
Successful Removal of Angiostrongylus cantonensis Larvae from the Central Nervous System of Rats 7- and 14-Days Post-Infection Using a Product Containing Moxidectin, Sarolaner and Pyrantel Embonate (Simparica Trio™) in Experimental Infections


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Dosage Calculation
2.3. Examination for Patent Infection and Presence of A. cantonensis L1 in Rat Faeces
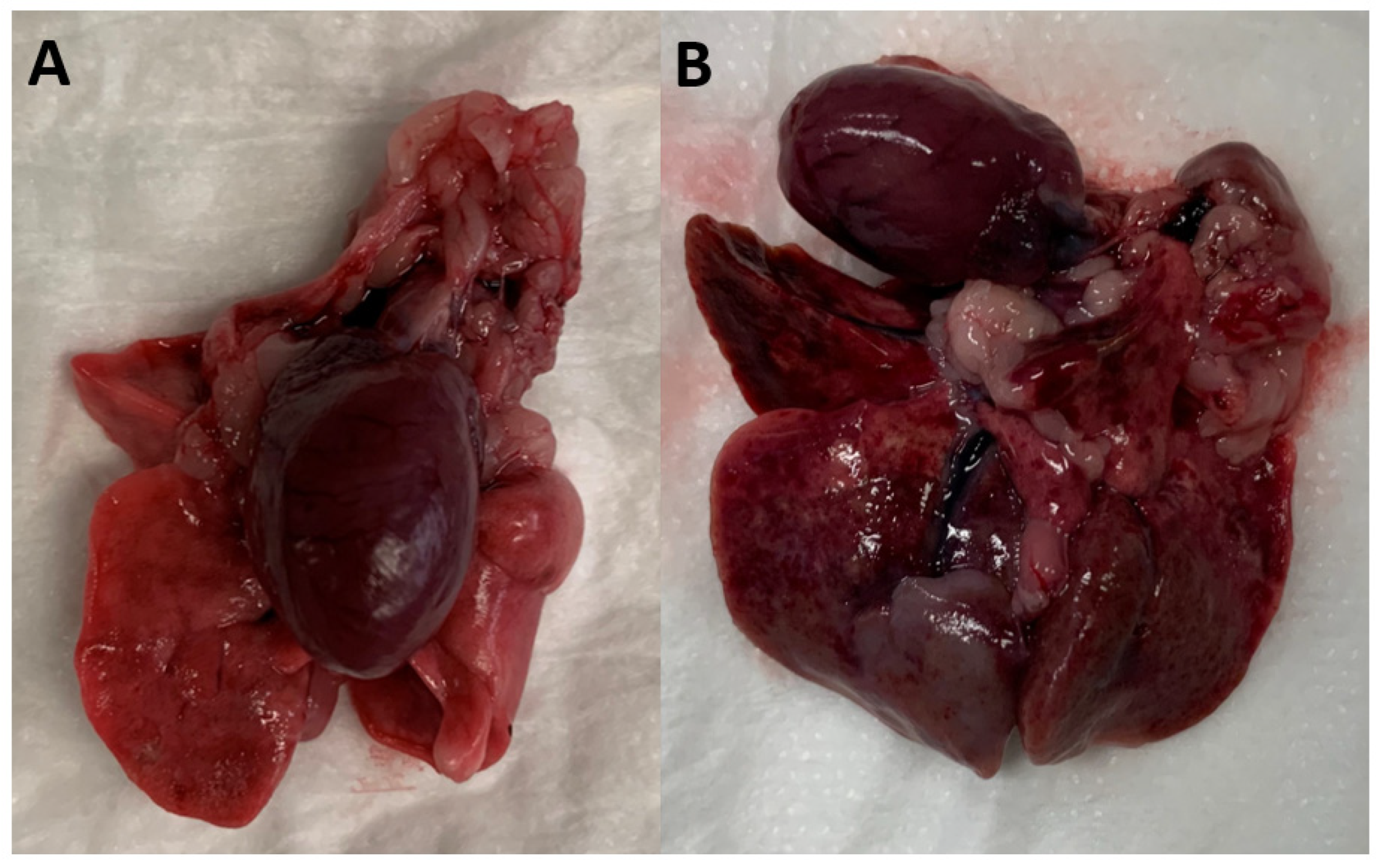
2.4. Necropsy Examination of Rats to Detect Adults A. cantonensis in the Right Ventricle and Pulmonary Artery of Rats
2.5. Statistical Analysis
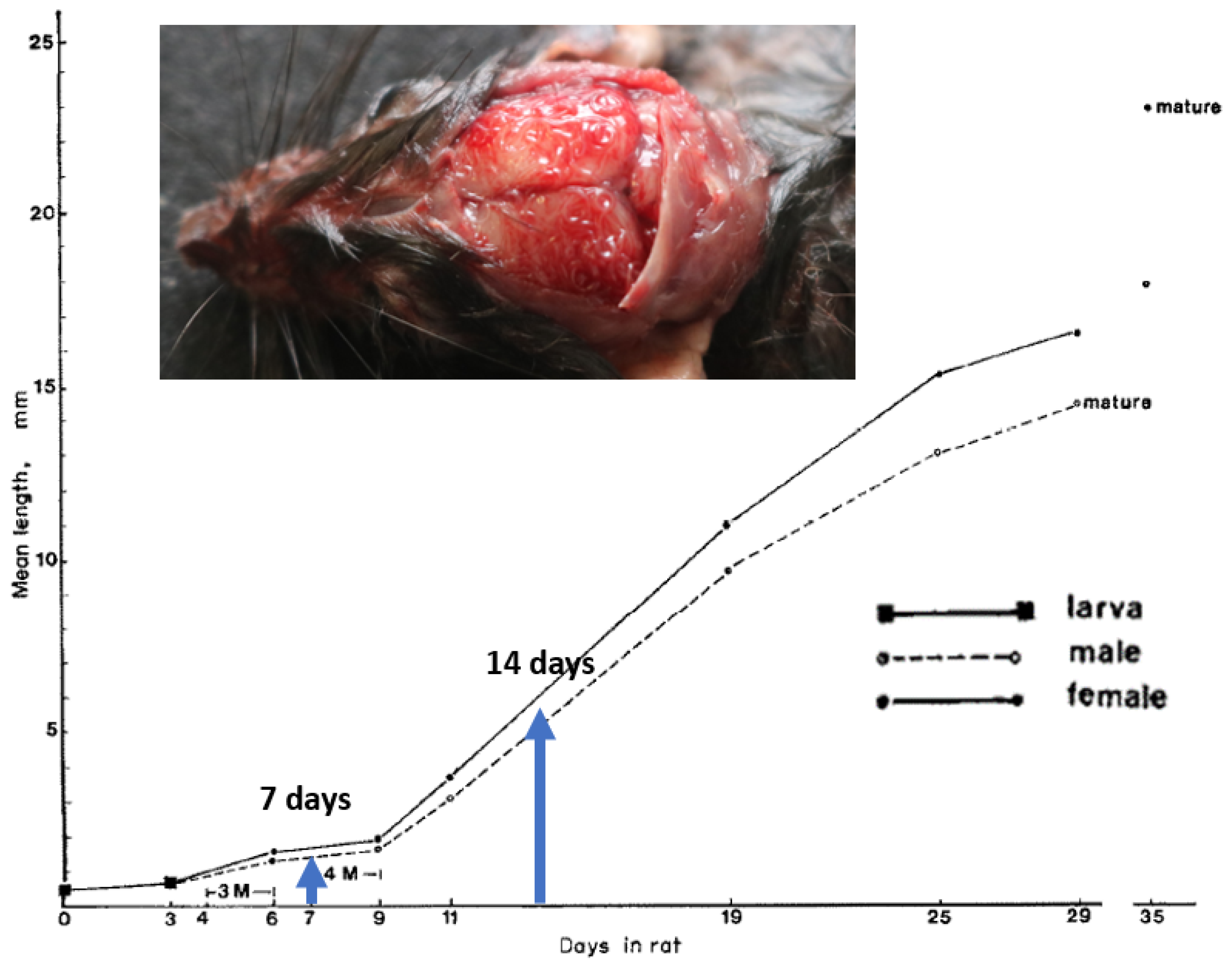
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Traub, R.; Brown, G.; Coleman, G.; Constantinoiu, C.; O’Handley, R.; Slapeta, J. Parasites of the dog and cat. 4.1.4. Parasites causing multi-systemic disease in small animals: Angiostrongylus spp. (Parastrongylus). In Australian Animal Parasites Inside & Out; Brown, G., Coleman, G., Constantinoiu, C., Gasser, R., Holyoake, P., Hobbs, R., Lymbery, A., O’Handley, R., Phalen, D., Pomroy, W., et al., Eds.; The Australian Society for Parasitology: Cairns North, Australia, 2015; pp. 546–549. [Google Scholar]
- Wun, M.K.; Davies, S.; Spielman, D.; Lee, R.; Hayward, D.; Malik, R. Gross, microscopic, radiologic, echocardiographic and haematological findings in rats experimentally infected with Angiostrongylus cantonensis. Parasitology 2020, 148, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Barratt, J.; Chan, D.; Sandaradura, I.; Malik, R.; Spielman, D.; Lee, R.; Marriott, D.; Harkness, J.; Ellis, J.; Stark, D. Angiostrongylus cantonensis: A review of its distribution, molecular biology and clinical significance as a human pathogen. Parasitology 2016, 143, 1087–1118. [Google Scholar] [CrossRef] [PubMed]
- Mackerras, M.; Sandars, D. The life history of the rat lung-worm, Angiostrongylus cantonensis (Chen) (Nematoda: Metastrongylidae). Aust. J. Zool. 1955, 3, 1–21. [Google Scholar] [CrossRef]
- Lee, R.; Pai, T.-Y.; Churcher, R.; Davies, S.; Braddock, J.; Linton, M.; Yu, J.; Bell, E.; Wimpole, J.; Dengate, A.; et al. Further studies of neuroangiostrongyliasis (rat lungworm disease) in Australian dogs: 92 new cases (2010–2020) and results for a novel, highly sensitive qPCR assay. Parasitology 2020, 148, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Mason, K.V. Canine neural angiostrongylosis: The clinical and therapeutic features of 55 natural cases. Aust. Veter. J. 1987, 64, 201–203. [Google Scholar] [CrossRef] [PubMed]
- Lunn, J.A.; Lee, R.; Smaller, J.; MacKay, B.M.; King, T.; Hunt, G.B.; Martin, P.; Krockenberger, M.B.; Spielman, D.; Malik, R. Twenty two cases of canine neural angiostrongylosis in eastern Australia (2002–2005) and a review of the literature. Parasites Vectors 2012, 5, 70. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.S.; Lúcio, C.D.S.; Bonfim, T.C.D.S.; Junior, A.M.; Tunholi, V.M.; Tunholi-Alves, V.M.; Mota, E.M.; Simões, R.D.O.; Santana, A.C.; Hooper, C.; et al. Metabolic and histopathological profile of Rattus norvegicus (Wistar) experimentally infected by Angiostrongylus cantonensis (Chen, 1935). Exp. Parasitol. 2014, 137, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Cowie, R.H. Biology, systematics, life cycle, and distribution of Angiostrongylus cantonensis, the cause of rat lungworm disease. Hawai’i J. Med. Public Health A J. Asia Pac. Med. Public Health 2013, 72 (Suppl. S2), 6–9. [Google Scholar]
- Willesen, J.; Kristensen, A.T.; Jensen, A.; Heine, J.; Koch, J. Efficacy and safety of imidacloprid/moxidectin spot-on solution and fenbendazole in the treatment of dogs naturally infected with Angiostrongylus vasorum (Baillet, 1866). Veter. Parasitol. 2007, 147, 258–264. [Google Scholar] [CrossRef]
- Becskei, C.; Willesen, J.L.; Schnyder, M.; Wozniakiewicz, M.; Miroshnikova, N.; Mahabir, S.P. Field safety and efficacy of an orally administered combination of sarolaner, moxidectin and pyrantel (Simparica Trio®) for the prevention of angiostrongylosis in dogs presented as veterinary patients. Parasites Vectors 2020, 13, 385. [Google Scholar] [CrossRef]
- Kryda, K.; Six, R.H.; Walsh, K.F.; Holzmer, S.J.; Chapin, S.; Mahabir, S.P.; Myers, M.; Inskeep, T.; Rugg, J.; Cundiff, B.; et al. Laboratory and field studies to investigate the efficacy of a novel, orally administered combination product containing moxidectin, sarolaner and pyrantel for the prevention of heartworm disease (Dirofilaria immitis) in dogs. Parasites Vectors 2019, 12, 445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Cesare, A.; Traversa, D. Canine angiostrongylosis: Recent advances in diagnosis, prevention, and treatment. Vet. Med. 2014, 5, 181–192. [Google Scholar] [CrossRef]
- The European Agency for the Evaluation of Medical Products; Committee for Veterinary Medicinal Products. Moxidectin. 1997. Available online: https://www.ema.europa.eu/en/documents/mrl-report/moxidectin-summary-report-1-committee-veterinary-medicinal-products_en.pdf (accessed on 20 May 2021).
- Bowman, D.D.; Charles, S.D.; Arther, R.G.; Settje, T. Laboratory Evaluation of the Efficacy of 10% Imidacloprid + 2.5% Moxidectin Topical Solution (Advantage® Multi, Advocate®) for the Treatment of Dirofilaria immitis Circulating Microfilariae in Dogs. Parasitol. Res. 2015, 114 Suppl. S1, S165–S174. [Google Scholar] [CrossRef] [PubMed]
- Simparica Trio Zoetis Petcare. Available online: https://www.zoetispetcare.com/products/simparica-trio (accessed on 20 May 2021).
- Snyder, P.W.; Hogg, J.T.; Ezenwa, V.O. Comparison of modified Flotac and Baermann techniques for quantifying lungworm larvae in free-ranging bighorn sheep (Ovis canadensis) feces, Montana, USA. J. Wildl. Dis. 2015, 51, 843–848. [Google Scholar] [CrossRef] [PubMed]
- Schmahl, G.; Mehlhorn, H.; Harder, A.; Klimpel, S.; Krieger, K.J. Efficacy of a Combination of Imidacloprid plus Moxidectin against Larval and Adult Stages of Nematodes (Trichuris muris, Angiostrongylus cantonensis) in Rodents. Parasitol. Res. 2007, 101 (Suppl. S1), 85–92. [Google Scholar] [CrossRef]
- Hess, A.S.; Hess, J.R. Understanding tests of the association of categorical variables: The Pearson chi-square test and Fisher’s exact test. Transfusion 2017, 57, 877–879. [Google Scholar] [CrossRef] [PubMed]
- Neely, J.G.; Hartman, J.M.; Forsen, J.W.; Wallace, M.S.; Clinical Research Working Group. Tutorials in Clinical Research: Part VII. Understanding Comparative Statistics (Contrast)—Part A: General Concepts of Statistical Significance. Laryngoscope 2003, 113, 1534–1540. [Google Scholar] [CrossRef] [PubMed]
- Bhaibulaya, M. Comparative studies on the life history of Angiostrongylus mackerrasae (Bhaibulaya, 1968) and Angiostrongylus cantonensis (Chen, 1935). Int. J. Parasitol. 1975, 5, 7–20. [Google Scholar] [CrossRef] [PubMed]
- Buchter, V.; Hofmann, D.; Häberli, C.; Keiser, J. Characterization of Moxidectin against Strongyloides ratti: In Vitro and In Vivo Activity and Pharmacokinetics in the Rat Model. ACS Infect. Dis. 2020, 7, 1069–1076. [Google Scholar] [CrossRef] [PubMed]
- Janko, C.; Geyer, J. Moxidectin has a lower neurotoxic potential but comparable brain penetration in P-glycoprotein-deficient CF-1 mice compared to ivermectin. J. Veter. Pharmacol. Ther. 2012, 36, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Nau, R.; Sörgel, F.; Eiffert, H. Penetration of Drugs through the Blood-Cerebrospinal Fluid/Blood-Brain Barrier for Treatment of Central Nervous System Infections. Clin. Microbiol. Rev. 2010, 23, 858–883. [Google Scholar] [CrossRef] [Green Version]
- Damsch, S.; Eichenbaum, G.; Tonelli, A.; Lammens, L.; Bulck, K.V.D.; Feyen, B.; Vandenberghe, J.; Megens, A.; Knight, E.; Kelley, M. Gavage-Related Reflux in Rats: Identification, pathogenesis, and toxicological implications (review). Toxicol. Pathol. 2010, 39, 348–360. [Google Scholar] [CrossRef] [PubMed]
- Helm, J.R.; Morgan, E.R.; Jackson, M.W.; Wotton, P.; Bell, R. Canine angiostrongylosis: An emerging disease in Europe. J. Veter. Emerg. Crit. Care 2010, 20, 98–109. [Google Scholar] [CrossRef] [PubMed]
- Page, S.W. Chapter 10. Antiparasitic Drugs. In Small Animal Clinical Pharmacology; Maddison, J.E., Page, S.W., Church, D.B., Eds.; Saunders/Elsevier: Edinburgh, UK, 2008; pp. 210–214. ISBN 9780702028588. [Google Scholar]



{kind=link}
{kind=link}
| Week | Group 1 (Control; n = 4) | Group 2 (Control; n = 4) | Group 3 (ST; n = 5) | Group 4 (ST; n = 5) | |
|---|---|---|---|---|---|
| 0 | M-S-P | M-S-P | |||
| 1 | |||||
| 2 | 30 L3 larvae PO | 30 L3 larvae PO | 30 L3 larvae PO |  | |
| 3 | |||||
| 4 | M-S-P | M-S-P | |||
| 5 | |||||
| 6 | |||||
| 7 | 30 L3 larvae PO | 30 L3 larvae PO |  | ||
| 8 | M-S-P | M-S-P | |||
| 9 | |||||
| 10 | |||||
| 11 | |||||
| 12 | |||||
| 13 | |||||
| 14 | Necropsy | Necropsy | Necropsy | Necropsy |
| Parasite Status | Group 1 (Control; n =4) One Challenge at 2 Weeks | Group 2 (Control; n = 4) One Challenge at 7 Weeks | Group 3 (M-S-P; n = 5) One Challenge at 2 Weeks | Group 4 (M-S-P; n = 5) Two Challenges at 2 and 7 Weeks |
|---|---|---|---|---|
| Infected | 4/4 | 4/4 | 0/5 | 2/5 |
| A.c.Male | 4,5,10,1 | 17,8,10,6 | 0,0,0,0,0 | 0,0,0,0,0 |
| A.c.Female | 8,6,6,0 | 6,9,6,7 | 0,0,0,0,0 | 1,0,0,0,1 |
| A.c.Total | 12,11,16,1 | 23,17,16,13 | 0,0,0,0,0 | 1,0,0,0,1 |
| L1 larvae in fresh faeces | 3/4 rats positive (1 rat with a single male worm was negative) | 4/4 positive | 0/5 positive | 0/5 positive (2/5 rats each had a single female worm and were negative) |
| Lung lesions | 3/4 | 4/4 | 0/5 | 0/5 |
| Body weight (g) | 690,674,662,595 (median 668) | 580,673,595,591 (median 593) | 646,586,595,616,538 (median 595) | 584,565,633,616,589 (median 589) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Henry, M.; Malik, R.; Šlapeta, J.; Lee, R. Successful Removal of Angiostrongylus cantonensis Larvae from the Central Nervous System of Rats 7- and 14-Days Post-Infection Using a Product Containing Moxidectin, Sarolaner and Pyrantel Embonate (Simparica Trio™) in Experimental Infections. Pathogens 2023, 12, 305. https://doi.org/10.3390/pathogens12020305
Henry M, Malik R, Šlapeta J, Lee R. Successful Removal of Angiostrongylus cantonensis Larvae from the Central Nervous System of Rats 7- and 14-Days Post-Infection Using a Product Containing Moxidectin, Sarolaner and Pyrantel Embonate (Simparica Trio™) in Experimental Infections. Pathogens. 2023; 12(2):305. https://doi.org/10.3390/pathogens12020305
Chicago/Turabian StyleHenry, Michaela, Richard Malik, Jan Šlapeta, and Rogan Lee. 2023. "Successful Removal of Angiostrongylus cantonensis Larvae from the Central Nervous System of Rats 7- and 14-Days Post-Infection Using a Product Containing Moxidectin, Sarolaner and Pyrantel Embonate (Simparica Trio™) in Experimental Infections" Pathogens 12, no. 2: 305. https://doi.org/10.3390/pathogens12020305