

Optimization and Application of Real-Time qPCR Assays in Detection and Identification of Chlamydiales in Products of Domestic Ruminant Abortion

Abstract
:1. Introduction
2. Materials and Methods
2.1. Synthetic Controls
2.2. Microorganisms
2.3. Clinical Samples
2.4. DNA Extraction
2.5. Primers and Probes for Detection of Chlamydiales

{kind=link}
{kind=link}
{kind=link}
| Chlamydiales | Target Gene | Primer and Probe Sequences (5′ to 3′) | Base Pair Sizes | Concentration (μM) | References |
|---|---|---|---|---|---|
| Singleplex qPCR assay | |||||
| Pan-Chlamydiales | 16S rRNA | For CCGCCAACACTGGGACT | 207 to 215 | 0.1 | [32] |
| Rev GGAGTTAGCCGGTGCTTCTTTAC | 0.1 | ||||
| P VIC-CTACGGGAGGCTGCAGTCGAGAATC-MGB | 0.1 | ||||
| Multiplex qPCR assays | |||||
| Assay 1 Chlamydia abortus | ompA | For GCAACTGACACTAAGTCGGCTACA | 82 | 0.9 | [34] |
| Rev ACAAGCATGTTCAATCGATAAGAGA | 0.9 | ||||
| P FAM-TAAATACCACGAATGGCAAGTTGGTTTAGCG-TAMRA | 0.2 | ||||
| Chlamydia pecorum | ompA | For CCATGTGATCCTTGCGCTACT | 76 | 0.9 | [35] |
| Rev TGTCGAAAACATAATCTCCGTAAAAT | 0.9 | ||||
| P VIC-TGCGACGCGATTAGCTTACGCGTAG-TAMRA | 0.2 | ||||
| Assay 2 Waddlia chondrophila | 16SrRNA | For GGCCCTTGGGTCGTAAAGTTCT | 101 | 0.5 (This study) | [27] |
| Rev CGGAGTTAGCCGGTGCTTCT | 0.5 | ||||
| P VIC-CATGGGAACAAGAGAAGGATG-MGB | 0.2 | ||||
| Parachlamydia acanthamoebae | 16SrRNA | For CTCAACTCCAGAACAGCATTT | 103 | 0.5 (This study) | [36] |
| Rev CTCAGCGTCAGGAATAAGC | 0.5 | ||||
| P FAM-TTCCACATGTAGCGGTGAAATGCGTAGATATG-MGB | 0.2 |
2.6. Quantitative Real-Time PCR (qPCR)
2.7. Analytical Sensitivity
2.8. Analytical Specificity
2.9. Limit of Detection of the qPCR Assays
2.10. Diagnostic Sensitivity
2.11. Diagnostic Specificity
2.12. Quality Assurance
2.13. Data Analysis
3. Results
3.1. Sensitivity, Specificity, Efficiency, and Limit of Detection of qPCR Assays for Detection of Chlamydiales
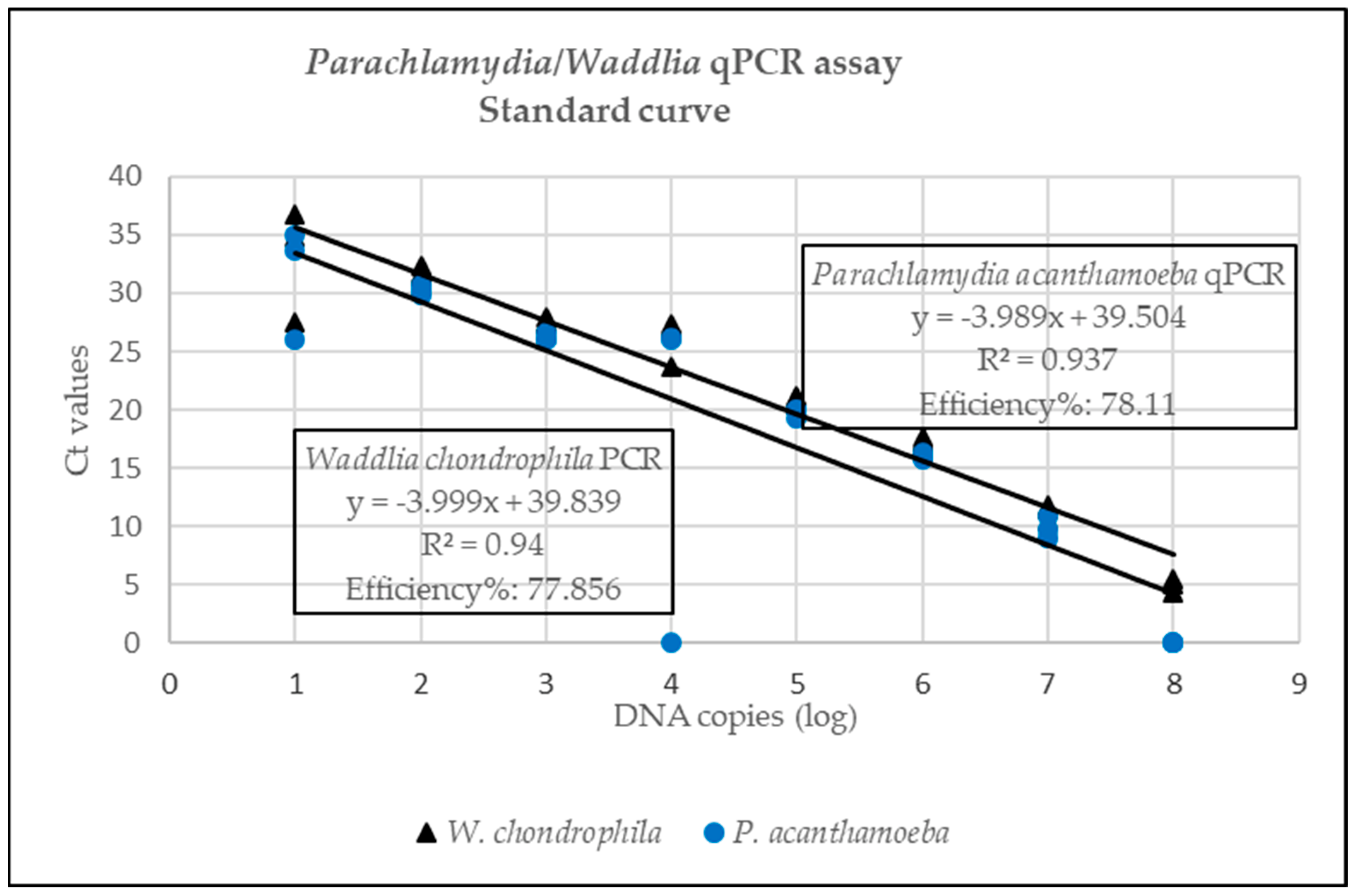
3.1.1. Analytical Sensitivity
3.1.2. Limit of Detection
3.1.3. Analytical Specificity
3.2. Clinical Samples
3.2.1. Diagnostic Sensitivity
3.2.2. Diagnostic Specificity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sebastiani, C.; Curcio, L.; Ciullo, M.; Cruciani, D.; Crotti, S.; Pesca, C.; Torricelli, M.; Sebastianelli, M.; Felici, A.; Biagetti, M. A multi-screening Fast qPCR approach to the identification of abortive agents in ruminants. J. Microbiol. Meth. 2018, 148, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Clothier, K.; Anderson, M. Evaluation of bovine abortion cases and tissue suitability for identification of infectious agents in California diagnostic laboratory cases from 2007 to 2012. Theriogenology 2016, 85, 933–938. [Google Scholar] [CrossRef] [PubMed]
- Baumgartner, W. Fetal Disease and abortion: Diagnosis and causes. In Bovine Reproduction, 1st ed.; Hopper, R.M., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2015; pp. 481–517. [Google Scholar] [CrossRef]
- Borel, N.; Polkinghorne, A.; Pospischil, A. A review on chlamydial diseases in animals: Still a challenge for pathologists? Vet. Pathol. 2018, 55, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Ammerdorffer, A.; Stojanov, M.; Greub, G.; Baud, D. Chlamydia trachomatis and Chlamydia-like bacteria: New enemies of human pregnancies. Curr. Opin. Infect. Dis. 2017, 30, 289–296. [Google Scholar] [CrossRef]
- Pichon, N.; Guindre, L.; Laroucau, K.; Cantaloube, M.; Nallatamby, A.; Parreau, S. Chlamydia abortus in pregnant woman with acute respiratory distress syndrome. Emerg. Infect. Dis. 2020, 26, 628–629. [Google Scholar] [CrossRef]
- Greub, G.; Raoult, D. Parachlamydiaceae: Potential Emerging Pathogens. Emerg. Infect. Dis. 2002, 8, 626–630. [Google Scholar] [CrossRef]
- Essig, A.; Longbottom, D. Chlamydia abortus: New aspects of infectious abortions in sheep and potential risk for pregnant women. Curr. Clin. Microbiol. Rep. 2015, 2, 22–34. [Google Scholar] [CrossRef]
- Walker, E.; Lee, E.J.; Timms, P.; Polkinghorne, A. Chlamydia pecorum infections in sheep and cattle: A common and under recognised infectious disease with significant impact on animal health. Vet. J. 2015, 206, 252–260. [Google Scholar] [CrossRef]
- Borel, N.; Frey, C.F.; Gottstein, B.; Hilbe, M.; Pospischil, A.; Franzoso, F.D.; Waldvogel, A. Laboratory diagnosis of ruminant abortion in Europe. Vet. J. 2014, 200, 218–229. [Google Scholar] [CrossRef] [Green Version]
- Markey, B.; Leonard, F.; Archambault, M.; Cullinane, A.; Maguire, D. Clinical Veterinary Microbiology, 2nd ed.; Mosby Elsevier: Beijing, China, 2013. [Google Scholar]
- Givens, M.D.; Marley, M.S.D. Infectious causes of embryonic and fetal mortality. Theriogenology 2008, 70, 270–285. [Google Scholar] [CrossRef]
- Gianitti, F.; Anderson, M.; Miller, M.; Rowe, J.; Sverlow, K.; Vasquez, M.; Cantón, G. Chlamydia pecorum: Fatal and placental lesions in sporadic caprine abortion. J. Vet. Diagn. Investig. 2015, 28, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Sachse, K.; Vretou, E.; Livingstone, M.; Borel, N.; Pospischil, A.; Longbottom, D. Recent developments in the laboratory diagnosis of chlamydial infections. Vet. Microbiol. 2009, 135, 2–21. [Google Scholar] [CrossRef]
- Clune, T.; Besier, S.; Hair, S.; Hancock, S.; Lockwood, A.; Thompson, A.; Jelocnik, M.; Jacobson, C. Chlamydia pecorum detection in aborted and stillborn lambs from Western Australia. BMC Vet. Res. 2021, 52, 84. [Google Scholar] [CrossRef] [PubMed]
- Henning, K.; Schares, G.; Granzow, H.; Polster, U.; Hartmann, M.; Hotzel, H.; Sachse, K.; Peters, M.; Rauser, M. Neospora caninum and Waddlia chondrophila strain 2032/99 in a septic stillborn calf. Vet. Microbiol. 2002, 85, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Barkallah, M.; Gharbi, Y.; Ben Hassena, A.; Ben Silma, A.; Mallek, Z.; Gautier, M.; Greub, G.; Gdoura, R.; Fendri, I. Survey of infectious etiologies of bovine abortion during mid-to late gestation in dairy herds. PLoS ONE 2014, 9, e91549. [Google Scholar] [CrossRef]
- Ruhl, S.; Casson, N.; Kaiser, C.; Thoma, R.; Pospischil, A.; Greub, G.; Borel, N. Evidence for Parachlamydia in bovine abortion. Vet. Microbiol. 2009, 135, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Barkallah, M.; Jribi, H.; Ben Silma, A.; Gharbi, Y.; Mallek, Z.; Gautier, M.; Fendri, I.; Gdoura, R. Molecular prevalence of Chlamydia and Chlamydia-like bacteria in Tunisian domestic ruminant farms and their influencing risk factors. Transbound. Emerg. Dis. 2018, 65, e329–e338. [Google Scholar] [CrossRef] [PubMed]
- Barkallah, M.; Gharbi, Y.; Ben Silma, A.; Elleuch, F.; Mallek, Z.; Ben Saad, R.; Gautier, M.; Gdoura, R.; Fendri, I. Simultaneous detection of Waddlia chondrophila and Listeria monocytogenes in aborted ruminant samples by real-time quantitative PCR. J. Microbiol. Methods 2016, 125, 64–69. [Google Scholar] [CrossRef]
- Hazlett, M.J.; McDowall, R.; DeLay, J.; Stalker, M.; McEwen, B.; van Dreumel, T.; Spinato, M.; Binnington, B.; Slavic, D.; Carman, S.; et al. A prospective study of sheep and goat abortion using real-time polymerase chain reaction and cut point estimation shows Coxiella burnetii and Chlamydophila abortus infection concurrently with other major pathogens. J. Vet. Diagn. Investig. 2013, 25, 359–368. [Google Scholar] [CrossRef] [Green Version]
- Blumer, S.; Greub, G.; Waldvogel, A.; Hässig, M.; Thoma, R.; Tschuor, A.; Pospischil, A.; Borel, N. Waddlia, Parachlamydia and Chlamydiaceae in bovine abortion. Vet. Microbiol. 2011, 152, 385–393. [Google Scholar] [CrossRef]
- Borel, N.; Ruhl, S.; Casson, N.; Kaiser, C.; Pospischil, A.; Greub, G. Parachlamydia spp. and related Chlamydia-like organisms and bovine abortion. In Emerg. Infect. Dis.; 2007; 13, p. 12. Available online: http://www.cdc.gov/EID/content/13/12/1904.htm (accessed on 6 February 2022). [CrossRef]
- Ruhl, S.; Goy, G.; Casson, N.; Thoma, R.; Pospischil, A.; Greub, G.; Borel, N. Parachlamydia acanthamoeba infection and abortion in small ruminants. Emerg. Infect. Dis. 2008, 14, 1966–1968. [Google Scholar] [CrossRef] [PubMed]
- Vidal, S.; Kegler, K.; Greub, G.; Aeby, S.; Borel, N.; Dagleish, M.P.; Posthaus, H.; Perreten, V.; Rodriguez-Campos, S. Neglected zoonotic agents in cattle abortion: Tackling the difficult to grow bacteria. BMC Vet. Res. 2017, 13, 373. [Google Scholar] [CrossRef] [PubMed]
- Rodolakis, A.; Laroucau, K. Chlamydiaceae and chlamydial infections in sheep or goats. Vet. Microbiol. 2015, 181, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Goy, G.; Croxatto, A.; Posfay-Barbe, K.M.; Gervaix, A.; Greub, G. Development of a real-time PCR for the specific detection of Waddlia chondrophila in clinical samples. Eur. J. Clin. Microbiol. Infect. Dis. 2009, 28, 1483–1486. [Google Scholar] [CrossRef]
- Kirkbride, C. Examination of Bovine and Ovine fetuses. Vet. Clin. N. Am. Food Anim. 1986, 2, 61–83. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Ren, H.; Zhou, H.; Quin, T.; Chen, Y. Simultaneous detection of nine key bacterial respiratory pathogens using Luminex xTAG® Technology. Int. J. Env. Res. Public Health 2017, 14, 223. [Google Scholar] [CrossRef]
- Iraola, G.; Pérez, R.; Betancor, L.; Marandino, A.; Morsella, C.; Méndez, A.; Paolicchi, F.; Piccirillo, A.; Tomás, G.; Velilla, A.; et al. A novel real-time PCR assay for quantitative detection of Campylobacter fetus based on ribosomal sequences. BMC Vet. Res. 2016, 12, 286. [Google Scholar] [CrossRef]
- Schnydrig, P.; Vidal, S.; Brodard, I.; Frey, C.F.; Posthaus, H.; Perreten, V.; Rodrigues-Campos, S. Bacterial, fungal, parasitological, and pathological analyses of abortions in small ruminants from 2012–2016. SAT/ASMV 2017, 12, 647–656. [Google Scholar] [CrossRef]
- Lienard, J.; Croxatto, A.; Aeby, S.; Jaton, K.; Posfay-Barbe, K.; Gervaix, A.; Greub, G. Development of a new Chlamydiales-specific real-time PCR and its application to respiratory clinical samples. J. Clin. Microbiol. 2011, 49, 2637–2642. [Google Scholar] [CrossRef]
- Wheelhouse, N.; Longbottom, D.; O’Donovan, J. Chlamydia in cases of bovine abortion in Ireland. Vet. Rec. 2014, 174, 560–561. [Google Scholar] [CrossRef] [PubMed]
- Pantchev, A.; Sting, R.; Bauerfeind, R.; Tyczka, J.; Sachse, K. New real-time PCR tests for species-specific detection of Chlamydophila psittaci and Chlamydophila abortus from tissue samples. Vet. J. 2009, 181, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Pantchev, A.; Sting, R.; Bauerfeind, R.; Tyczka, J.; Sachse, K. Detection of all Chlamydophila and Chlamydia spp. of veterinary interest using species-specific real-time PCR assays. Comp. Immunol. Microbiol. Infect. Dis. 2010, 33, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Casson, N.; Posfay-Barbe, K.M.; Gervaix, A.; Greub, G. New diagnostic real-time PCR for specific detection of Parachlamydia acanthamoebae DNA in clinical samples. J. Clin. Microbiol. 2008, 46, 1491–1493. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, J.; O’Donovan, J.; Proctor, A.; Brady, C.; Marques, P.X.; Worral, S.; Nally, J.E.; McElroy, M.; Basset, H.; Fagan, J.; et al. Application of quantitative real-time polymerase chain reaction for the diagnosis of toxoplasmosis and enzootic abortion of ewes. J. Vet. Diagn. Inv. 2012, 24, 846–854. [Google Scholar] [CrossRef] [PubMed]
- Berri, M.; Rekiki, A.; Sidi Boumedine, K.; Rodolakis, A. Simultaneous differential detection of Chlamydophila abortus, Chlamydophila pecorum and Coxiella burnetii from aborted ruminant’s clinical samples using multiplex PCR. BMC Microbiol. 2009, 9, 130. Available online: http://www.biomedcentral.com/1471-2180/9/130 (accessed on 6 February 2022). [CrossRef]
- Azeem, S.; Ghorbani-Nezami, S.; Allam, A.; Cooper, V.L.; Burrough, E.; Schwartz, K.; Plummer, P.; Arruda, P.; Gauger, P.; Kinyon, J.M.; et al. A multiplex real-time PCR panel for detection of major bacterial and viral agents causing abortion in small ruminants. In Proceedings of the American Association of Veterinary Laboratory Diagnosticians Annual Conference, Providence, RI, USA, 22–28 October 2015. [Google Scholar]
- Tramutra, C.; Lacerenza, D.; Zoppi, S.; Goria, M.; Dondo, A.; Ferroglio, E.; Nebbia, P.; Rosati, S. Development of a set of multiplex standard polymerase chain reaction assays for the identification of infectious agents from aborted bovine clinical samples. J. Vet. Diagn. Investig. 2011, 23, 657–664. [Google Scholar] [CrossRef]
- Jonker, A.; Thompson, P.N.; Michel, A.L. Approaches to increase recovery of bacterial and fungal abortion agents in domestic ruminants, Onderstepoort. J. Vet. Res. 2022, 90, a2010. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Hugget, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum information for publication of quantitative Real-Time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Kralik, P.; Ricchi, M. A basic guide to real time PCR in microbial diagnostics: Definitions, parameters, and everything. Front. Microbiol. 2017, 8, 108. [Google Scholar] [CrossRef]
- Selim, A.; Elhaigh, M.M.; Gaede, W. Development of multiplex real-time PCR assay for the detection of Brucella spp., Leptospira spp. and Campylobacter foetus. Vet. Ital. 2014, 50, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Deuchande, R.; Gidlow, J.; Caldow, G.; Baily, J.; Longbottom, D.; Wheelhouse, N.; Borel, N.; Greub, G. Parachlamydia involvement in bovine abortions in a beef herd in Scotland. Vet. Rec. 2010, 166, 598–599. [Google Scholar] [CrossRef] [PubMed]
- Merdja, S.-E.; Khaled, H.; Aaziz, R.; Vorimore, F.; Bertin, C.; Dahmani, A.; Bouyoucef, A.; Laroucau, K. Detection and genotyping of Chlamydia species responsible for reproductive disorders in Algerian small ruminants. Trop. Anim. Health 2015, 47, 437–443. [Google Scholar] [CrossRef] [PubMed]



| Bacteria | Source or Strain |
|---|---|
| Acholeplasma laidlawlii | NCTC 10116 |
| Brucella abortus | Clinical sample |
| Chlamydia abortus | ATCC VR-165 |
| Escherichia coli | ATCC 25922 |
| Ochrobactrum anthropi | ATCC 49687 |
| Parachlamydia acanthamoeba | ATCC VR-1476 |
| Pasteurella multocida | ATCC 12945 |
| Salmonella Typhimurium | ATCC 13311 |
| Staphylococcus aureus | ATCC 25923 |
| Streptococcus agalactiae | ATCC 27956 |
| Trueperella pyogenes | Clinical sample |
| Waddlia chondrophila | ATCC VR-1470 |
| Amoeba | |
| Acanthamoeba castellani | ATCC 50739 |
| Case No. | Pathology | Bacteria/Fungi (Previous Study [41]) | Samples | DNA Detected (Mean Ct Values) |
|---|---|---|---|---|
| Bovine | ||||
| 1 | Necrotic placentitis Purulent bronchopneumonia | Bacillus licheniformis Rhizopus sp. | Stomach content | ND |
| Lung | ND | |||
| 2 | Necrotic placentitis and vasculitis | Mycoplasma sp. Aspergillus fumigatus | Placenta | PanChl (36.05) |
| Stomach content | PanChl (33.38) Chlab (32.33) Chlpec (36.90) Wchon (31.85) | |||
| Lung | ND | |||
| 3 | Necrotic placentitis | ND | Stomach content | ND |
| 4 | Necrotic placentitis | Salmonella sp. | Placenta | PanChl (32.90) Wchon (26.02) |
| 5 | Necrotic placentitis with vasculitis | Coxiella burnetii | Stomach content | PanChl (32.25) Chlab (32.18) Chlpec (35.86) |
| 6 | Necrotic placentitis | Streptococcus dysgalactiae Acholeplasma sp. | Placenta | PanChl (28.63) |
| Stomach content | Wchon (34.36) | |||
| Lung | ND | |||
| 7 | Purulent placentitis | ND | Placenta | ND |
| 8 | Necrotic placentitis | Lichtheimia corymbifera | Placenta | PanChl (31.21) Wchon (23.97) |
| Stomach content | ND | |||
| Lung | ND | |||
| 9 | Meningitis bronchointerstitial pneumonia | Brucella abortus biovar 1 | Stomach content | ND |
| 10 | Meningoencephalitis Necrotic placentitis Hepatic necrosis | ND | Stomach content | ND |
| 11 | Necrotic placentitis with vasculitis | ND | Placenta | ND |
| ND | Lung | ND | ||
| 12 | Necrotic placentitis | Campylobacter fetus Mannheimia varigena Streptococcus pluranimalium Mycoplasma sp. | Placenta | PanChl (30.02) Pacanth (36.88) Wchon (32.47) |
| Stomach content | ND | |||
| Lung | ND | |||
| 13 | Necrotic placentitis Purulent pneumonia | Chlamydia sp. Salmonella sp. | Placenta | PanChl (31.63) |
| Stomach content | PanChl (33.94) | |||
| Lung | ND | |||
| 14 | Necrotic placentitis | ND | Placenta | PanChl (5.03) Chlab (5.65) |
| Stomach content | ND | |||
| 15 | Viral placentitis | Pasteurella multocida | Placenta | Chlab (30.79) |
| 16 | Interstitial pneumonia | Brucella abortus biovar 1 | Stomach content | ND |
| 17 | Necropurulent placentitis | Trueperella pyogenes | Placenta | PanChl (34.68) Pacanth (33.48) |
| 18 | Necrotic placentitis Pneumonia | Chlamydia sp. | Placenta | PanChl (23.13) Chlpec (18.96) |
| Stomach content | PanChl (31.69) Chlpec (29.83) | |||
| 19 | Bacterial & fungal placentitis Pleuropneumonia | Chlamydia abortus | Stomach content | ND |
| Lung | ND | |||
| 20 | Bronchopneumonia | ND | Stomach content | PanChl (27.42) Chlpec (22.19) Wchon (35.17) |
| Lung | PanChl (25.74) Chlpec (23.42) Wchon (35.39) | |||
| 21 | Bacterial placentitis | Arcobacter sp. | Placenta | ND |
| Lung | ND | |||
| 22 | Necrotic placentitis | Salmonella Budapest Mycoplasma species | Placenta | PanChl (35.03) Wchon (33.35) |
| Stomach content | ND | |||
| 23 | Fibrinopurulent bacterial pneumonia | ND | Placenta | PanChl (29.49) Chlpec (34.48) Wchon (32.52) |
| Lung | ND | |||
| 24 | Necrotic placentitis | Coxiella burnetiid Salmonella typhimurium | Placenta | ND |
| Stomach content Foetus A | ND | |||
| Lung Foetus A | PanChl (32.41) Chlpec (35.84) Wchon (31.57) | |||
| Stomach content Foetus B | ND | |||
| Lung Foetus B | ND | |||
| 25 | Necrotizing placentitis Subcutaneous oedema Hydrocephalus | E. coli Lichtheimia corymbifera | Placenta | PanChl (10.27) |
| Stomach content | ND | |||
| Lung | ND | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jonker, A.; Michel, A.L. Optimization and Application of Real-Time qPCR Assays in Detection and Identification of Chlamydiales in Products of Domestic Ruminant Abortion. Pathogens 2023, 12, 290. https://doi.org/10.3390/pathogens12020290
Jonker A, Michel AL. Optimization and Application of Real-Time qPCR Assays in Detection and Identification of Chlamydiales in Products of Domestic Ruminant Abortion. Pathogens. 2023; 12(2):290. https://doi.org/10.3390/pathogens12020290
Chicago/Turabian StyleJonker, Annelize, and Anita L. Michel. 2023. "Optimization and Application of Real-Time qPCR Assays in Detection and Identification of Chlamydiales in Products of Domestic Ruminant Abortion" Pathogens 12, no. 2: 290. https://doi.org/10.3390/pathogens12020290