
Campylobacter jejuni Response When Inoculated in Bovine In Vitro Fecal Microbial Consortia Incubations in the Presence of Metabolic Inhibitors
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbial Sources
2.2. Comparison of C. jejuni in Fecal versus Rumen Mixed Microbial Populations
2.3. Impact of Different Gas Atmospheres and Anti-Methanogenic Treatments on C. jejuni Survivability and Select Incubation Characteristics in Mixed Rumen and Fecal Populations
2.4. Statistical Analysis
3. Results
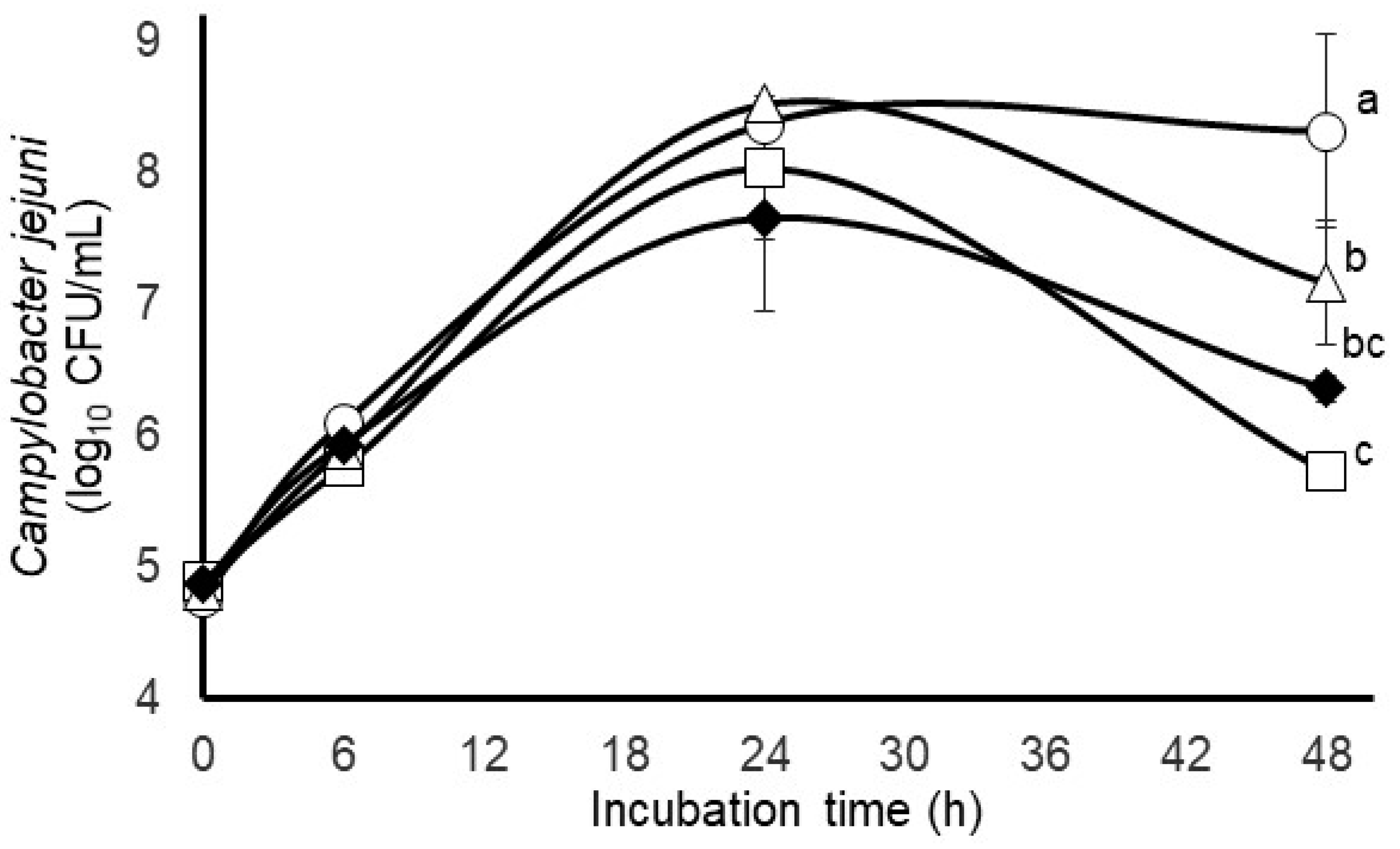
3.1. Comparison of C. jejuni Survival in Mixed Rumen and Bovine Fecal In Vitro Microbial Cultures
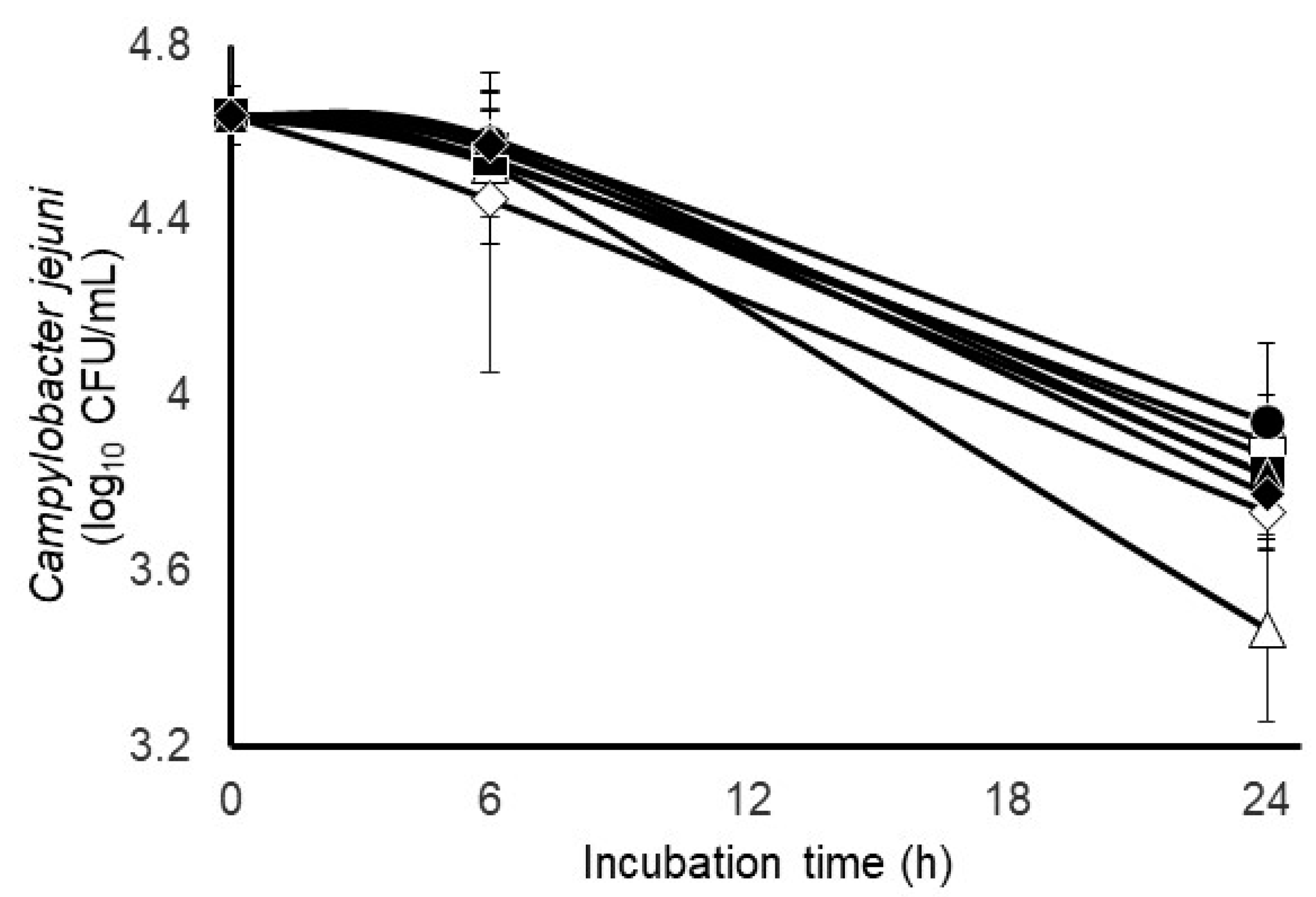
3.2. Impact of Different Gas Atmospheres and Anti-Methanogenic Treatments on C. jejuni Survivability and Select Incubation Characteristics in Mixed Fecal Populations
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pearce, R.A.; Wallace, F.M.; Call, J.E.; Dudley, R.L.; Oser, A.; Yoder, L.; Sheridan, J.J.; Luchansky, J.B. Prevalence of Campylobacter within a swine slaughter and processing facility. J. Food Prot. 2003, 66, 1550–1556. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.E.; Corcoran, D.; Dooley, J.S.G.; Fanning, S.; Lucey, B.; Matsuda, M.; McDowell, D.A.; Mégraud, F.; Millar, B.C.; O’Mahony, R.; et al. Campylobacter. Vet. Res. 2005, 36, 351–382. [Google Scholar] [CrossRef] [PubMed]
- Corry, J.E.L.; Atabay, H.I. Poultry as a source of Campylobacter and related organisms. J. Appl. Microbiol. 2001, 90, 96S–114S. [Google Scholar] [CrossRef]
- Sahin, O.; Morishita, T.Y.; Zhang, Q. Campylobacter colonization in poultry: Sources of infection and modes of transmission. Anim. Health Res. Rev. 2002, 3, 95–105. [Google Scholar] [CrossRef]
- Horrocks, S.M.; Anderson, R.C.; Nisbet, D.J.; Ricke, S.C. Incidence and ecology of Campylobacter in animals. Anaerobe 2009, 15, 18–25. [Google Scholar] [CrossRef]
- Umaraw, P.; Prajapati, A.; Verma, A.K.; Pathak, V.; Singh, V.P. Control of Campylobacter in poultry industry from farm to poultry processing unit: A review. Critical Revs. Food Sci. Nutr. 2017, 57, 659–665. [Google Scholar] [CrossRef]
- Stanley, K.; Jones, K. Cattle and sheep farms as reservoirs of Campylobacter. J. Appl. Microbiol. 2003, 94, 104S–113S. [Google Scholar] [CrossRef]
- Harvey, R.B.; Young, C.R.; Ziprin, R.L.; Hume, M.E.; Genovese, K.J.; Anderson, R.C.; Droleskey, R.E.; Stanker, L.H.; Nisbet, D.J. Prevalence of Campylobacter spp. isolated from the intestinal tract of pigs raised in an integrated swine production system. J. Am. Vet. Med. A 1999, 215, 1601–1604. [Google Scholar]
- Jensen, A.N.; Dalsgaard, A.; Baggesen, D.L.; Nielsen, E.M. The occurrence and characterization of Campylobacter jejuni and C. coli in organic pigs and their outdoor environment. Vet. Microbiol. 2006, 116, 96–105. [Google Scholar] [CrossRef]
- Natsos, G.; Koutoulis, K.C.; Sossidou, E.; Chemaly, M.; Mouttotou, N.K. Campylobacter spp. infection in humans and poultry. J. Hell. Vet. Med. Soc. 2016, 67, 65–82. [Google Scholar] [CrossRef]
- Chlebicz, A.; Śliżewska, K. Campylobacteriosis, salmonellosis, yersiniosis, and listeriosis as zoonotic foodborne diseases: A review. Int. J. Environ. Res. Public Health 2018, 15, 863. [Google Scholar] [CrossRef]
- Yang, Y.; Feye, K.M.; Shi, Z.; Pavlidis, H.O.; Kogut, M.; Ashworth, J.A.; Ricke, S.C. A historical review on antibiotic resistance of foodborne Campylobacter. Front. Microbiol. 2019, 10, 1509. [Google Scholar] [CrossRef]
- Ricke, S.C.; Feye, K.M.; Chaney, W.E.; Shi, Z.; Pavlidis, H.; Yang, Y. Developments in rapid detection methods for the detection of foodborne Campylobacter in the United States. Front. Microbiol. 2019, 9, 3280. [Google Scholar] [CrossRef] [PubMed]
- Grout, L.; Marshall, J.; Hales, S.; Baker, M.G. Dairy cattle density and temporal patterns of human campylobacteriosis and cryptosporidiosis in New Zealand. EcoHealth 2022, 19, 273–289. [Google Scholar] [CrossRef]
- An, J.-U.; Ho, H.; Kim, J.; Kim, W.H.; Kim, J.; Lee, S.; Mun, S.-H.; Guk, J.-H.; Hong, S.; Cho, S. Dairy cattle, a potential reservoir of human campylobacteriosis: Epidemiological and molecular characterization of Campylobacter jejuni from cattle farms. Front. Microbiol. 2018, 9, 3136. [Google Scholar] [CrossRef]
- Aksomaitiene, J.; Ramonaite, S.; Tamuleviciene, E.; Novoslavskij, A.; Alter, T.; Malakauskas, M. Overlap of antibiotic resistant Campylobacter jejuni MLST genotypes isolated from humans, broiler products, dairy cattle and wild birds in Lithuania. Front. Microbiol. 2019, 10, 1377. [Google Scholar] [CrossRef] [PubMed]
- St Charles, J.L.; Brooks, P.T.; Bell, J.A.; Ahmed, H.; Van Allen, M.; Manning, S.D.; Mansfield, L.S. Zoonotic transmission of Campylobacter jejuni to caretakers from sick pen calves carrying a mixed population of strains with and without Guillain Barré Syndrome-associated lipooligosaccharide loci. Front. Microbiol. 2022, 13, 800269. [Google Scholar] [CrossRef]
- Bae, W.; Hancock, D.D.; Call, D.R.; Park, Y.H.; Berge, A.C.B.; Finger, R.M.; Sischo, W.M.; Besser, T.E. Dissemination of antimicrobial resistant strains of Campylobacter coli and Campylobacter jejuni among cattle in Washington State and California. Vet. Microbiol. 2007, 122, 306–315. [Google Scholar] [CrossRef]
- Bae, W.; Kaya, K.N.; Hancock, D.D.; Call, D.R.; Park, Y.H.; Besser, T.E. Prevalence and antimicrobial resistance of thermophilic Campylobacter spp. from cattle farms in Washington state. Appl. Environ. Microbiol. 2005, 71, 169–174. [Google Scholar] [CrossRef]
- Karama, M.K.; Kambuyi, B.T.; Cenci-Goga, M.; Malahlela, A.; Jonker, C.; He, J.; Ombui, T.; Tshuma, T.; Etter, E.; Kalake, A. Occurrence and antimicrobial resistance profiles of Campylobacter jejuni, Campylobacter coli, and Campylobacter upsaliensis in beef cattle on cow–calf operations in South Africa. Foodborne Path. Dis. 2020, 17, 440–446. [Google Scholar] [CrossRef]
- Plishka, M.; Sargeant, J.M.; Greer, A.L.; Hookey, S.; Winder, C. The prevalence of Campylobacter in live cattle, turkey, chicken, and swine in the United States and Canada: A systematic review and meta-analysis. Foodborne Path. Dis. 2021, 18, 230–242. [Google Scholar] [CrossRef] [PubMed]
- Premarathne, J.M.K.J.K.; Anuar, A.S.; Thung, T.Y.; Satharasinghe, D.A.; Jambari, N.N.; Abdul-Mutalib, N.A.; Huat, J.T.Y.; Basri, D.F.; Rukayadi, Y.; Nakaguchi, Y.; et al. Prevalence and antibiotic resistance against tetracycline in Campylobacter jejuni and C. coli in cattle and beef meat from Selangor, Malaysia. Front. Microbiol. 2017, 8, 2254. [Google Scholar] [CrossRef] [PubMed]
- Karki, A.B.; Marasini, D.; Oakey, C.K.; Mar, K.; Fakhr, M.K. Campylobacter coli from retail liver and meat products is more aerotolerant than Campylobacter jejuni. Front. Microbiol. 2018, 9, 2951. [Google Scholar] [CrossRef] [PubMed]
- Ghatak, S.; He, Y.; Reed, S.; Irwin, P. Comparative genomic analysis of a multidrug-resistant Campylobacter jejuni strain YH002 isolated from retail beef liver. Foodborne Path. Dis. 2020, 17, 576–584. [Google Scholar] [CrossRef]
- Facciolà, A.; Riso, R.; Avventuroso, E.; Visalli, G.; Delia, S.A.; Lagana, P. Campylobacter: From microbiology to prevention. J. Prev. Med. Hyg. 2017, 58, E79–E92. [Google Scholar]
- Dittoe, D.K.; Anderson, R.C.; Krueger, N.A.; Harvey, R.B.; Poole, T.L.; Crippen, T.L.; Callaway, T.R.; Ricke, S.C. Survival of Campylobacter jejuni during in vitro culture with mixed bovine ruminal microorganisms in the presence of methanogen inhibitors. J. Environ. Sci. Health B 2023, 58, 711–718. [Google Scholar] [CrossRef]
- Byrd, J.A.; Hargis, B.M.; Caldwell, D.J.; Herron, K.L.; McReynolds, J.L.; Brewer, R.L.; Anderson, R.C.; Bischoff, K.M.; Callaway, T.R.; Kubena, L.F. Effect of lactic acid administration in the drinking water during preslaughter feed withdrawal on Salmonella and Campylobacter contamination of broilers. Poult. Sci. 2001, 80, 278–283. [Google Scholar] [CrossRef]
- Anderson, R.C.; Krueger, N.A.; Byrd, J.A.; Harvey, R.B.; Callaway, T.R.; Edrington, T.S.; Nisbet, D.J. Effects of thymol and diphenyliodonium chloride against Campylobacter spp. during pure and mixed culture in vitro. J. Appl. Microbiol. 2009, 107, 1258–1268. [Google Scholar] [CrossRef]
- Stern, N.J.; Wojton, B.; Kwiater, K. A differential selective medium, and dry-ice generated atmosphere for recovery of Campylobacter jejuni. J. Food Prot. 1992, 55, 514–517. [Google Scholar] [CrossRef]
- Bryant, M.P.; Burkey, L.A. Cultural methods and some characteristics of some of the more numerous groups of bacteria in the bovine rumen. J. Dairy Sci. 1953, 36, 205–217. [Google Scholar] [CrossRef]
- Chaney, A.L.; Marbach, E.P. Modified reagents for determination of urea and ammonia. Clin. Chem. 1962, 8, 130–132. [Google Scholar] [CrossRef] [PubMed]
- Horrocks, S.M.; Jung, Y.S.; Huwe, J.K.; Harvey, R.B.; Ricke, S.C.; Carstens, G.E.; Callaway, T.R.; Anderson, R.C.; Ramlachan, N.; Nisbet, D.J. Effects of short-chain nitrocompounds against Campylobacter jejuni and Campylobacter coli in vitro. J. Food Sci. 2007, 72, M50–M55. [Google Scholar] [CrossRef] [PubMed]
- Cataldo, D.A.; Haroon, M.; Schrader, L.E.; Youngs, V.L. Rapid colorimetric determination of nitrate in plant tissue by nitration of salicylic acid. Commun. Soil Sci. Plant Anal. 1975, 6, 71–80. [Google Scholar] [CrossRef]
- Anderson, R.C.; Ripley, L.H.; Bowman, J.G.P.; Callaway, T.R.; Genovese, K.J.; Beier, R.C.; Harvey, R.B.; Nisbet, D.J. Ruminal fermentation of anti-methanogenic nitrate- and nitro-containing forages in vitro. Front. Vet. Sci 2016, 3, 62. [Google Scholar] [CrossRef] [PubMed]
- Indikova, I.; Humphrey, T.J.; Hilbert, F. Survival with a helping hand: Campylobacter and microbiota. Front. Microbiol. 2015, 6, 1266. [Google Scholar] [CrossRef] [PubMed]
- Krueger, N.A.; Anderson, R.C.; Krueger, W.K.; Horne, W.J.; Callaway, T.R.; Edrington, T.S.; Carstens, G.E.; Harvey, R.B.; Nisbet, D.J. Prevalence and concentration of Campylobacter in rumen contents and feces in pasture and feedlot fed cattle. Foodborne Path. Dis. 2008, 5, 571–577. [Google Scholar] [CrossRef]
- Deng, W.; Dittoe, D.K.; Pavilidis, H.O.; Chaney, W.E.; Yang, Y.; Ricke, S.C. Current perspectives and potential of probiotics to limit foodborne Campylobacter in poultry. Front. Microbiol. 2020, 11, 583429. [Google Scholar] [CrossRef]
- Rath, A.; Rautenschlein, S.; Rzeznitzeck, J.; Lalk, M.; Methling, K.; Rychlik, I.; Peh, E.; Kittler, S.; Waldmann, K.-H.; von Altrock, A. Investigation on the colonisation of Campylobacter strains in the pig intestine depending on available metabolites. Comp. Immunol. Microbiol. Infect. Dis. 2022, 88, 101865. [Google Scholar] [CrossRef]
- Welch, C.B.; Lourenco, J.M.; Seidel, D.S.; Krause, T.R.; Rothrock, M.J.; Pringle, T.D.; Callaway, T.R. The impact of pre-slaughter fasting on the ruminal microbial population of commercial Angus steers. Microorganisms 2021, 9, 2625. [Google Scholar] [CrossRef]
- Feye, K.M.; Rubinelli, P.M.; Chaney, W.E.; Pavlidis, H.O.; Kogut, M.H.; Ricke, S.C. The preliminary development of an in vitro poultry cecal culture model to evaluate the effects of Original XPCTM for the reduction of Campylobacter jejuni and its potential effects on the microbiota. Front. Microbiol. 2020, 10, 3062. [Google Scholar] [CrossRef]
- Sergeant, M.J.; Constantinidou, C.; Cogan, T.A.; Bedford, M.R.; Penn, C.W.; Pallen, M.J. Extensive microbial and functional diversity within the chicken cecal microbiome. PLoS ONE 2014, 9, e91941. [Google Scholar] [CrossRef] [PubMed]
- Epps, S.V.R.; Harvey, R.B.; Hume, M.E.; Phillips, T.D.; Anderson, R.C.; Nisbet, D.J. Foodborne Campylobacter: Infections, metabolism, pathogenesis and reservoirs. Int. J. Environ. Res. Public Health. 2013, 10, 6292–6304. [Google Scholar] [CrossRef] [PubMed]
- Pattison, M. Practical intervention strategies for Campylobacter. J. Appl. Microbiol. 2001, 90, 121S–125S. [Google Scholar] [CrossRef] [PubMed]
- Rath, A.; Rautenschlein, S.; Rzeznitzeck, J.; Breves, G.; Hewicker-Trautwein, M.; Waldmann, K.-H.; von Altrock, A. Impact of Campylobacter spp. on the integrity of the porcine gut. Animals 2021, 11, 2742. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Campylobacter jejuni Mean Specific Growth Rate (h−1) | Total Culturable Anaerobes (log10 CFUs/mL) | pH | Ammonia (µmol/mL) | |
|---|---|---|---|---|
| Treatments | during initial 6 h incubation | After 48 h incubation | ||
| Campylobacter jejuni only | 0.222 | NA † | 6.42 a | 1.25 |
| 2X Rumen fluid | 0.145 | 7.13 | 6.18 b | 1.90 |
| 2X Feces | 0.181 | 7.17 | 6.16 b | 1.75 |
| 1X Rumen fluid/1X feces | 0.178 | 7.15 | 6.17 b | 1.83 |
| Treatment effect | p = 0.1860 | p = 0.7122 | p = 0.0474 | p = 0.1983 |
| Standard error of the mean | 0.022 | 0.037 | 0.062 | 0.211 |
| Change in Campylobacter jejuni (log10 CFUs/mL) | Gas Produced (mL) | Hydrogen Accumulation (µmol/mL) | Methane Accumulation (µmol/mL) | Nitrate Metabolized † (µmol/mL) | ||
|---|---|---|---|---|---|---|
| Parameter | After 6 h | After 24 h | After 24 h | After 24 h | After 24 h | After 24 h |
| Initial gas phase | ||||||
| 100% carbon dioxide | −0.12 | −0.90 | 3.81 a | 0.52 b | 0.51 a | 2.06 |
| 50:50 Hydrogen/carbon dioxide | −0.07 | −0.80 | 2.11 b | 32.07 a | 0.04 b | 1.99 |
| p value | 0.4617 | 0.2123 | 0.0296 | <0.0001 | 0.0139 | 0.9233 |
| SEM | 0.050 | 0.055 | 0.517 | 0.854 | 0.123 | 0.523 |
| Anti-methanogen treatment | ||||||
| No treatment | −0.08 | −0.72 | 3.75 x | 17.26 | 0.13 | ND |
| 0.05 mM 2-Bromosulfonate | −0.09 | −0.80 | 4.87 x | 18.29 | 0.21 | ND |
| 5 mM sodium nitrate | −0.08 | −0.99 | 1.50 y | 15.42 | 0.32 | 1.81 |
| Combined | −0.13 | −0.88 | 1.72 y | 14.20 | 0.43 | 2.24 |
| p value | 0.9628 | 0.0613 | 0.0011 | 0.9773 | 0.7465 | 0.5678 |
| SEM | 0.075 | 0.070 | 0.579 | 7.13 | 0.204 | 0.514 |
| Change in Campylobacter jejuni (log10 CFUs/mL) | Gas Produced (mL) | Hydrogen Accumulation (µmol/mL) | Methane Accumulation (µmol/mL) | Ammonia Accumulation (µmol/mL) | ||
|---|---|---|---|---|---|---|
| Parameter | After 6 h | After 24 h | After 24 h | After 24 h | After 24 h | After 24 h |
| Anti-methanogen treatment | ||||||
| No Treatment | −0.16 | NA† | 18.67 | 0.25 | Undectable | 0.96 |
| 0.05 mM 2-Bromosulfonate | −0.08 | NA | 17.67 | 0.15 | Undectable | 0.04 |
| 0.001 mM Monensin | −0.02 | NA | 19.00 | 0.54 | Undectable | 0.11 |
| Combined | −0.14 | NA | 18.00 | 0.40 | Undectable | 0.19 |
| p value | 0.6080 | - | 0.1189 | 0.2086 | - | 0.0856 |
| SEM | 0.080 | - | 0.373 | 0.130 | - | 0.242 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dittoe, D.K.; Anderson, R.C.; Krueger, N.A.; Harvey, R.B.; Poole, T.L.; Crippen, T.L.; Callaway, T.R.; Ricke, S.C. Campylobacter jejuni Response When Inoculated in Bovine In Vitro Fecal Microbial Consortia Incubations in the Presence of Metabolic Inhibitors. Pathogens 2023, 12, 1391. https://doi.org/10.3390/pathogens12121391
Dittoe DK, Anderson RC, Krueger NA, Harvey RB, Poole TL, Crippen TL, Callaway TR, Ricke SC. Campylobacter jejuni Response When Inoculated in Bovine In Vitro Fecal Microbial Consortia Incubations in the Presence of Metabolic Inhibitors. Pathogens. 2023; 12(12):1391. https://doi.org/10.3390/pathogens12121391
Chicago/Turabian StyleDittoe, Dana K., Robin C. Anderson, Nathan A. Krueger, Roger B. Harvey, Toni L. Poole, Tawni L. Crippen, Todd R. Callaway, and Steven C. Ricke. 2023. "Campylobacter jejuni Response When Inoculated in Bovine In Vitro Fecal Microbial Consortia Incubations in the Presence of Metabolic Inhibitors" Pathogens 12, no. 12: 1391. https://doi.org/10.3390/pathogens12121391