
Discovery of the Role of Tick Salivary Glands in Enhancement of Virus Transmission—Beginning of an Exciting Story


Abstract
:1. Introduction
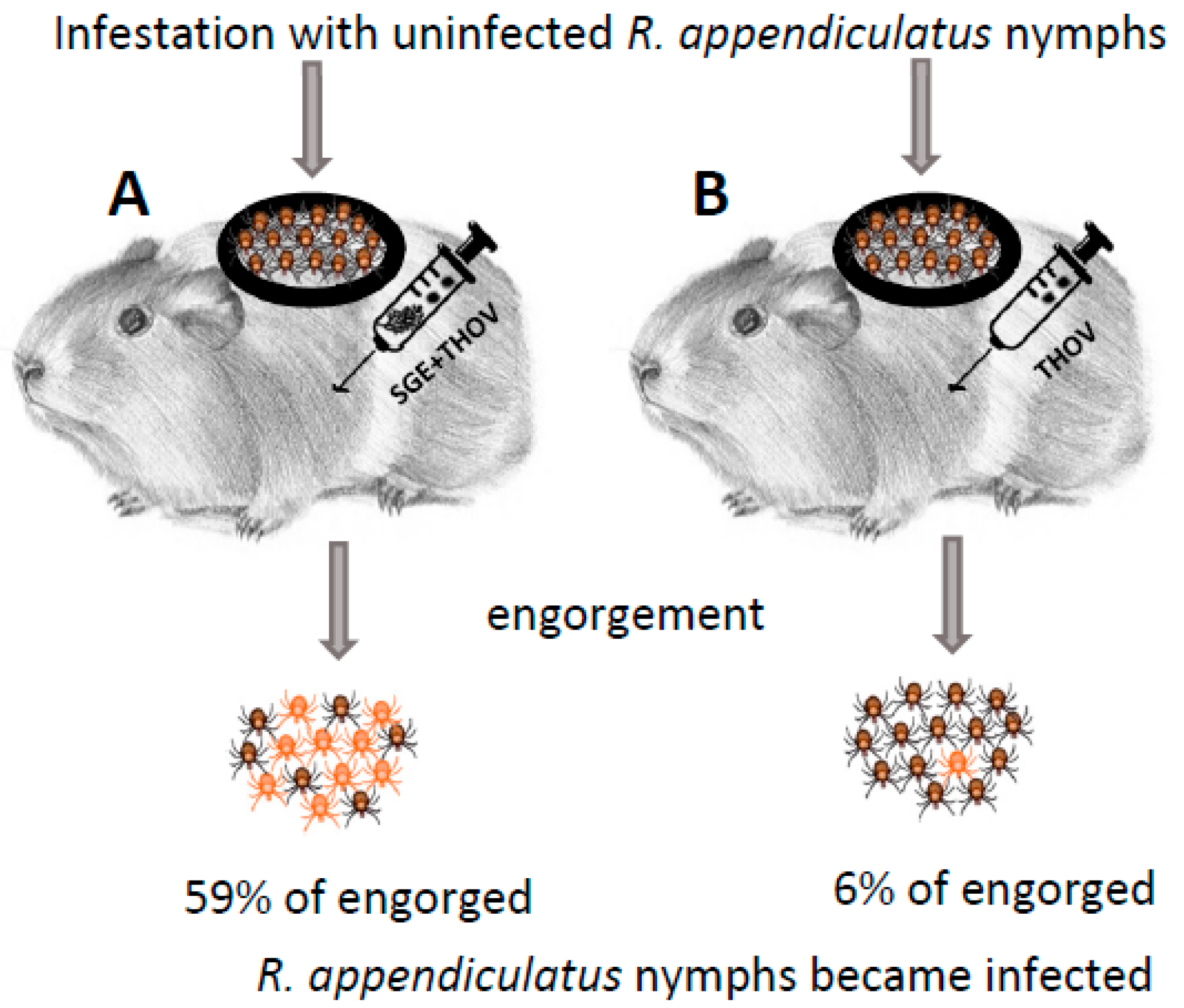
2. Discovery
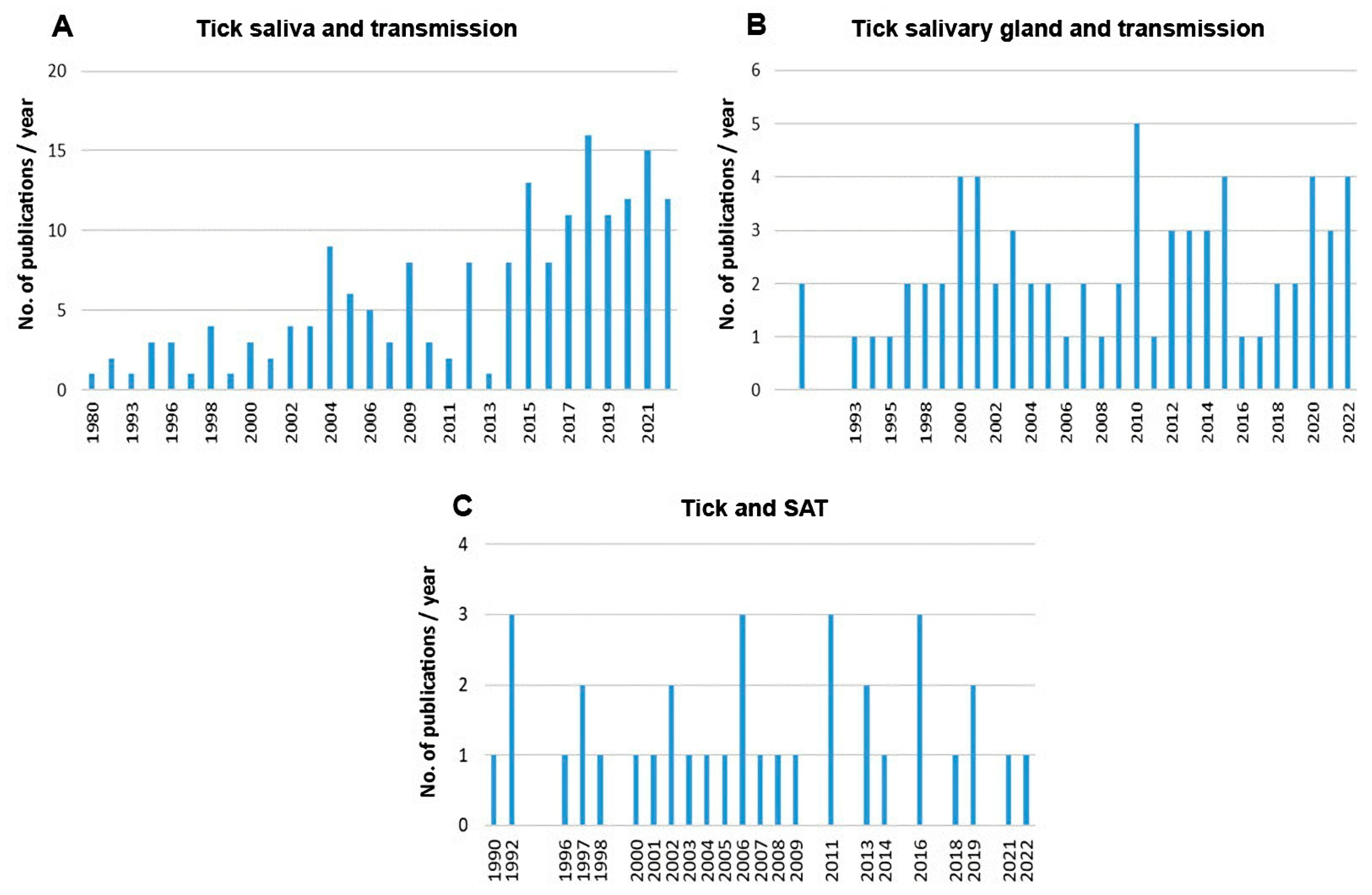
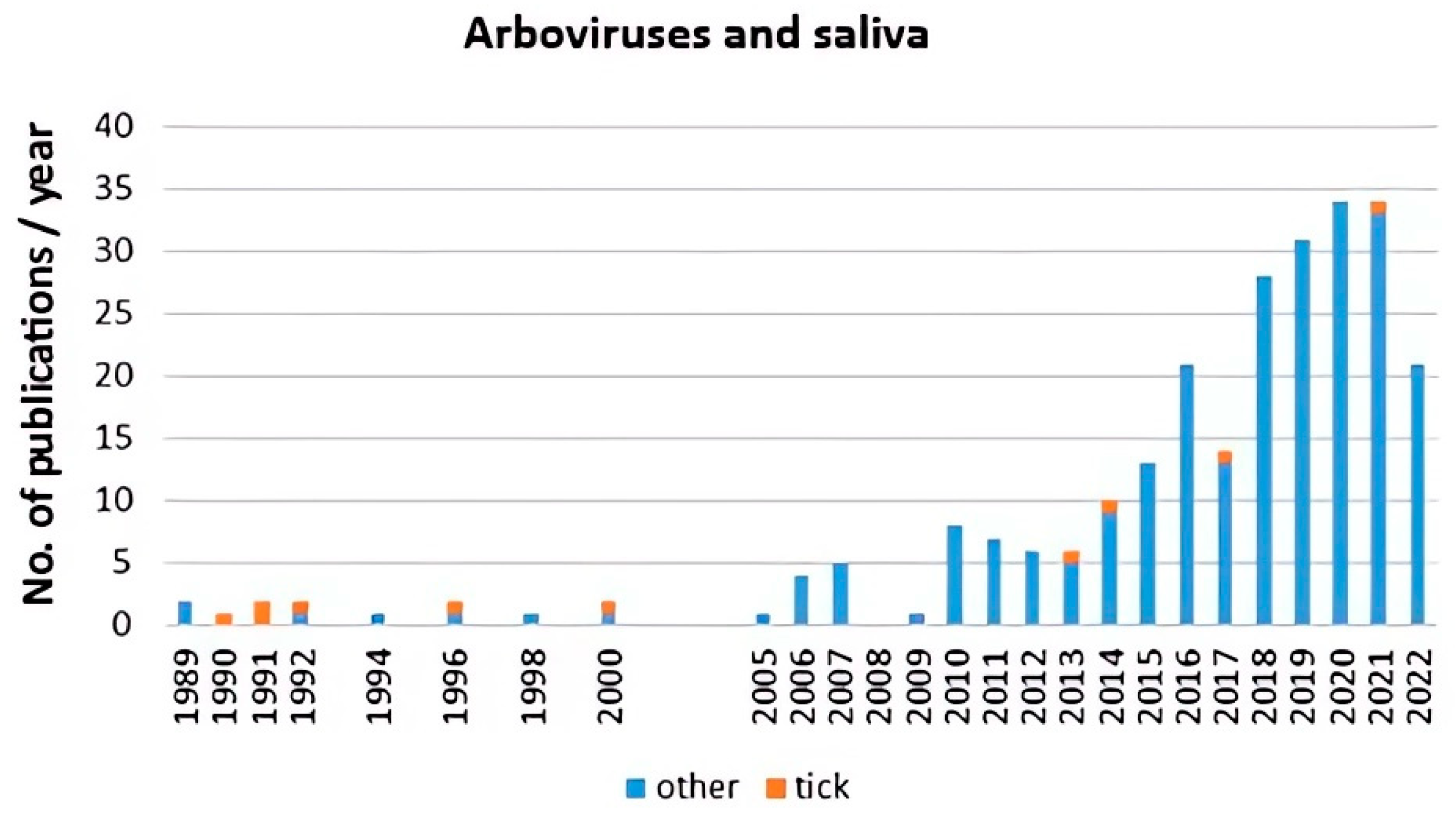
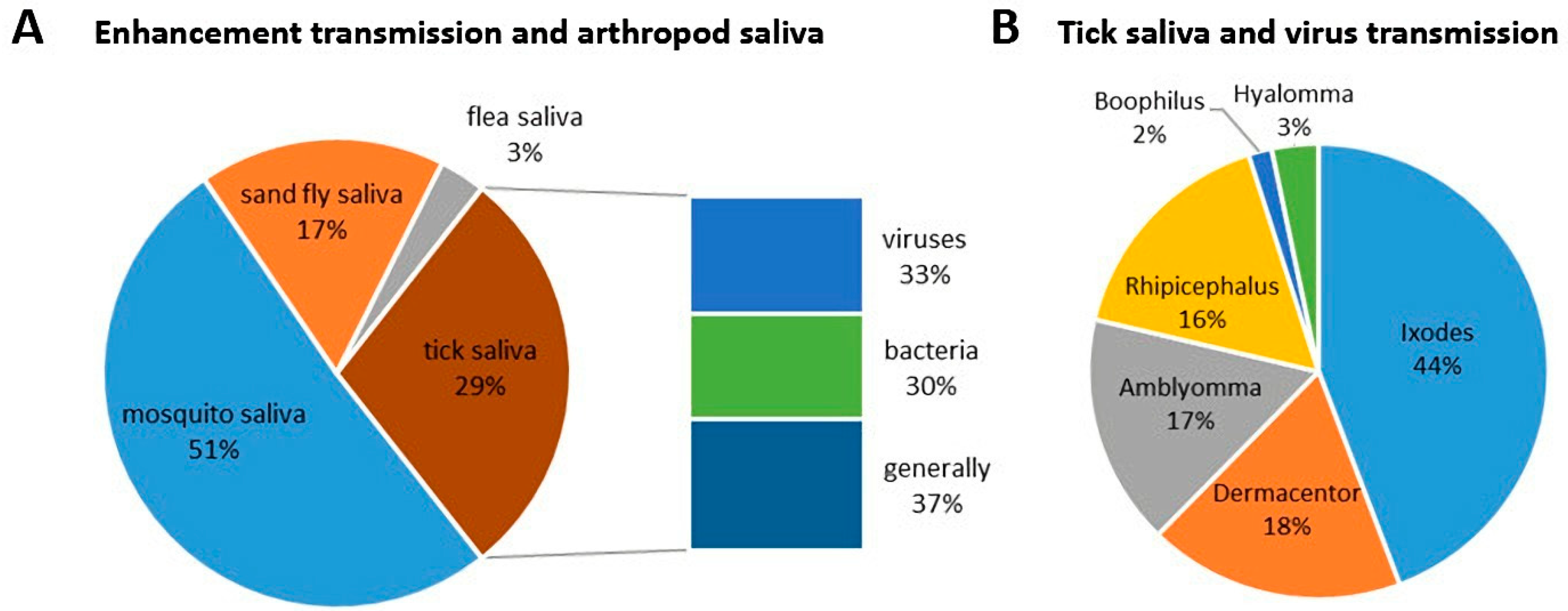
3. Impact

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Discovery of Non-Viraemic and Saliva-Assisted Transmission (1987, 1989) | ||
|---|---|---|
| Thogoto virus | Rhipicephalus appendiculatus | [8,10] |
| Amblyomma variegatum, A. hebraeum, A. cajennense, Rhipicephalus evertsi, R. microplus, Hyalomma dromedarii, H. marginatum | [11,30,66] | |
| TBEV | Ixodes persulcatus | [18,19] |
| I. ricinus, Dermacentor reticulatus, D. marginatus, R. appendiculatus | [14,15,16,17] | |
| Louping ill virus | I. ricinus | [38] |
| Powassan virus | I. scapularis | [41] |
| CCHFV | H. marginatum | [39] |
| Palma, Bhanja | R. appendiculatus, D. marginatus | [40] |
| Heartland virus | A. americanum | [42] |
| African Swine Fever virus | Ornithodoros porcinus | [43] |
| Discovery of the Effect of Host Resistance to Tick Infestation on Virus Transmission (1990) | ||
| Thogoto virus | R. appendiculatus | [28] |
| TBEV | I. ricinus | [16] |
| Description of the Tick Feeding Site as a Focus for Virus Replication in the Early Phase of Transmission (1996) | ||
| TBEV | I. ricinus | [36] |
| Modulation of Host Innate Responses by Tick Saliva or Salivary Gland Extracts (1994) and Enhancement of Virus Replication In Vitro (1998) | ||
| Natural killer cells | I. ricinus, D. reticulatus, A. variegatum, Haemaphysalis inermis, H. concinna | [45,46,47,48] |
| Interferon | I. ricinus, D. reticulatus | [44,47] |
| Dendritic cells | I. ricinus | [49,50] |
| Dendritic cells | I. scapularis salivary cystatin | [51] |
| Vesicular stomatitis virus | D. reticulatus | [33] |
| TBEV | I. ricinus | [49,50] |
| Introduction of High-Throughput and Systems Biology Approaches | ||
| A: Expression Profiles in Tick Salivary Glands During Feeding and Virus Infection (2012) | ||
| Langat virus | I. scapularis | [54] |
| Powassan virus | I. scapularis | [54,55] |
| TBEV | I. ricinus | [56] |
| B: Immunoprofiling of Host Cutaneous Responses to Tick Attachment and Virus Infection (2012) | ||
| Powassan virus | I. scapularis | [58,59,60] |
| TBEV | I. ricinus | [62] |
| Discovery of Extracellular Vesicles Serving as: | ||
| A: Vehicles for Transmission of Viral RNA (2018) | ||
| Langat virus | I. scapularis cell line | [63] |
| B: Modulators of Immune Responses in the Host Skin and Wound Healing (2020) | ||
| A. maculatum, I. scapularis | [64] | |
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kemp, D.H.; Stone, B.R.; Binnington, K.C. Chapter 4: Tick attachment and feeding: Role of the mouthparts, feeding apparatus, salivary gland secretions and host response. In Physiology of Ticks; Obenchain, F.D., Galun, R., Eds.; Pergamon Press: Oxford, UK, 1982; pp. 119–168. [Google Scholar]
- Ribeiro, J.M.; Makoul, G.T.; Levine, J.; Robinson, D.R.; Spielman, A. Antihemostatic, antiinflammatory, and immunosuppressive properties of the saliva of a tick Ixodes dammini. J. Exp. Med. 1985, 161, 332–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, J.M.C. Role of saliva in blood-feeding by arthropods. Annu. Rev. Entomol. 1987, 32, 463–478. [Google Scholar] [CrossRef] [PubMed]
- Šimo, L.; Kazimirova, M.; Richardson, J.; Bonnet, S.I. The essential role of tick salivary glands and saliva in tick feeding and pathogen transmission. Front. Cell. Infect. Microbiol. 2017, 7, 281. [Google Scholar] [CrossRef]
- Demarta-Gatsi, C.; Mécheri, S.; Paul, R.E. Arthropod saliva and its role in pathogen transmission: Insect saliva: Mosquito saliva, skin, allergy, and the outcome of malaria infection—From mice to men. In Skin and Arthropod Vectors, 1st ed.; Boulanger, N., Ed.; Academic Press Elsevier: London, UK, 2018; pp. 83–119. [Google Scholar] [CrossRef]
- Wikel, S.K. Tick-host-pathogen systems immunobiology: An interactive trio. Front. Biosci. 2018, 23, 265–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Titus, R.G.; Ribeiro, J.M.C. Salivary gland lysates from the sand fly Lutzomyia longipalpis enhance Leishmania infectivity. Science 1988, 239, 1306–1308. [Google Scholar] [CrossRef]
- Jones, L.D.; Davies, C.R.; Steele, G.M.; Nuttall, P.A. A novel mode of arbovirus transmission involving a nonviremic host. Science 1987, 237, 775–777. [Google Scholar] [CrossRef]
- Jones, L.D.; Nuttall, P.A. Non-viraemic transmission of Thogoto virus: Influence of time and distance. Trans. R. Soc. Trop. Med. Hyg. 1989, 83, 712–724. [Google Scholar] [CrossRef]
- Jones, L.D.; Hodgson, E.; Nuttall, P.A. Enhancement of virus transmission by tick salivary glands. J. Gen. Virol. 1989, 70, 1895–1898. [Google Scholar] [CrossRef]
- Jones, L.D.; Hodgson, E.; Williams, T.; Higgs, S.; Nuttall, P.A. Saliva activated transmission (SAT) of Thogoto virus: Relationship with vector potential of different haematophagous arthropods. Med. Vet. Entomol. 1992, 6, 261–265. [Google Scholar] [CrossRef]
- Nuttall, P.A.; Labuda, M. Tick-host interactions: Saliva-activated transmission. Parasitology 2004, 129, S177–S189. [Google Scholar] [CrossRef]
- Nuttall, P.A.; Labuda, M. Saliva-assisted transmission of tick borne pathogens. In Ticks: Biology, Disease and Control, 1st ed.; Bowman, A.S., Nuttall, P.A., Eds.; Cambridge University Press: Cambridge, UK, 2008; pp. 205–219. [Google Scholar]
- Labuda, M.; Jones, L.D.; Williams, T.; Nuttall, P.A. Enhancement of tick-borne encephalitis virus transmission by tick salivary gland extracts. Med. Vet. Entomol. 1993, 7, 193–196. [Google Scholar] [CrossRef]
- Labuda, M.; Jones, L.D.; Williams, T.; Danielova, V.; Nuttall, P.A. Efficient transmission of tick-borne encephalitis virus between cofeeding ticks. J. Med. Entomol. 1993, 30, 295–299. [Google Scholar] [CrossRef]
- Labuda, M.; Nuttall, P.A.; Kožuch, O.; Elečková, E.; Williams, T.; Žuffová, E.; Sabó, A. Non-viraemic transmission of tick-borne encephalitis virus: A mechanism for arbovirus survival in nature. Experientia 1993, 49, 802–805. [Google Scholar] [CrossRef]
- Labuda, M.; Danielova, V.; Jones, L.D.; Nuttall, P.A. Amplification of tick-borne encephalitis virus infection during co-feeding of ticks. Med. Vet. Entomol. 1993, 7, 339–342. [Google Scholar] [CrossRef] [PubMed]
- Alekseev, A.N.; Chunikhin, S.P.; Rukhkian, M.I.; Stefutkina, L.F. The possible role of the salivary gland substrate in ixodid ticks as an adjuvant enhancing arbovirus transmission. Med. Parazitol. 1991, 1, 28–31. [Google Scholar]
- Alekseev, A.N.; Chunikhin, S.P. Virus exchange between feeding ticks in the absence of viremia in a vertebrate host (distant transmission). Med. Parazitol. 1991, 2, 50–54. [Google Scholar]
- Nuttall, P.A. Tick saliva and its role in pathogen transmission. Wien. Klin. Wochenschr. 2019. [Google Scholar] [CrossRef] [Green Version]
- Pham, M.; Underwood, J.; Oliva Chávez, A.S. Changing the recipe: Pathogen directed changes in tick saliva components. Int. J. Environ. Res. Public Health 2021, 18, 1806. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Zeb, I.; Alouffi, A.; Zahid, H.; Almutairi, M.M.; Ayed Alshammari, F.; Alrouji, M.; Termignoni, C.; Vaz da Silva, I., Jr.; Tanaka, T. Host immune responses to salivary components—A critical facet of tick-host interactions. Front. Cell. Infect. Microbiol. 2022, 12, 809052. [Google Scholar] [CrossRef]
- Neelakanta, G.; Sultana, H. Tick saliva and salivary glands: What do we know so far on their role in arthropod blood feeding and pathogen transmission. Front. Cell. Infect. Microbiol. 2022, 11, 816547. [Google Scholar] [CrossRef]
- Kazimírová, M.; Thangamani, S.; Bartíková, P.; Hermance, M.; Holíková, V.; Štibrániová, I.; Nuttall, P.A. Tick-borne viruses and biological processes at the tick-host-virus interface. Front. Cell. Infect. Microbiol. 2017, 7, 339. [Google Scholar] [CrossRef] [Green Version]
- Hermance, M.E.; Thangamani, S. Tick-virus-host interactions at the cutaneous interface: The nidus of flavivirus transmission. Viruses 2018, 10, 362. [Google Scholar] [CrossRef] [Green Version]
- Maqbool, M.; Sajid, M.S.; Saqib, M.; Anjum, F.R.; Tayyab, M.H.; Rizwan, H.M.; Rashid, M.I.; Rashid, I.; Iqbal, A.; Siddique, R.M.; et al. Potential mechanisms of transmission of tick-borne viruses at the virus-tick interface. Front. Microbiol. 2022, 13, 846884. [Google Scholar] [CrossRef] [PubMed]
- Jones, L.D.; Hodgson, E.; Nuttall, P.A. Characterization of tick salivary gland factor(s) that enhance Thogoto virus transmission. Arch. Virol. 1990, 1, 227–234. [Google Scholar] [CrossRef]
- Jones, L.D.; Nuttall, P.A. The effect of host resistance to tick infestation on the transmission of Thogoto virus by ticks. J. Gen. Virol. 1990, 71, 1039–1043. [Google Scholar] [CrossRef] [PubMed]
- Jones, L.D.; Kaufman, W.R.; Nuttall, P.A. Modification of the skin feeding site by tick saliva mediates virus transmission. Experientia 1992, 48, 779–782. [Google Scholar] [CrossRef] [PubMed]
- Jones, L.D.; Matthewson, M.; Nuttall, P.A. Saliva-activated transmission (SAT) of Thogoto virus: Dynamics of SAT factor activity in the salivary glands of Rhipicephalus appendiculatus, Amblyomma variegatum, and Boophilus microplus ticks. Exp. Appl. Acarol. 1992, 13, 241–248. [Google Scholar] [CrossRef]
- Alekseev, A.N.; Chunikhin, S.P. Exchange of the tick-borne encephalitis virus between Ixodidae simultaneously feeding on the animals with subthreshold levels of viremia. Med. Parazitol. Parazit. Bolez. 1990, 2, 48–50. [Google Scholar]
- Alekseev, A.N.; Chunikhin, S.P. Difference in distant transmission ability of tick-borne encephalitis virus by ixodid ticks belonging to different subfamilies. Parazitologiya 1992, 26, 506–515. [Google Scholar]
- Hajnická, V.; Fuchsberger, N.; Slovák, M.; Kocáková, P.; Labuda, M.; Nuttall, P.A. Tick salivary gland extracts promote virus growth in vitro. Parasitology 1998, 116, 533–538. [Google Scholar] [CrossRef]
- Kocáková, P.; Hajnická, V.; Slovák, M.; Nuttall, P.A.; Fuchsberger, N. Promotion of vesicular stomatitis virus nucleocapsid protein production by arthopod saliva. Acta Virol. 1999, 43, 251–254. [Google Scholar] [PubMed]
- Sláviková, M.; Kocáková, P.; Slovák, M.; Vančová, I.; Hajnická, V.; Gašperík, J.; Fuchsberger, N.; Nuttall, P.A. Vesicular stomatitis virus nucleocapsid protein production in cells treated with selected fast protein liquid chromatography fractions of tick salivary gland extracts. Acta Virol. 2002, 46, 117–120. [Google Scholar] [PubMed]
- Labuda, M.; Austyn, J.M.; Zuffova, E.; Kozuch, O.; Fuchsberger, N.; Lysy, J.; Nuttall, P.A. Importance of localized skin infection in tick-borne encephalitis virus transmission. Virology 1996, 219, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Labuda, M.; Kozuch, O.; Zuffová, E.; Elecková, E.; Hails, R.S.; Nuttall, P.A. Tick-borne encephalitis virus transmission between ticks cofeeding on specific immune natural rodent hosts. Virology 1997, 235, 138–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, L.D.; Gaunt, M.; Hails, R.S.; Laurenson, K.; Hudson, P.J.; Reid, H.; Henbest, P.; Gould, E.A. Transmission of louping ill virus between infected and uninfected ticks co-feeding on mountain hares. Med. Vet. Entomol. 1997, 11, 172–176. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.W.; Linthicum, K.J.; Moulton, J.R. Transmission of Crimean-Congo hemorrhagic fever virus in two species of Hyalomma ticks from infected adults to cofeeding immature forms. Am. J. Trop. Med. Hyg. 1993, 48, 576–580. [Google Scholar] [CrossRef] [PubMed]
- Labuda, M.; Alves, M.J.; Elečková, E.; Kožuch, O.; Filipe, A.R. Transmission of tick-borne bunyaviruses by cofeeding ixodid ticks. Acta Virol. 1997, 41, 325–328. [Google Scholar]
- Hermance, M.E.; Thangamani, S. Tick saliva enhances Powassan virus transmission to the host, influencing its dissemination and the course of disease. J. Virol. 2015, 89, 7852–7860. [Google Scholar] [CrossRef] [Green Version]
- Godsey, M.S.; Savage, H.M.; Burkhalter, K.L.; Bosco-Lauth, A.M.; Delorey, M.J. Transmission of Heartland virus (Bunyaviridae: Phlebovirus) by experimentally infected Amblyomma americanum (Acari: Ixodidae). J. Med. Entomol. 2016, 53, 1226–1233. [Google Scholar] [CrossRef] [Green Version]
- Bernard, J.; Hutet, E.; Paboeuf, F.; Randriamparany, T.; Holzmuller, P.; Lancelot, R.; Rodrigues, V.; Vial, L.; Le Potier, M.F. Effect of O. porcinus tick salivary gland extract on the African swine fever virus infection in domestic pig. PLoS ONE 2016, 11, e0147869. [Google Scholar] [CrossRef] [Green Version]
- Hajnicka, V.; Kocakova, P.; Slovak, M.; Labuda, M.; Fuchsberger, N.; Nuttall, P.A. Inhibition of the antiviral action of interferon by tick salivary gland extract. Parasite Immunol. 2000, 22, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Kubeš, M.; Fuchsberger, N.; Labuda, M.; Žuffová, E.; Nuttall, P.A. Salivary-gland extracts of partially fed Dermacentor reticulatus ticks decrease natural-killer-cell activity in vitro. Immunology 1994, 82, 113–116. [Google Scholar] [PubMed]
- Kubeš, M.; Kocáková, P.; Slovák, M.; Sláviková, M.; Fuchsberger, N.; Nuttall, P.A. Heterogeneity in the effect of different ixodid tick species on human natural killer cell activity. Parasite Immunol. 2002, 24, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Kopecký, J.; Kuthejlová, M. Suppressive effect of Ixodes ricinus salivary gland extract on mechanisms of natural immunity in vitro. Parasite Immunol. 1998, 20, 169–174. [Google Scholar] [PubMed]
- Bartíková, P.; Slovák, M.; Štibrániová, I. Impact of tick salivary gland extracts on cytotoxic activity of mouse natural killer cells. Biologia 2022, 77, 1675–1683. [Google Scholar] [CrossRef]
- Fialová, A.; Cimburek, Z.; Iezzi, G.; Kopecký, J. Ixodes ricinus tick saliva modulates tick-borne encephalitis virus infection of dendritic cells. Microbes Infect. 2010, 12, 580–585. [Google Scholar] [CrossRef] [PubMed]
- Lieskovská, J.; Páleníková, J.; Langhansová, H.; Chmelař, J.; Kopecký, J. Saliva of Ixodes ricinus enhances TBE virus replication in dendritic cells by modulation of pro-survival Akt pathway. Virology 2018, 514, 98–105. [Google Scholar] [CrossRef]
- Lieskovská, J.; Páleníková, J.; Širmarová, J.; Elsterová, J.; Kotsyfakis, M.; Chagas, A.C.; Calvo, E.; Růžek, D.; Kopecký, J. Tick salivary cystatin sialostatin L2 suppresses IFN responses in mouse dendritic cells. Parasite Immunol. 2015, 37, 70–78. [Google Scholar] [CrossRef]
- Santos, R.I.; Hermance, M.E.; Reynolds, E.S.; Thangamani, S. Salivary gland extract from the deer tick, Ixodes scapularis, facilitates neuroinvasion by Powassan virus in BALB/c mice. Sci. Rep. 2021, 11, 20873. [Google Scholar] [CrossRef]
- Rodriguez, S.E.; McAuley, A.J.; Gargili, A.; Bente, D.A. Interactions of human dermal dendritic cells and Langerhans cells treated with Hyalomma tick saliva with Crimean-Congo hemorrhagic fever virus. Viruses 2018, 10, 381. [Google Scholar] [CrossRef] [Green Version]
- McNally, K.L.; Mitzel, D.N.; Anderson, J.M.; Ribeiro, J.M.C.; Valenzuela, J.G.; Myers, T.G.; Godinez, A.; Wolfinbarger, J.B.; Best, S.M.; Bloom, M.E. Differential salivary gland transcript expression profile in Ixodes scapularis nymphs upon feeding or flavivirus infection. Ticks Tick-Borne Dis. 2012, 3, 18–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermance, M.E.; Widen, S.G.; Wood, T.G.; Thangamani, S. Ixodes scapularis salivary gland microRNAs are differentially expressed during Powassan virus transmission. Sci. Rep. 2019, 9, 13110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hart, C.E.; Ribeiro, J.M.; Kazimirova, M.; Thangamani, S. Tick-borne encephalitis virus infection alters the sialome of Ixodes ricinus ticks during the earliest stages of feeding. Front. Cell. Infect. Microbiol. 2020, 10, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinze, D.M.; Carmical, J.R.; Aronson, J.F.; Thangamani, S. Early immunologic events at the tick-host interface. PLoS ONE 2012, 7, e47301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinze, D.M.; Wikel, S.K.; Thangamani, S.; Alarcon-Chaidez, F.J. Transcriptional profiling of the murine cutaneous response during initial and subsequent infestations with Ixodes scapularis nymphs. Parasites Vectors 2012, 5, 26. [Google Scholar] [CrossRef] [Green Version]
- Hermance, M.E.; Thangamani, S. Proinflammatory cytokines and chemokines at the skin interface during Powassan virus transmission. J. Investig. Dermatol. 2014, 134, 2280–2283. [Google Scholar] [CrossRef] [Green Version]
- Hermance, M.E.; Santos, R.I.; Kelly, B.C.; Valbuena, G.; Thangamani, S. Immune cell targets of infection at the tick-skin interface during Powassan virus transmission. PLoS ONE 2016, 11, e0155889. [Google Scholar] [CrossRef]
- Hermance, M.E.; Thangamani, S. Utilization of RNA in situ hybridization to understand the cellular localization of Powassan virus RNA at the tick-virus-host interface. Front. Cell. Infect. Microbiol. 2020, 10, 172. [Google Scholar] [CrossRef]
- Thangamani, S.; Hermance, M.E.; Santos, R.I.; Slovak, M.; Heinze, D.; Widen, S.G.; Kazimirova, M. Transcriptional im-munoprofiling at the tick-virus-host interface during early stages of tick-borne encephalitis virus transmission. Front. Cell. Infect. Microbiol. 2017, 7, 494. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Woodson, M.; Neupane, B.; Bai, F.; Sherman, M.B.; Choi, K.H.; Neelakanta, G.; Sultana, H. Exosomes serve as novel modes of tick-borne flavivirus transmission from arthropod to human cells and facilitates dissemination of viral RNA and proteins to the vertebrate neuronal cells. PLoS Pathog. 2018, 14, e1006764. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Tahir, F.; Wang, J.C.; Woodson, M.; Sherman, M.B.; Karim, S.; Neelakanta, G.; Sultana, H. Discovery of exosomes from tick saliva and salivary glands reveals therapeutic roles for CXCL12 and IL-8 in wound healing at the tick-human skin interface. Front. Cell. Dev. Biol. 2020, 8, 554. [Google Scholar] [CrossRef] [PubMed]
- Oliva Chávez, A.S.; Wang, X.; Marnin, L.; Archer, N.K.; Hammond, H.L.; Carroll, E.E.M.; Shaw, D.K.; Tully, B.G.; Buskirk, A.D.; Ford, S.L.; et al. Tick extracellular vesicles enable arthropod feeding and promote distinct outcomes of bacterial infection. Nat. Commun. 2021, 12, 3696. [Google Scholar] [CrossRef] [PubMed]
- Jones, L.D.; Davies, C.R.; Williams, T.; Cory, J.; Nuttall, P.A. Non-viraemic transmission of Thogoto virus: Vector efficiency of Rhipicephalus appendiculatus and Amblyomma variegatum. Trans. R. Soc. Trop. Med. Hyg. 1990, 84, 846–848. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bartíková, P.; Štibrániová, I.; Kazimírová, M. Discovery of the Role of Tick Salivary Glands in Enhancement of Virus Transmission—Beginning of an Exciting Story. Pathogens 2023, 12, 334. https://doi.org/10.3390/pathogens12020334
Bartíková P, Štibrániová I, Kazimírová M. Discovery of the Role of Tick Salivary Glands in Enhancement of Virus Transmission—Beginning of an Exciting Story. Pathogens. 2023; 12(2):334. https://doi.org/10.3390/pathogens12020334
Chicago/Turabian StyleBartíková, Pavlína, Iveta Štibrániová, and Mária Kazimírová. 2023. "Discovery of the Role of Tick Salivary Glands in Enhancement of Virus Transmission—Beginning of an Exciting Story" Pathogens 12, no. 2: 334. https://doi.org/10.3390/pathogens12020334