholar]
- Grønvold, J. Isolation of Cooperia spp. larvae from soil samples. Acta vet. Scand. 1984, 25, 139–142. [Google Scholar]
- Niec, A. Cultivo e Identificación de Larvas Infectantes de Nematodes Gastrointestinales del Bovino y Ovino; INTA Boletín Técnico N° 5; Instituto Nacional de Tecnología Agropecuaria: Buenos Aires, Argentina, 1968; pp. 1–28. [Google Scholar]
- Van Wyk, J.A.; Mayhew, E. Morphological identification of parasitic nematode infective larvae of small ruminants and cattle: A practical lab guide. Onderstepoort J. Vet. Res. 2013, 80, 539. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Zhang, N.; Gong, P.; Li, J.; Yang, J.; Zhang, X.; Cai, K. Effect of temperature, pH, physical and chemical factors on germination rate of the chlamydospores of the nematophagous fungus Duddingtonia flagrans. FEMS Microbiol. Lett. 2019, 366, fnz212. [Google Scholar] [CrossRef]
- Fiel, C.; Steffan, P.; Entrocasso, C. Epidemiología e impacto productivo de nematodos en la Pampa Húmeda. In Enfermedades Parasitarias de Importancia Clínica y Productiva en Rumiantes; Fiel, C., Nari, A., Eds.; Fundamentos Epidemiológicos para su Prevención y Control; Editorial Hemisferio Sur, SRL: Montevideo, Uruguay, 2013; pp. 29–58. [Google Scholar]
- Zegbi, S. Eficacia de Duddingtonia Flagrans Sobre Larvas Pre-Parasíticas de Rumiantes em Verano. Undergraduate Thesis, Universidad Nacional del Centro de la Provincia de Buenos Aires, Tandil, Argentina, 2015. [Google Scholar]
- Peloille, M. Selection of nematode-trapping fungi for use in biological control. In Working Group “Integrated Control of Soil Pests”, “Methods for Studying Nematophagous Fungi”; Kerry, B.R., Crump, D.H., Eds.; IOBC/WPRS Bulletin; IOBC: Zurich, Switzerland, 1991; Volume 14, pp. 13–17. [Google Scholar]
- Grønvold, J.; Nansen, P.; Henriksen, S.A.; Larsen, M.; Wolstrup, J.; Bresciani, J.; Fribert, L. Induction of traps by Ostertagia ostertagi larvae, chlamydospore production and growth rate in the nematode-trapping fungus Duddingtonia flagrans. J. Helminthol. 1996, 70, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Fernández, A.S.; Larsen, M.; Wolstrup, J.; Grønvold, J.; Nansen, P.; Bjørn, H. Growth rate and trapping efficacy of nematode-trapping fungi under constant and fluctuating temperatures. Parasitol. Res. 1999, 85, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Kahn, L.P.; Norman, T.M.; Walkden-Brown, S.W.; Crampton, A.; O’Connor, L.J. Trapping efficacy of Duddingtonia flagrans at temperatures existing at lambing in Australia. Vet. Parasitol. 2007, 146, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Sagües, F.; Saumell, C.; Fusé, L.; Iglesias, L.; Fiel, C. Evaluación de la capacidad predadora de Duddingtonia flagrans en materia fecal expuesta al sol y a la sombra. Acta Bioq. Clín. Latinoam. Buenos. Aires. 2009, 2009 (Suppl. 1), 96. [Google Scholar]
- Zegbi, S.; Sagües, F.; Saumell, C.; Guerrero, I.; Iglesias, L.; Fernández, S. In vitro efficacy of different concentrations of Duddingtonia flagrans on varying egg densities of gastrointestinal nematodes of cattle. Exp. Parasitol. 2021, 230, 108156. [Google Scholar] [CrossRef]
- Persson, L. Studies on the survival of eggs and infective larvae of Ostertagia ostertagi and Cooperia oncophora in herbage. Zbl. Vet. Med. B 1974, 21, 787–798. [Google Scholar] [CrossRef] [PubMed]
- Thieltges, D.W.; Jensen, K.T.; Poulin, R. The role of biotic factors in the transmission of free-living endohelminth stages. Parasitology 2008, 135, 407–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández, S.; Saumell, C. Biological Control of Parasites in Veterinary Medicine. In Fungi: Types, Environmental Impact and Role in Disease; Paz Silva, A., Arias Vázquez, M.S., Eds.; Nova Science Publishers Inc.: New York, NY, USA, 2013; pp. 309–340. [Google Scholar]
- Li, J.; Zou, C.; Xu, J.; Ji, X.; Niu, X.; Yang, J.; Huang, X.; Zhang, K.Q. Molecular mechanisms of nematode-nematophagous microbe interactions: Basis for biological control of plant-parasitic nematodes. Annu. Rev. Phytopathol. 2015, 53, 67–95. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Winter | Spring | Summer | Autumn | |||||
|---|---|---|---|---|---|---|---|---|
| Control | Fungus | Control | Fungus | Control | Fungus | Control | Fungus | |
| Ostertagia | 33.1 (8.452) | 33.6 (4.695) | 63.1 (16.84) | 68.44 (15.07) | 48.8 (9.762) | 49.56 (21.93) | 31.92 (8.835) | 54.29 (34.83) |
| Haemonchus | 13.7 (3.234) | 11.4 (4.624) | 16.64 (11.54) | 4.74 (6.888) | 15.2 (10.46) | 22.56 (16.91) | 18.56 (5.688) | 16.55 (21.33) |
| Trichostrongylus | 22.5 (7.487) | 22.2 (15.01) | 16.57 (10.04) | 5.37 (5.852) | 21.2 (6.812) | 11.78 (16.0) | 18.75 (10.65) | 13.44 (12.30) |
| Cooperia | 30.7 (3.622) | 32.8 (13.34) | 2.55 (3.951) | 19.9 (13.21) | 10.8 (4.237) | 4.33 (5.431) | 22.95 (9.555) | 13.13 (16.77) |
| Oesophagostomum | 0 | 0 | 1.15 (1.498) | 1.55 (3.259) | 4.0 (3.266) | 11.78 (13.26) | 7.82 (3.297) | 2.60 (4.943) |
| Winter | Spring | Summer | Autumn | |||||
|---|---|---|---|---|---|---|---|---|
| Control | Fungus | Control | Fungus | Control | Fungus | Control | Fungus | |
| Ostertagia | 47.5 (5.206) | 48.63 (9.086) | 37.02 (23.33) | 52.19 (27.00) | 41.5 (12.01) | 53.16 (13.23) | 29.72 (5.414) | 35.65 (15.03) |
| Haemonchus | 3.67 (3.445) | 3.5 (2.976) | 17.96 (11.76) | 10.29 (11.63) | 18.64 (12.65) | 10.85 (9.386) | 16.26 (3.094) | 11.39 (11.20) |
| Trichostrongylus | 4.0 (4.899) | 9.13 (4.704) | 4.98 (6.943) | 6.59 (6.539) | 18.12 (12.67) | 19.75 (14.97) | 23.51 (8.352) | 15.63 (10.87) |
| Cooperia | 28.5 (4.461) | 36.75 (5.339) | 31.24 (17.71) | 29.96 (14.71) | 12.42 (11.53) | 7.92 (7.883) | 30.51 (5.616) | 37.33 (16.76) |
| Oesophagostomum | 0 | 0 | 5.72 (8.501) | 0.98 (1.806) | 9.3 (8.830) | 8.32 (7.108) | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández, S.; Zegbi, S.; Sagües, F.; Iglesias, L.; Guerrero, I.; Saumell, C. Trapping Behaviour of Duddingtonia flagrans against Gastrointestinal Nematodes of Cattle under Year-Round Grazing Conditions. Pathogens 2023, 12, 401. https://doi.org/10.3390/pathogens12030401
Fernández S, Zegbi S, Sagües F, Iglesias L, Guerrero I, Saumell C. Trapping Behaviour of Duddingtonia flagrans against Gastrointestinal Nematodes of Cattle under Year-Round Grazing Conditions. Pathogens. 2023; 12(3):401. https://doi.org/10.3390/pathogens12030401
Chicago/Turabian StyleFernández, Silvina, Sara Zegbi, Federica Sagües, Lucía Iglesias, Inés Guerrero, and Carlos Saumell. 2023. "Trapping Behaviour of Duddingtonia flagrans against Gastrointestinal Nematodes of Cattle under Year-Round Grazing Conditions" Pathogens 12, no. 3: 401. https://doi.org/10.3390/pathogens12030401